
Tar Spot on Maize: Impact of Soil Types and Environmental Conditions on the Survival of Phyllachora maydis in the Subtropical Climate of Florida



Abstract
1. Introduction
2. Materials and Methods
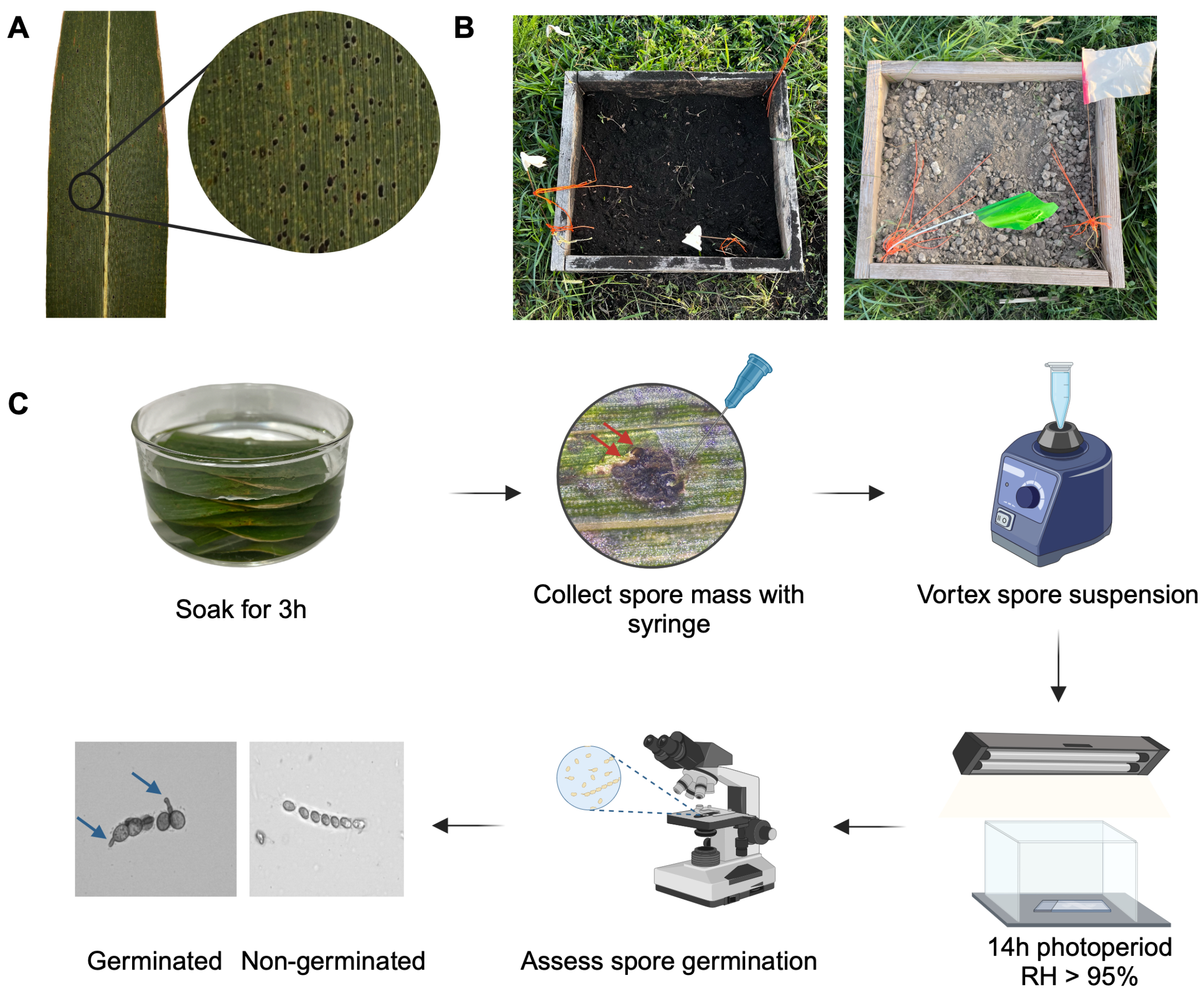
2.1. Experimental Design
2.2. Assessment of Leaf Decomposition, Tar Spot Stromata Integrity, and Ascospore Viability
2.2.1. Leaf Retrieval and Sampling Procedures
2.2.2. Leaf Decomposition and Tar Spot Stroma Integrity Assessment
2.2.3. Ascospore Germination Assay and Quantification
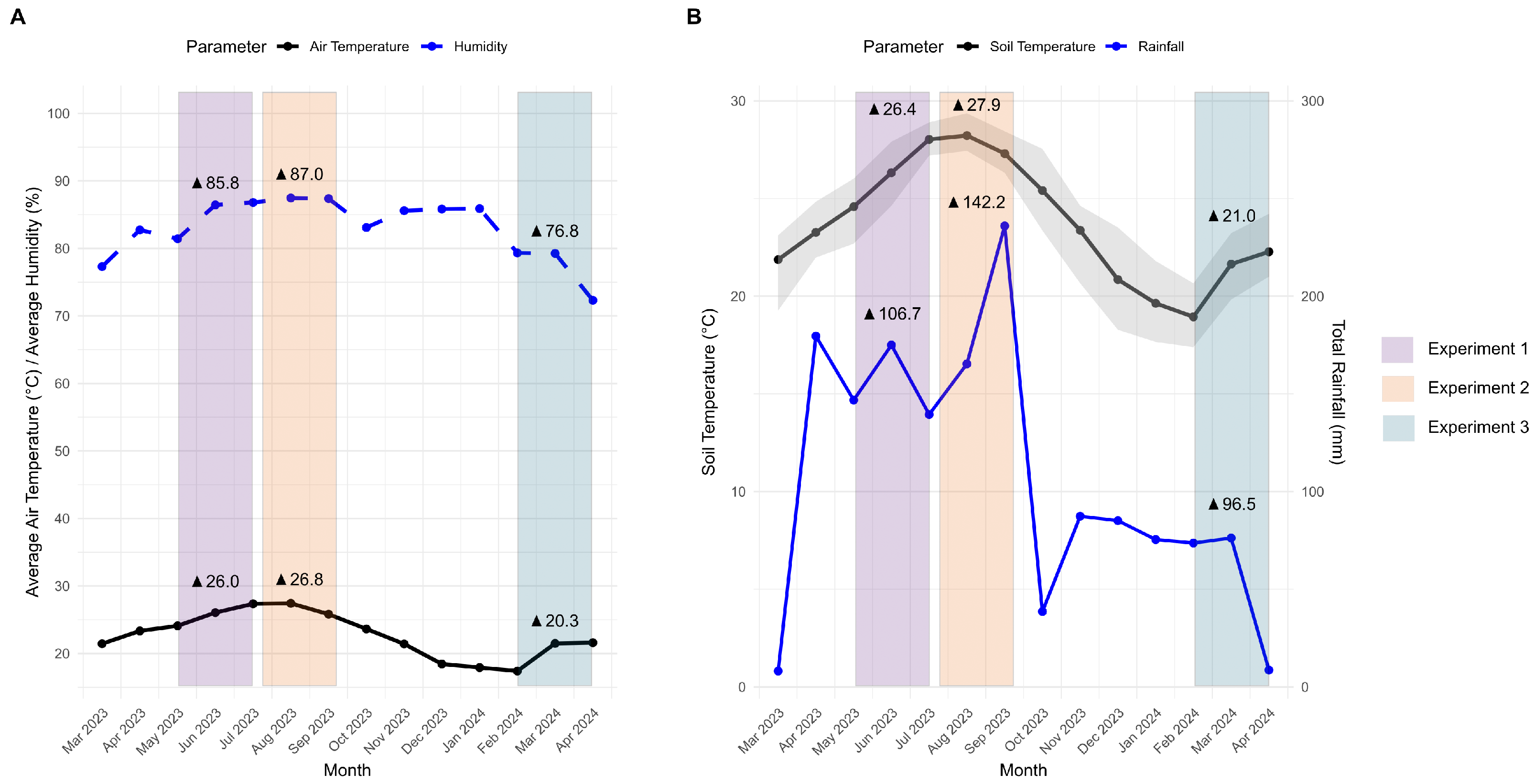
2.3. Weather Data Processing
2.4. Statistical Analysis
3. Results
3.1. Seasonal Variations and Environmental Factors Affecting Phyllachora maydis Survival and Tar Spot-Related Leaf Decomposition
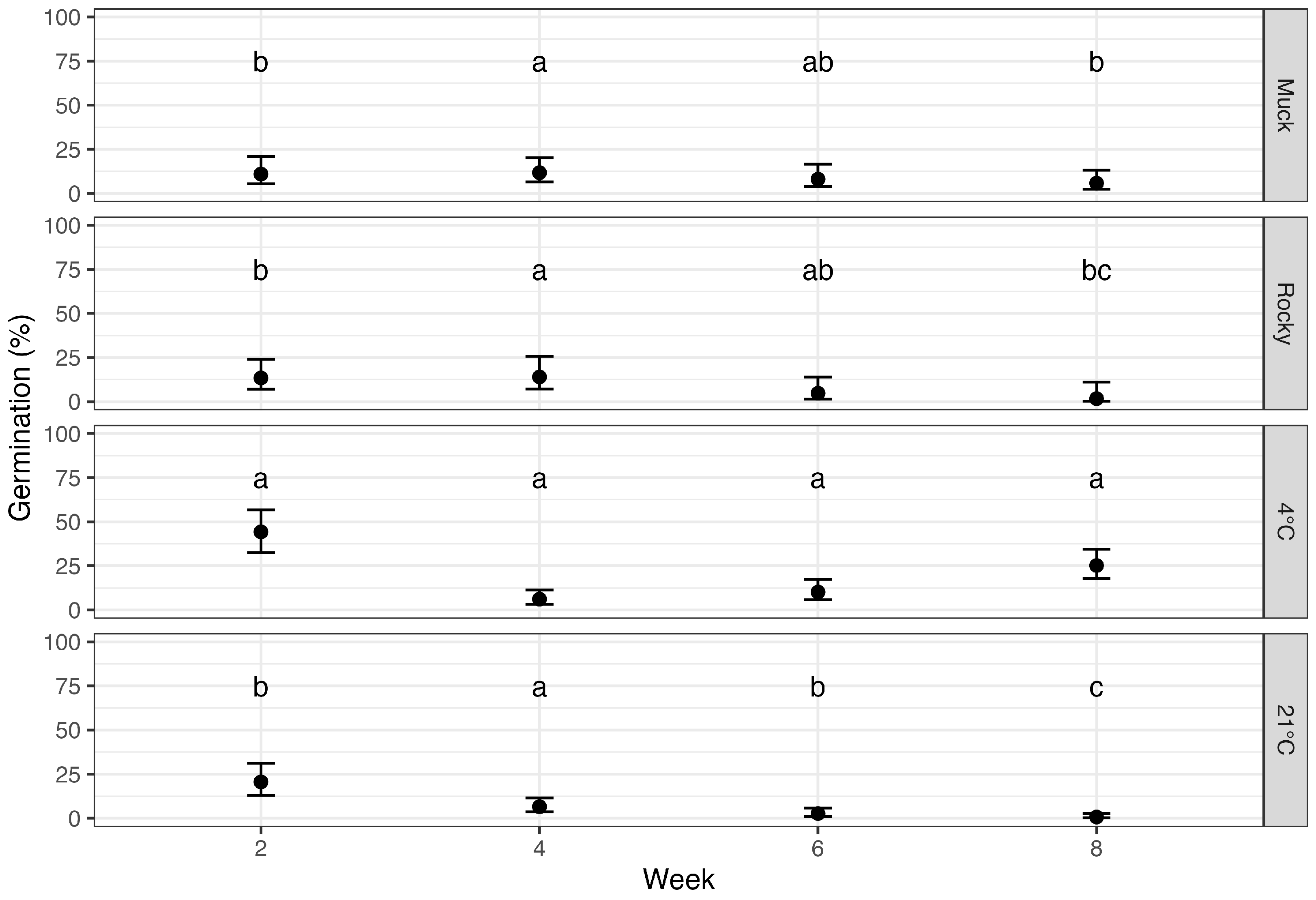
3.2. Effect of Soil Types and Environmental Conditions on P. maydis Ascospore Germination
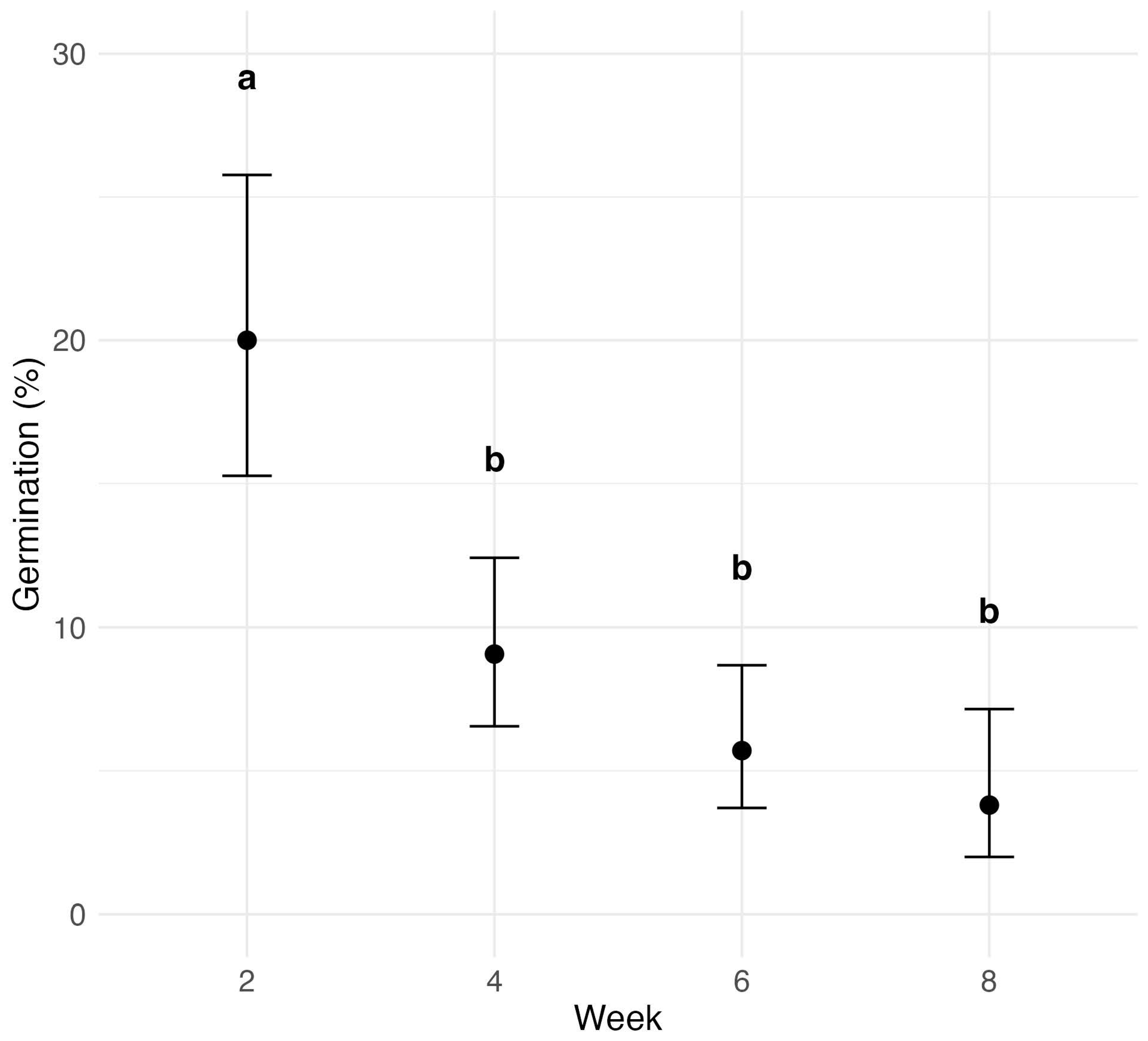
3.3. Temporal Impact on Ascospore Germination Post-Leaf Detachment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eckhoff, S.R.; Paulsen, M.R. Maize. In Cereal Grain Quality; Henry, R.J., Kettlewell, P.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 77–98. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J. Maize. In Cereals Handbook of Plant Breeding; Carena, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 3, pp. 3–98. [Google Scholar] [CrossRef]
- Tracy, W.F. Sweet corn. In Specialty Corns, 2nd ed.; Hallauer, A.R., Ed.; CRC: Boca Raton, FL, USA, 2001; pp. 155–199. [Google Scholar] [CrossRef]
- USDA National Agricultural Statistical Service. Quick Stats. 2024 State Agriculture Overview Florida. 2025. Available online: https://www.nass.usda.gov/Quick_Stats/Ag_Overview/stateOverview.php?state=FLORIDA (accessed on 3 June 2025).
- Mueller, D.S.; Wise, K.A.; Sisson, A.J.; Allen, T.W.; Bergstrom, G.C.; Bissonnette, K.M.; Bradley, C.A.; Byamukama, E.; Chilvers, M.I.; Collins, A.A.; et al. Corn yield loss estimates due to diseases in the United States and Ontario, Canada, from 2016 to 2019. Plant Health Prog. 2020, 21, 238–247. [Google Scholar] [CrossRef]
- Ruhl, G.; Romberg, M.K.; Bissonnette, S.; Plewa, D.; Creswell, T.; Wise, K.A. First report of tar spot on corn caused by Phyllachora maydis in the United States. Plant Dis. 2016, 100, 1496. [Google Scholar] [CrossRef]
- Mueller, D.; Wise, K.; Sisson, A. Corn Disease Loss Estimates from the United States and Ontario, Canada–2018. United States: Crop Protection Network. 2020. Available online: https://cropprotectionnetwork.org/publications/corn-disease-loss-estimates-2018 (accessed on 3 June 2025).
- Telenko, D.E.P.; Chilvers, M.I.; Kleczewski, N.; Smith, D.L.; Byrne, A.M.; Devillez, P.; Diallo, T.; Higgins, R.; Joos, D.; Kohn, K. How tar Spot of Corn Impacted Hybrid Yields During the 2018 Midwest Epidemic. United States: Crop Protection Network. 2019. Available online: https://cropprotectionnetwork.org/publications/how-tar-spot-of-corn-impacted-hybrid-yields-during-the-2018-midwest-epidemic#:~:text=It%20was%20estimated%20that%20a,evaluated%20in%20Indiana%20in%202018 (accessed on 3 June 2025).
- Sikora, E.; Faske, T.; Meyer, R.; Koehler, A.; Kemerait, B.; Harbach, C.; Telenko, D.; Robertson, A.; Onofre, R.; Price, T.; et al. Corn Disease Loss Estimates from the United States and Ontario, Canada—2021. Ames (Iowa): Iowa State University. 2022. Available online: https://cropprotectionnetwork.org/publications/corn-disease-loss-estimates-from-the-united-states-and-ontario-canada-2021 (accessed on 3 June 2025).
- Mueller, D.; Wise, K.; Sisson, A.; Sikora, E.; Faske, T.; Meyer, R.; Koehler, A.; Kemerait, B.; Harbach, C.; Telenko, D.; et al. Corn Disease Loss Estimates from the United States and Ontario, Canada—2022. United States: Crop Protection Network. 2023. Available online: https://cropprotectionnetwork.org/publications/corn-disease-loss-estimates-from-the-united-states-and-ontario-canada-2022 (accessed on 3 June 2025).
- Sikora, E.; Faske, T.; Meyer, R.; Koehler, A.; Kemerait, B.; Mideros, S.; Telenko, D.; Robertson, A.; Mueller, D.; Sisson, A.; et al. Corn Disease Loss Estimates from the United States and Ontario, Canada—2023; Crop Protection Network: Ames, IA, USA, 2024. [Google Scholar] [CrossRef]
- Sikora, E.; Faske, T.; Meyer, R.; Betts, A.; Kemerait, B.; Camiletti, B.; Telenko, D.; Robertson, A.; Mueller, D.; Sisson, A.; et al. Corn Disease Loss Estimates from the United States and Ontario, Canada—2024; Crop Protection Network: Ames, IA, USA, 2025. [Google Scholar] [CrossRef]
- Ferreira, L.C.; Moura, V.A.S.; Resende, M.F.; Xavier, K.V. Tar Spot of Maize in Florida. EDIS 2024. [Google Scholar] [CrossRef]
- McCoy, A.G.; Romberg, M.K.; Zaworski, E.R.; Robertson, A.E.; Phibbs, A.; Hudelson, B.D.; Smith, D.L.; Beiriger, R.L.; Raid, R.N.; Byrne, J.M.; et al. First report of tar spot on corn (Zea mays) caused by Phyllachora maydis in Florida, Iowa, Michigan, and Wisconsin. Plant Dis. 2018, 102, 1851. [Google Scholar] [CrossRef]
- Solórzano, J.E.; Cruz, C.D.; Arenz, B.E.; Malvick, D.K.; Kleczewski, N.M. Tar spot of corn: A diagnostic and methods guide. Plant Health Prog. 2023, 24, 117–122. [Google Scholar] [CrossRef]
- Valle-Torres, J.; Ross, T.J.; Plewa, D.; Avellaneda, M.C.; Check, J.; Chilvers, M.I.; Cruz, A.P.; Dalla Lana, F.; Groves, C.; Gongora-Canul, C.; et al. Tar spot: An understudied disease threatening corn production in the Americas. Plant Dis. 2020, 104, 2541–2550. [Google Scholar] [CrossRef]
- Groves, C.L.; Kleczewski, N.M.; Telenko, D.E.; Chilvers, M.I.; Smith, D.L. Phyllachora maydis ascospore release and germination from overwintered corn residue. Plant Health Prog. 2020, 21, 26–30. [Google Scholar] [CrossRef]
- Kleczewski, N.M.; Donnelly, J.; Higgins, R. Phyllachora maydis, causal agent of tar spot on corn, can overwinter in northern Illinois. Plant Health Prog. 2019, 20, 178. [Google Scholar] [CrossRef]
- Lingg, A.J.; Donaldson, M.D. Biotic and abiotic factors affecting stability of Beauveria bassiana conidia in soil. J. Invertebr. Pathol. 1981, 38, 191–200. [Google Scholar] [CrossRef]
- Mondal, S.N.; Hyakumachi, M. Biological characteristics of chlamydospores of Fusarium solani f. sp. phaseoli. Mycoscience 1998, 39, 75–79. [Google Scholar]
- Eastburn, D.M.; Gubler, W.D. Effects of temperature and wetness duration on the viability of Colletotrichum acutatum conidia. Phytopathology 1992, 82, 1032–1035. [Google Scholar]
- Beare, M.H.; Hendrix, P.F.; Cabrera, M.L.; Coleman, D.C. Aggregate-protected and unprotected organic matter pools in conventional- and no-tillage soils. Soil Sci. Soc. Am. J. 1992, 56, 782–787. [Google Scholar] [CrossRef]
- Wise, K.; Mueller, D. Are Fungicides no Longer just for Fungi? An Analysis of Foliar Fungicide Use in Corn. APSnet Features. 2011. Available online: https://www.apsnet.org/edcenter/apsnetfeatures/Pages/fungicide.aspx (accessed on 3 June 2025).
- Amon, K.; Wade, T.; Mészáros, A.; Rumenik, T. A review of fresh market sweet corn production and price trends from 2010 to 2022. EDIS 2024. [Google Scholar] [CrossRef]
- Li, Y. Calcareous Soils in Miami-Dade County. EDIS 2001. Available online: https://edis.ifas.ufl.edu/publication/TR004 (accessed on 3 June 2025).
- Mylavarapu, R.; Harris, W.; Hochmuth, G. Agricultural soils of Florida. EDIS 2016. [Google Scholar] [CrossRef]
- Bayabil, H.; Li, Y.; Olczyk, T.; Liu, G. Sweet Corn Production in Miami-Dade County, Florida. EDIS 2022. Available online: https://edis.ifas.ufl.edu/publication/TR013 (accessed on 3 June 2025).
- USDA National Agricultural Statistical Service. The State of Agriculture in Florida 2022 USDA AG Census Update. 2024. Available online: https://ccmedia.fdacs.gov/content/download/118013/file/2022-florida-usda-census-of-agriculture-update (accessed on 3 June 2025).
- Mathur, S.P.; Lévesque, M.P. Relationship between acid phosphatase activities of soils and biochemical parameters of fungal decomposition. Plant Soil 1983, 74, 141–149. [Google Scholar] [CrossRef]
- Brooks, M.; Bolker, B.; Kristensen, K.; Maechler, M.; Magnusson, A.; McGillycuddy, M.; Skaug, H.; Nielsen, A.; Berg, C.; van Bentham, K. Package ‘glmmtmb’. 2023. Available online: https://cran.r-project.org/web/packages/glmmTMB/index.html (accessed on 3 June 2025).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://www.john-fox.ca/Companion/ (accessed on 3 June 2025).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2018. Available online: https://cran.r-project.org/web/packages/DHARMa/index.html (accessed on 3 June 2025).
- Lenth, R.; Lenth, M.R. Package ‘lsmeans’. Am. Stat. 2018, 34, 216–221. Available online: https://cran.r-project.org/web/packages/lsmeans/index.html (accessed on 3 June 2025).
- Huang, F.; Ding, X.; Li, W.; Jia, H.; Wei, X.; Zhao, X. The effect of temperature on the decomposition of different parts of maize residues in a solonchak. Catena 2021, 201, 105207. [Google Scholar] [CrossRef]
- Huang, F.; Zhang, W.; Xue, L.; Razavi, B.; Zamanian, K.; Zhao, X. The microbial mechanism of maize residue decomposition under different temperature and moisture regimes in a Solonchak. Sci. Rep. 2025, 15, 2215. [Google Scholar] [CrossRef]
- Fanin, N.; Mooshammer, M.; Sauvadet, M.; Meng, C.; Alvarez, G.; Bernard, L.; Bertr, I.; Blagodatskaya, E.; Bon, L.; Fontaine, S.; et al. Soil enzymes in response to climate warming: Mechanisms and feedbacks. Funct. Ecol. 2022, 36, 1378–1395. [Google Scholar] [CrossRef]
- Kirchman, D.L. Degradation of organic matter. In Processes in Microbial Ecology; Oxford University Press: Oxford, UK, 2018; pp. 113–129. [Google Scholar] [CrossRef]
- Donhauser, J.; Qi, W.; Bergk-Pinto, B.; Frey, B. High temperatures enhance the microbial genetic potential to recycle C and N from necromass in high-mountain soils. Glob. Change Biol. 2020, 27, 1365–1386. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, F.M.; Torn, M.S.; Trumbore, S.E. Warming accelerates decomposition of decades-old carbon in forest soils. Proc. Natl. Acad. Sci. USA 2012, 109, E1753–E1761. [Google Scholar] [CrossRef] [PubMed]
- Zuccarini, P.; Asensio, D.; Ogaya, R.; Sardans, J.; Peñuelas, J. Effects of seasonal and decadal warming on soil enzymatic activity in a P-deficient Mediterranean shrubland. Glob. Change Biol. 2020, 26, 3698–3714. [Google Scholar] [CrossRef]
- Gesch, R.W.; Reicosky, D.C.; Gilbert, R.A.; Morris, D.R. Influence of tillage and plant residue management on respiration of a Florida Everglades Histosol. Soil Tillage Res. 2007, 92, 156–166. [Google Scholar] [CrossRef]
- Lohila, A.; Aurela, M.; Regina, K.; Laurila, T. Soil and total ecosystem respiration in agricultural fields: Effect of soil and crop type. Plant Soil 2003, 251, 303–317. [Google Scholar] [CrossRef]
- Savabi, M.R. Determining soil water characteristics for application of WEPPmodel in South Florida. Trans. ASAE 2001, 44, 59. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S. Tropical soil biology and fertility: A handbook of methods. Soil Sci. 1994, 157, 265. [Google Scholar] [CrossRef]
- Góngora-Canul, C.; Jiménez-Beitia, F.E.; Puerto-Hernández, C.; Avellaneda, M.C.; Kleczewski, N.; Telenko, D.E.P.; Shim, S.; Solórzano, J.E.; Goodwin, S.B.; Scofield, S.R.; et al. Assessment of symptom induction via artificial inoculation of the obligate biotrophic fungus Phyllachora maydis (Maubl.) on corn leaves. BMC Res. Notes 2023, 16, 69. [Google Scholar] [CrossRef]
- Solórzano, J.E.; Subbaiah, A.; Floyd, C.M.; Kleczewski, N.M.; Malvick, D.K. Foliar inoculation of Phyllachora maydis into corn induces infection and local spread in field environments. Plant Dis. 2024, 108, 2672–2676. [Google Scholar] [CrossRef]
- Parbery, D. Studies on graminicolous species of Phyllachora Fckl. I. Ascospores–their liberation and germination. Aust. J. Bot. 1963, 11, 117. [Google Scholar] [CrossRef]
- Telenko, D.; Chilvers, M.; Kleczewski, N.; Mueller, D.; Plewa, D.; Robertson, A.; Smith, D.; Tenuta, A.; Wise, K. An Overview of Tar Spot; Crop Protection Network: Ames, IA, USA, 2020. [Google Scholar] [CrossRef]
- Rocco da Silva, C.; Check, J.; MacCready, J.S.; Alakonya, A.E.; Beiriger, R.; Bissonnette, K.M.; Collins, A.; Cruz, C.D.; Esker, P.D.; Goodwin, S.B.; et al. Recovery plan for tar spot of corn, caused by Phyllachora maydis. Plant Health Prog. 2021, 22, 596–616. [Google Scholar] [CrossRef]
- Ross, T.J.; Chilvers, M.I.; Byrne, A.M.; Smith, D.L.; Mueller, B.; Shim, S.; Telenko, D.E.P. Effects of disease resistance and fungicide application on management of tar spot on corn in the North Central United States. Plant Health Prog. 2023, 24, 439–444. [Google Scholar] [CrossRef]
- Moura, V.A.S.; Ferreira, L.C.; Resende, M.F.; Xavier, K.V. The history of tar spot on maize in South Florida: A continuous survey. Phytopathology 2023, 113, S3.1–S3.197. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Initial Germination (%) | Germination Decline (%) | Odds Ratio (%) a | Odds Ratio SE b | p-Values c |
|---|---|---|---|---|---|
| Muck | 10.9 | 49.9 | 50.1 | 30.1 | 0.2501 |
| Rocky | 13.4 | 88.9 | 11.1 | 11.9 | 0.0403 |
| 4 °C | 44.2 | 97.3 | 42.4 | 14.4 | 0.0113 |
| 23 °C | 20.6 | 57.3 | 2.7 | 2.0 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, V.A.S.; Ferreira, L.C.; Resende, M.F.R.; Xavier, K.V. Tar Spot on Maize: Impact of Soil Types and Environmental Conditions on the Survival of Phyllachora maydis in the Subtropical Climate of Florida. J. Fungi 2025, 11, 443. https://doi.org/10.3390/jof11060443
Moura VAS, Ferreira LC, Resende MFR, Xavier KV. Tar Spot on Maize: Impact of Soil Types and Environmental Conditions on the Survival of Phyllachora maydis in the Subtropical Climate of Florida. Journal of Fungi. 2025; 11(6):443. https://doi.org/10.3390/jof11060443
Chicago/Turabian StyleMoura, Vitor A. S., Larissa C. Ferreira, Marcio F. R. Resende, and Katia V. Xavier. 2025. "Tar Spot on Maize: Impact of Soil Types and Environmental Conditions on the Survival of Phyllachora maydis in the Subtropical Climate of Florida" Journal of Fungi 11, no. 6: 443. https://doi.org/10.3390/jof11060443
APA StyleMoura, V. A. S., Ferreira, L. C., Resende, M. F. R., & Xavier, K. V. (2025). Tar Spot on Maize: Impact of Soil Types and Environmental Conditions on the Survival of Phyllachora maydis in the Subtropical Climate of Florida. Journal of Fungi, 11(6), 443. https://doi.org/10.3390/jof11060443