
Isolation and Identification of Colletotrichum nymphaeae as a Causal Agent of Leaf Spot on Rhododendron hybridum Ker Gawl and Its Effects on the Ultrastructure of Host Plants

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Disease Investigation of R. hybridum Ker Gawl
2.2. Pathogen Isolation and Purification
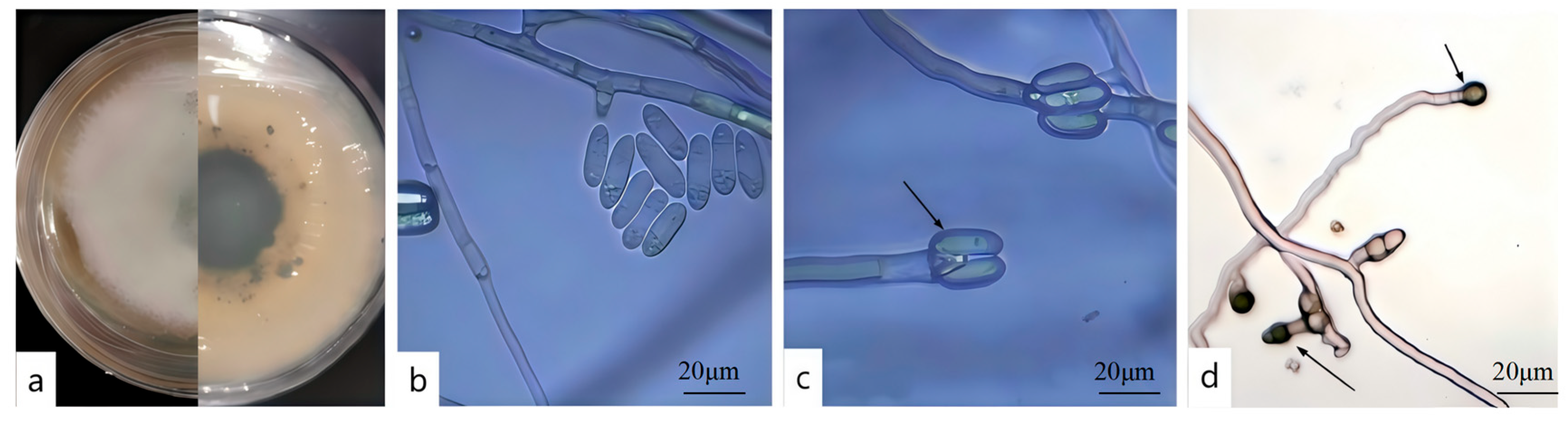
2.3. Pathogen Morphological Identification
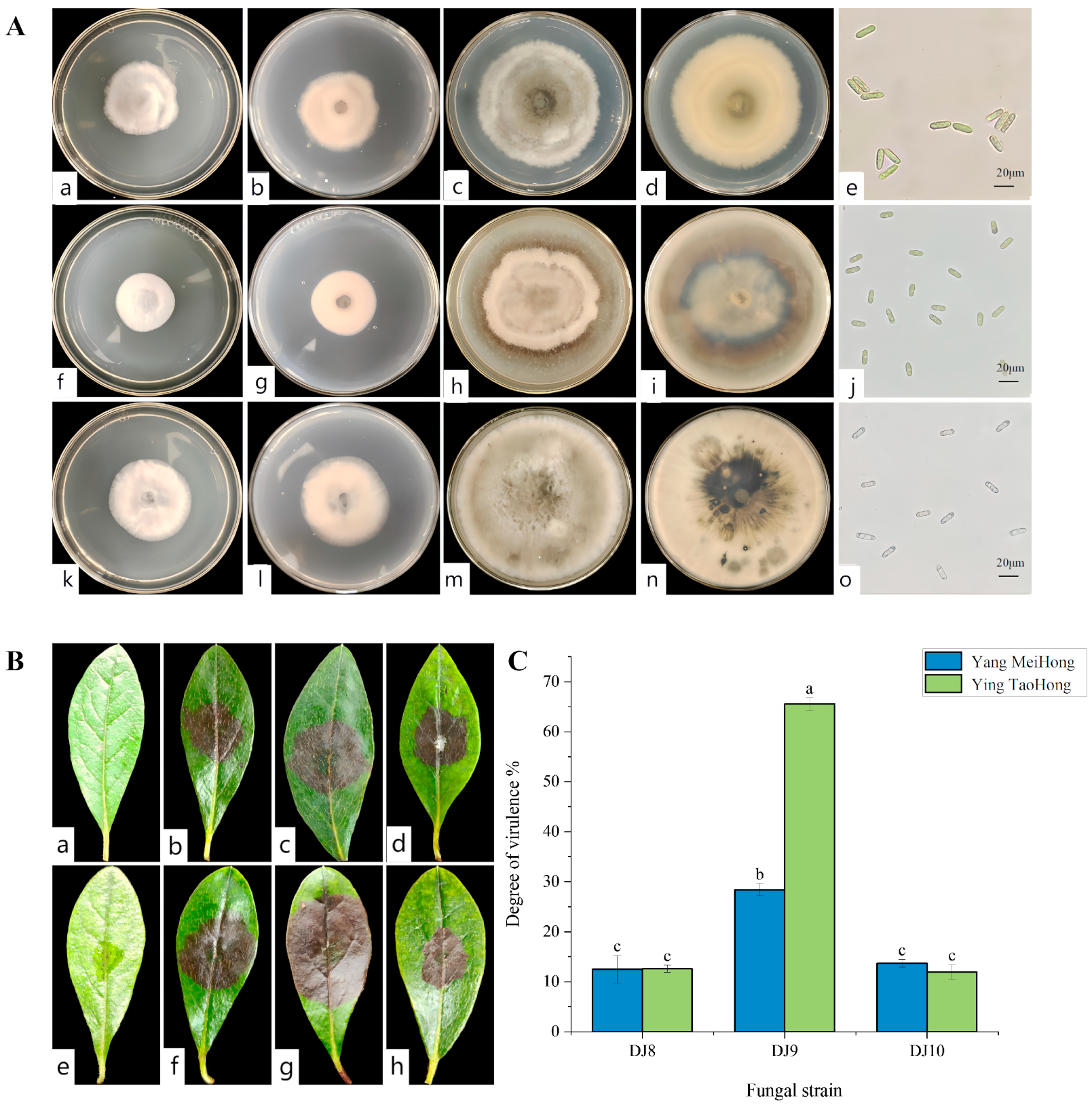
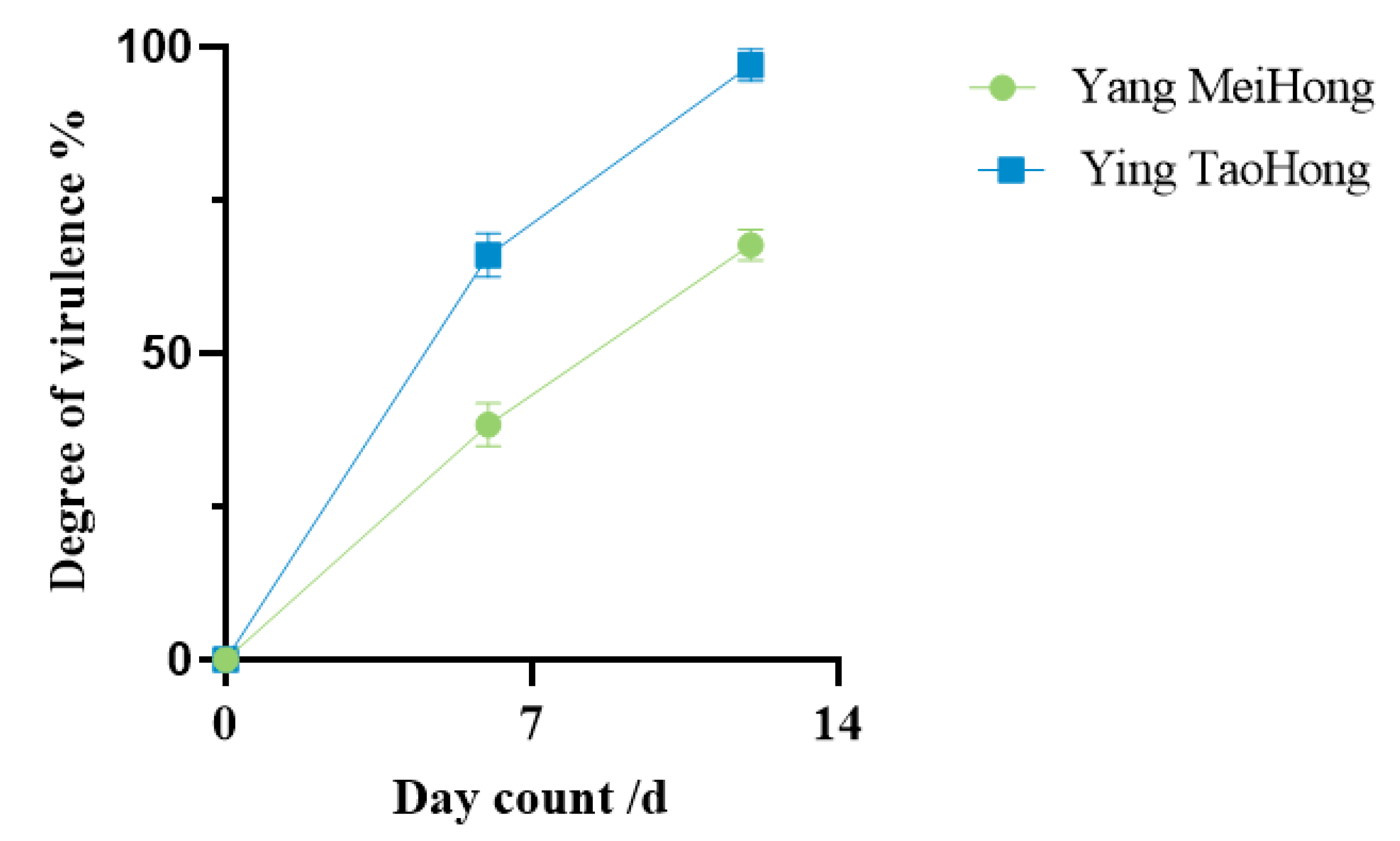
2.4. Pathogen Virulence Determination
2.5. Pathogen Molecular Biological Identification
2.6. Research on the Biological Characteristics of the Pathogen
2.6.1. Effect of Temperature on Mycelial Growth
2.6.2. Effect of pH on Mycelial Growth [41]
2.6.3. Effect of Light on Mycelial Growth
2.6.4. Carbon Source Screening Test
2.6.5. Nitrogen Source Screening Test
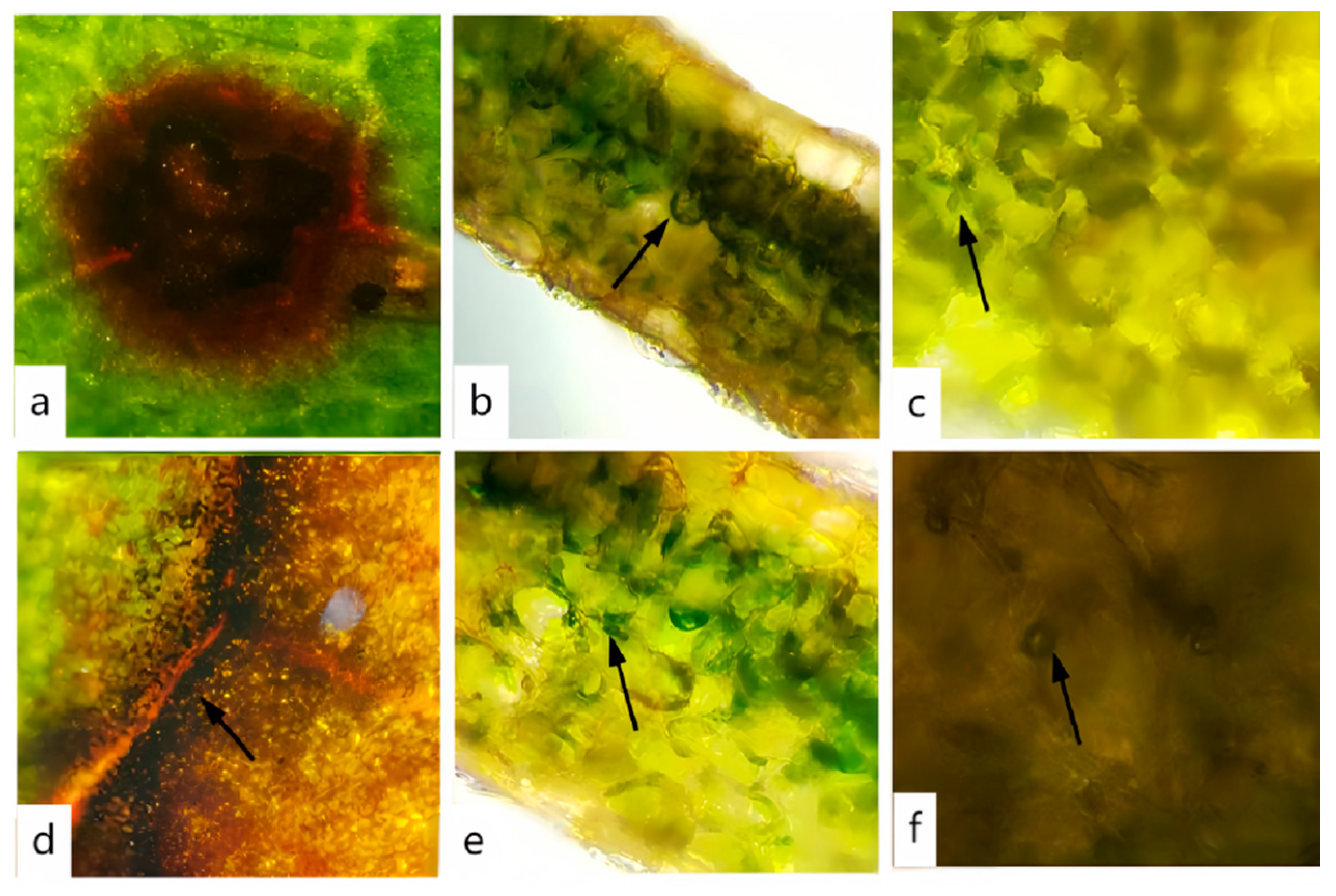
2.7. TEM Observation of Rhododendron Leaves Infected by Pathogen
- (1)
- Sample collection: Select mature Rhododendron leaves, ensuring the main vein is avoided. Cut a small leaf piece (1 mm × 1 mm) from the midpoint where the midrib intersects with the leaf. Fix the sample overnight in 2.5% glutaraldehyde for pre-fixation, keep it at room temperature for 2 h, and then store it at 4 °C in a refrigerator.
- (2)
- Fixation: Before observation, rinse the samples three times with 0.1 mol/L phosphate buffer solution (pH 7.4), with each rinse lasting 15 min. Then, fix the samples in a 1% osmium tetroxide solution for 2 h. After fixation, rinse the samples 4 times with 0.1 mol/L phosphate buffer solution, with each rinsing lasting 15 min.
- (3)
- Dehydration: Perform stepwise dehydration using ethanol solutions of increasing concentrations: 50%, 70%, 80%, 90%, 95%, and 100%. Each dehydration step should last for 15 min.
- (4)
- Embedding: Prepare a mixed solution by combining acetone and Epon 618 epoxy resin embedding agent at a 1:1 ratio. Allow the samples to undergo infiltration overnight. Afterward, proceed with polymerization at 60 °C for 48 h. Once polymerization is complete, use an ultramicrotome to cut ultrathin sections with a thickness of 60 to 80 nm.
- (5)
- Staining: Perform double staining using a saturated aqueous solution of 2% uranyl acetate followed by lead citrate. Each staining step should last for 15 min to enhance contrast for electron microscopy observation.
- (6)
- Observation: Examine the prepared ultrathin sections under a transmission electron microscope (JEM-1400Flash) to assess the cellular structure of the Rhododendron leaves. Observe for signs of plasmolysis and analyze ultrastructural changes in organelles such as chloroplasts, mitochondria, and the nucleus. Capture images for further analysis and documentation of structural alterations. The JEM-1400Flash device was produced by JEOL Ltd. in Tokyo, Japan.
3. Results
3.1. Disease Investigation and Sampling
3.2. Screening of Highly Virulent Pathogenic Fungi and Their Morphological Characteristics
3.3. Virulence Determination
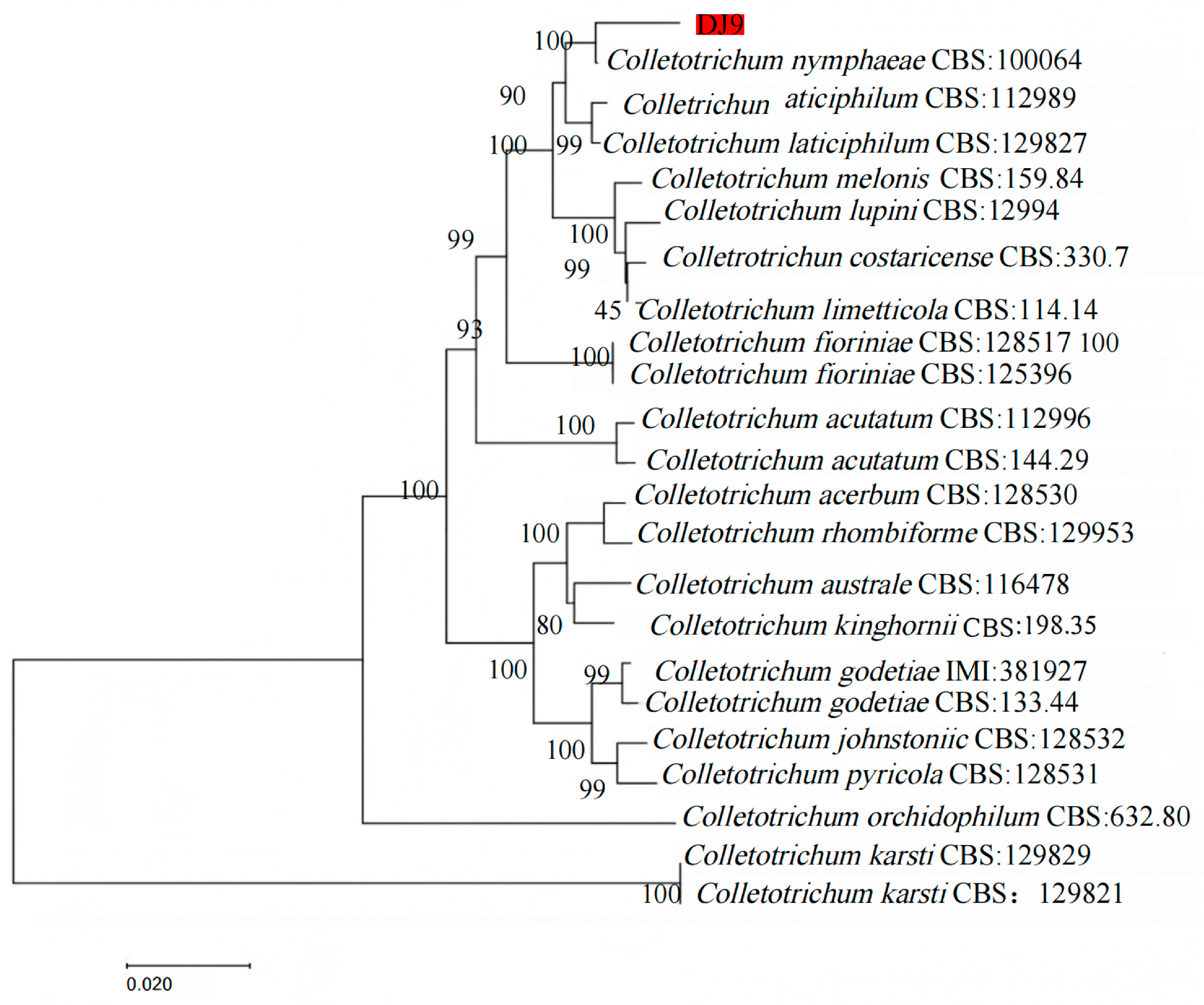
3.4. Molecular Phylogenetic Tree Analysis of Pathogens
3.5. Biological Characteristics of the Pathogen
3.5.1. Effect of Temperature on the Growth Rate of the Pathogen
3.5.2. Effect of pH on the Growth Rate of the Pathogen
3.5.3. Effect of Light Conditions on the Growth Rate of the Pathogen
3.5.4. Effect of Different Carbon Sources on the Growth Rate of the Pathogen
3.5.5. Effect of Different Nitrogen Sources on the Growth Rate of the Pathogen
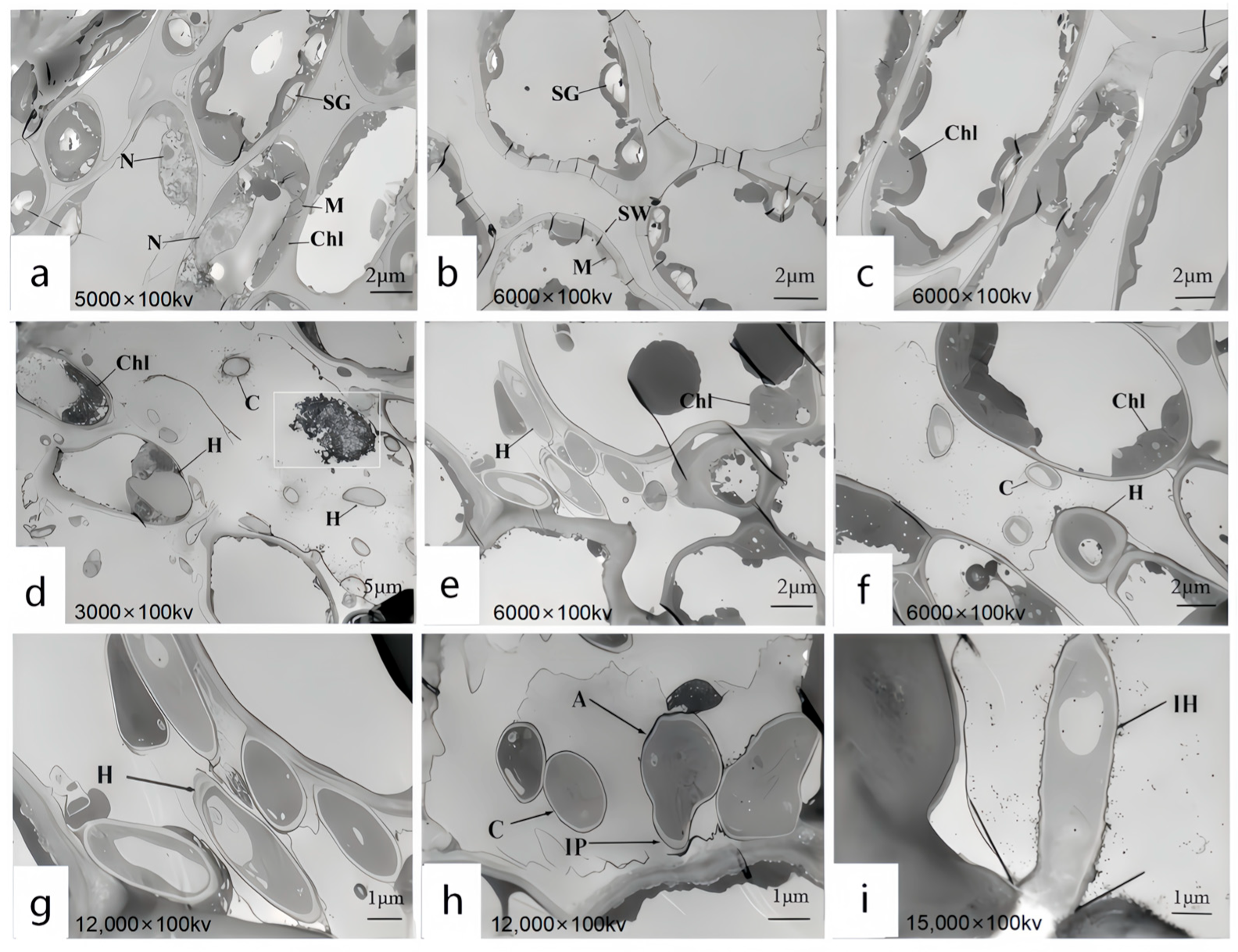
3.6. Ultrastructural Observation of the Host Under Pathogen Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kowalik, M. Fungi and fungi-like Oomycetes isolated from affected leaves of rhododendron. Acta Mycol. 2008, 43, 21–27. [Google Scholar] [CrossRef]
- Xu, J.; Luo, H.; Nie, S.; Zhang, R.G.; Mao, J.F. The complete mitochondrial and plastid genomes of Rhododendron simsii, an important parent of widely cultivated azaleas. Mitochondrial DNA Part B 2021, 6, 1197–1199. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, Y.; Gong, S.; Liu, X.; Tang, M.; Tang, J.; Zhang, X. The Preliminary Analysis of Flavonoids in the Petals of Rhododendron delavayi, Rhododendron agastum and Rhododendron irroratum Infected with Neopestalotiopsis clavispora. Int. J. Mol. Sci. 2024, 25, 9605. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, L.; Wang, X.; Zhang, R.; Jin, G.; Hu, Y.; Wang, J. Evolutionary history of two evergreen Rhododendron species as revealed by chromosome-level genome assembly. Front. Plant Sci. 2023, 14, 1123707. [Google Scholar] [CrossRef]
- Chao, X.; Sun, G.; Zhao, H.; Li, M.; He, D. Identification of apple tree leaf diseases based on deep learning models. Symmetry 2020, 12, 1065. [Google Scholar] [CrossRef]
- Masunaka, A.; Ohtani, K.; Peever, T.L.; Timmer, L.W.; Tsuge, T.; Yamamoto, M.; Akimitsu, K. An isolate of Alternaria alternata that is pathogenic to both tangerines and rough lemon and produces two host-selective toxins, ACT-and ACR-toxins. Phytopathology 2005, 95, 241–247. [Google Scholar] [CrossRef]
- Copes, W.E. Spread potential of binucleate Rhizoctonia from nursery propagation floors to trays containing azalea stem cuttings and sanitary control options. Plant Dis. 2015, 99, 842–847. [Google Scholar] [CrossRef]
- Rivera, M.C.; Wright, E.R. First report of Azalea petal blight caused by Pestalotiopsis guepini in argentina. Plant Dis. 2000, 84, 100. [Google Scholar] [CrossRef]
- Garibaldi, A.; Bertetti, D.; Vinnere, O.; Gullino, M.L. Presence of Colletotrichum acutatum causing leaf spot on Azalea japonica in Italy. Plant Dis. 2004, 88, 572. [Google Scholar] [CrossRef]
- Mrazkova, M.; Cerny, K.; Gabrielova, S.; Tomsovsky, M. First report of leaf spot, shoot blight, and stem and collar canker of Rhododendron spp. caused by Phytophthora citricola in the Czech Republic. Plant Dis. 2007, 91, 1515. [Google Scholar] [CrossRef]
- Álvarez, L.A.; Pérez-Sierra, A.; García-Jiménez, J.; Abad-Campos, P.; Landeras, E.; Alzugaray, R. First report of leaf spot and twig blight of Rhododendron spp. caused by Phytophthora hibernalis in Spain. Plant Dis. 2007, 91, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Inghelbrecht, S.; Gehesquière, B.; Heungens, K. First report of Calonectria leaf spot caused by Calonectria colhounii (Anamorph Cylindrocladium colhounii) on Rhododendron in Belgium. Plant Dis. 2011, 95, 1477. [Google Scholar] [CrossRef] [PubMed]
- Yakabe, L.E.; Soriano, M.C.; Negrete, M.A. First report of leaf spot caused by Phytophthora taxon Pgchlamydo on evergreen nursery stock in California. Plant Dis. 2012, 96, 1691. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef]
- Zhu, W.; Hu, S.; Zhou, Z.; Tang, X.; Wu, X.; Li, Z.; Ding, H. First report of leaf spot on Rhododendron delavayi caused by Pestalotiopsis scoparia in China. Plant Dis. 2023, 107, 1626. [Google Scholar] [CrossRef]
- Liu, Y.L.; Yang, J.; Lian, S.W.; Xie, Z.; Luo, J.Q.; Cheng, P.Y.; Li, Z.J. Pseudocercospora rhododendricola causing leaf dark spot on Rhododendron pulchrum by the first phylogenetic analyses. Plant Dis. 2023, 107, 1631. [Google Scholar] [CrossRef]
- Jones, R.K.; Benson, D.M.; Rhododendron Diseases. JARS 1984. Available online: https://scholar.lib.vt.edu/ejournals/JARS/v38n3/v38n3-jones.html (accessed on 21 March 2025).
- Do Vale, A.; Cabanes, D.; Sousa, S. Bacterial toxins as pathogen weapons against phagocytes. Front. Microbiol. 2016, 7, 42. [Google Scholar] [CrossRef]
- Hodge, S.; Powell, G. Complex interactions between a plant pathogen and insect parasitoid via the shared vector-host: Consequences for host plant infection. Oecologia 2008, 157, 387–397. [Google Scholar] [CrossRef]
- Lorrai, R.; Ferrari, S. Host cell wall damage during pathogen infection: Mechanisms of perception and role in plant-pathogen interactions. Plants 2021, 10, 399. [Google Scholar] [CrossRef]
- Vidhyasekaran, P.; Ponmalar, T.R.; Samiyappan, R. Host-specific toxin production by Rhizoctonia solani, the rice sheath blight pathogen. Phytopathology 1997, 87, 1258–1263. [Google Scholar] [CrossRef]
- Park, P.; Ikeda, K.I. Ultrastructural analysis of responses of host and fungal cells during plant infection. J. Gen. Plant Pathol. 2008, 74, 2–14. [Google Scholar] [CrossRef]
- Hu, G.G.; Linning, R.; Bakkeren, G. Sporidial mating and infection process of the smut fungus, Ustilago hordei, in susceptible barley. Can. J. Bot. 2003, 80, 1103–1114. [Google Scholar] [CrossRef]
- Martinez, C.; Roux, C.; Jauneau, A.; Dargent, R. The biological cycle of Sporisorium reilianum f.sp. zeae: An overview using microscopy. Mycologia 2002, 94, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Hongmin, G.; Jialin, N.; Bing, Y. The influence of the pathogenic factors of corn stalk rot pathogens to the ultrastructure of maize. J. Maize Sci. 2006, 14, 147–149. [Google Scholar]
- Zhang, W.; Guo, M.; Yang, W.; Liu, Y.; Wang, Y.; Chen, G. The role of cell wall polysaccharides disassembly and enzyme activity changes in the softening process of Hami melon (Cucumis melo L.). Foods 2022, 11, 841. [Google Scholar] [CrossRef]
- Mou, X.L.; Li, X.X.; Shi, G.Y.; Wang, W.Z.; Li, M.Q.; Zhang, A.J. Identification of pathogens causing Fusarium wilt of Lanzhou lily, and the observation of cell ultrastructures in the infected bulb scales. J. Plant Prot. 2022, 49, 1111–1118. Available online: https://m.doc88.com/p-17739675171613.html (accessed on 21 March 2025).
- Baka, Z.A. Light and electron microscopy of the micromorphology and development of pycniospores and aeciospores of the sunflower rust, Puccinia helianthi. Micron 2025, 189, 103733. [Google Scholar] [CrossRef]
- Zhao, S.; Hu, D. Ultrastructure and cytochemistry of barley mesophyll attacked by Blumeria graminis f. sp. hordei. Acta Phytopathol. Sin. 2003, 33, 444–448. [Google Scholar]
- Baka, Z.A. Ultrastructural studies of the host (Senecio aegyptius L.) responses after the infection by the pathogenic rust fungus, Puccinia lagenophorae Cooke. Arch. Phytopathol. Plant Prot. 2023, 56, 234–255. [Google Scholar] [CrossRef]
- Pandey, A.; Kumar, B.; Verma, C.; Johnson, D.; Singh, E. A novel disease rating scale for assessing severity of fungal leaf spots in horticultural crops. Plant Dis. 2023, 107, 1234–1242. [Google Scholar]
- Johnson, D.A.; Carris, L.M.; Rogers, J.D. Morphological and molecular characterization of Colletotrichum nymphaeae and C. nupharicola sp. nov. on water-lilies (Nymphaea and Nuphar). Mycol. Res. 1997, 101, 641–649. [Google Scholar] [CrossRef]
- Wang, W.; Liang, X.; Zhang, R.; Gleason, M.L.; Sun, G. Liquid shake culture overcomes solid plate culture in inducing conidial production of Colletotrichum isolates. Australas. Plant Pathol. 2017, 46, 285–287. [Google Scholar] [CrossRef]
- Chen, J.; Xu, L.L.; Liu, B.; Liu, X.Z. Taxonomy of Dactylella complex and Vermispora. I. Generic concepts based on morphology and ITS sequences data. Fungal Divers. 2007, 26, 73–83. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Wang, R.Y.; Gao, B.; Li, X.H.; Ma, J.; Chen, S.L. First report of Fusarium solani causing Fusarium root rot and stem canker on storage roots of sweet potato in China. Plant Dis. 2014, 98, 160. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.A.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef]
- Mtimet, N.; Guégan, S.; Durand, L.; Mathot, A.G.; Venaille, L.; Leguérinel, I.; Coroller, L.; Couvert, O. Effect of pH on Thermoanaerobacterium thermosaccharolyticum DSM 571 growth, spore heat resistance and recovery. Food Microbiol. 2016, 55, 64–72. [Google Scholar] [CrossRef]
- Wang, Y.X.; Xu, X.W.; Cai, F.; Huang, F.X.; Chen, W.S.; Wang, Q.Z. First report of Colletotrichum nymphaeae causing walnut anthracnose in China. Plant Dis. 2022, 106, 2991. [Google Scholar] [CrossRef] [PubMed]
- Antelmi, I.; Sion, V.; Nigro, F. First report of Colletotrichum nymphaeae on olive in Italy. Plant Dis. 2019, 103, 765. [Google Scholar] [CrossRef]
- Mulvey, M.A.; Hultgre, S.J. Bacterial Spelunkers. Science 2000, 289, 732–733. [Google Scholar] [CrossRef] [PubMed]
- Al-Askar, A.A.; Baka, Z.A.; Rashad, Y.M.; Ghoneem, K.M.; Abdulkhair, W.M.; Hafez, E.E.; Shabana, Y.M. Evaluation of Streptomyces griseorubens E44G for the bio-control of Fusarium oxysporum f. sp. lycopersici: Ultrastructural and cytochemical investigations. Ann. Microbiol. 2015, 65, 1815–1824. [Google Scholar] [CrossRef]
- Mohamed, M.S.; Saleh, A.M.; Abdel-Farid, I.B.; El-Naggar, S.A. Growth, hydrolases and ultrastructure of Fusarium oxysporum as affected by phenolic rich extracts from several xerophytic plants. Pestic. Biochem. Physiol. 2017, 141, 57–64. [Google Scholar] [CrossRef]
- Coates, L.M.; Muirhead, I.F.; Irwin, J.A.G.; Gowanlock, D.H. Initial infection processes by Colletotrichum gloeosporioides on avocado fruit. Mycol. Res. 1993, 97, 1363–1370. [Google Scholar] [CrossRef]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum Species with Curved Conidia from Herbaceous Hosts. Fungal Divers. 2009, 39, 45–87. [Google Scholar]
- Zhang, L.; Meng, F.; Ge, W.; Ren, Y.; Bao, H.; Tian, C. Effects of colletotrichum gloeosporioides and poplar secondary metabolites on the composition of poplar phyllosphere microbial communities. Microbiol. Spectr. 2023, 11, e04603-22. [Google Scholar] [CrossRef]
- Van der Aa, H.A. A leaf spot disease of Nymphaea alba in the Netherlands. Neth. J. Plant Pathol. 1978, 84, 109–115. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef]
- Velho, A.C.; Stadnik, M.J.; Casanova, L.; Mondino, P.; Alaniz, S. First report of Colletotrichum nymphaeae causing apple bitter rot in southern Brazil. Plant Dis. 2014, 98, 567. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.C.; Zeng, X.G.; Xiang, F.Y.; Ren, L.; Chen, F.Y.; Gu, Y.C. Distribution and characteristics of Colletotrichum spp. associated with anthracnose of strawberry in Hubei, China. Plant Dis. 2016, 100, 996–1006. [Google Scholar] [CrossRef]
- Luongo, L.; Galli, M.; Garaguso, I.; Petrucci, M.; Vitale, S. First report of Colletotrichum fioriniae and C. nymphaeae as Causal Agents of anthracnose on walnut in Italy. Plant Dis. 2022, 106, 327. [Google Scholar] [CrossRef]
- Wu, J.; Wang, H.; Fang, L.; Xie, X.; Wang, L. First report of Colletotrichum fructicola and Colletotrichum nymphaeae causing leaf spot on Rubus corchorifolius in Zhejiang province, China. Plant Dis. 2021, 105, 3746. [Google Scholar] [CrossRef]
- Garcia-Lopez, M.T.; Serrano, M.S.; Camiletti, B.X.; Gordon, A.; Estudillo, C.; Trapero, A.; Moral, J. Study of the competition between Colletotrichum godetiae and C. nymphaeae, two pathogenic species in olive. Sci. Rep. 2023, 13, 5344. [Google Scholar] [CrossRef]
- Liu, H.; Li, Y.; Li, X.; Liu, H.; Huang, J.; Zheng, L. First report of tobacco anthracnose caused by Colletotrichum nymphaeae in China. Plant Dis. 2023, 107, 2537. [Google Scholar] [CrossRef]
- Zaitlin, B.; Zehr, E.I.; Dean, R.A. Latent infection of peach caused by Colletotrichum gloeosporioides and Colletotrichum acutatum. Can. J. Plant Pathol. 2000, 22, 224–228. [Google Scholar] [CrossRef]
- Fan, R.; Liu, Y.; Bin, Y.; Huang, J.; Yi, B.; Tang, X.; Li, Y.; Cai, Y.; Yang, Z.; Yang, M. Identification of Colletotrichum aenigma as the new causal agent of leaf blight disease on Aucuba japonica Thunb, and screenings of effective fungicides for its sustainable management. Front. Microbiol. 2023, 14, 1222844. [Google Scholar] [CrossRef]
- Macioszek, V.K.; Chalamońska, K.; Oliwa, J.; Staszak, A.M.; Sobczak, M. Phenological, Physiological, and Ultrastructural Analyses of ‘Green Islands’ on Senescent Leaves of Norway Maple (Acer platanoides L.). Plants 2025, 14, 909. [Google Scholar] [CrossRef]
- Li, M.; Daygon, V.D.; Solah, V.; Dhital, S. Starch granule size: Does it matter? Crit. Rev. Food Sci. Nutr. 2023, 63, 3683–3703. [Google Scholar] [CrossRef]
- Cao, B.D.; Wu, W.; Cheng, H.; Wu, Y.S. Ultrastructure of leaf tissues of resistant or susceptible wheat exposed to toxin produced by Helminthosporium sativum. Acta Phytopathol. Sin. 1993, 23, 107–113. [Google Scholar]
- Türkkan, M.; Andolfi, A.; Zonno, M.C.; Erper, I.; Perrone, C.; Cimmino, A.; Evidente, A. Phytotoxins produced by Pestalotiopsis guepinii, the causal agent of hazelnut twig blight. Phytopathol. Mediterr. 2011, 50, 154–158. [Google Scholar] [CrossRef]
- van Kan, J.A. Licensed to kill: The lifestyle of a necrotrophic plant pathogen. Trends Plant Sci. 2006, 11, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Mares, D.J. A light and electron microscope study of the interaction of yellow rust Puccinia striiformis with a susceptible wheat cultivar. Ann. Bot. 1979, 43, 183–190. [Google Scholar] [CrossRef]
- Yang, H.; Luo, P. Changes in photosynthesis could provide important insight into the interaction between wheat and fungal pathogens. Int. J. Mol. Sci. 2021, 22, 8865. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Grade | Standard |
|---|---|
| 0 | no visible lesions |
| 1 | The area of lesions accounts for less than 5% of the leaf area |
| 3 | The area of lesions accounts for 6–10% of the leaf area |
| 5 | The area of lesions accounts for 11–20% of the leaf area |
| 7 | The area of lesions accounts for 21–40% of the leaf area |
| 9 | The area of lesions accounts for more than 40% of the leaf area |
| Gene | Primer | Sequence (5′-3′) | Length of DNA Fragment |
|---|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGG | 429 bp |
| ITS4 | TCCTCCGCTTATTGATATGC | ||
| ACT | ACT-512F | ATGTGCAAGGCCGGTTTCGC | 252 bp |
| ACT-783R | TACGAGTCCTTCTGGCCCAT | ||
| TUB2 | Btub2Fd | GTBCACCTYCARACCGGYCARTG | 750 bp |
| Btub4Rd | CCRGAYTGRCCRAARACRAAGTTGTC | ||
| CHS-1 | CHS-79F | TGGGGCAAGGATGCTTGGAAGAAG | 300 bp |
| CHS-354R | TGGAAGAACCATCTGTGAGAGAGTTG | ||
| GAPDH | GDF1 | GCCGTCAACGACCCCTTCATTGA | 150 bp |
| GDR1 | GGGTGGAGTCGTACTTGAGCATGT | ||
| HIS3 | CYLH3F | AGGTCCACTGGTGGCAAG | 500 bp |
| CYLH3R | AGCTGGATGTCCTTGGACTG |
| Survey Area | Number of Plants at Each Grade | Disease Index | Incidence Rate | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 3 | 5 | 7 | 9 | |||
| Kunming World Horticultural Expo Garden | 200 | 120 | 75 | 60 | 35 | 20 | 23.31% | 60.79% |
| Southwest Forestry University, Kunming | 0 | 1 | 5 | 4 | 30 | 20 | 78.88% | 100% |
| Kunming Cuihu Park | 20 | 20 | 40 | 50 | 10 | 10 | 40.74% | 86.66% |
| Species | Accession | Gene Bank | |||||
|---|---|---|---|---|---|---|---|
| ITS | GAPDH | CHS-1 | HIS3 | ACT | TUB2 | ||
| C. acerbum | CBS:128530 * | JQ948459 | JQ948790 | JQ949120 | JQ949450 | JQ949780 | JQ950110 |
| C. australe | CBS 116478 | JQ948455 | JQ948786 | JQ949116 | JQ949446 | JQ949776 | JQ950106 |
| C. acutatum | CBS 112996 | JQ005776 | JQ948677 | JQ005797 | JQ005818 | JQ005839 | JQ005860 |
| C. acutatum | CBS 144.29 | JQ948401 | JQ948732 | JQ949062 | JQ949392 | JQ949722 | JQ950052 |
| C. costaricense | CBS 330.75 * | JQ948180 | JQ948510 | JQ948841 | JQ949171 | JQ949501 | JQ949831 |
| C. fiorinae | CBS128517 * | JQ948292 | JQ948622 | JQ948953 | JQ949283 | JQ949613 | JQ949943 |
| C. fiorinae | CBS125396 | JQ948299 | JQ948629 | JQ948960 | JQ949290 | JQ949620 | JQ949950 |
| C. godetiae | IMI 381927 | JQ948438 | JQ948769 | JQ949099 | JQ949429 | JQ949759 | JQ950089 |
| C. godetiae | CBS133.44 * | JQ948402 | JQ948733 | JQ949063 | JQ949393 | JQ949723 | JQ950053 |
| C. johnstonii | CBS 128532 | JQ948444 | JQ948775 | JQ949105 | JQ949435 | JQ949765 | JQ950095 |
| C.karstii | CBS:129829 | JQ005189 | JQ005276 | JQ005363 | JQ005450 | JQ005537 | JQ005623 |
| C.karstii | CBS:130235 | JQ005190 | JQ005277 | JQ005364 | JQ005451 | JQ005538 | JQ005624 |
| C. kinghornii | CBS 198.35 * | JQ948454 | JQ948785 | JQ949115 | JQ949445 | JQ949775 | JQ950105 |
| C. laticiphilum | CBS 112989 | JQ948289 | JQ948619 | JQ948950 | JQ949280 | JQ949610 | JQ949940 |
| C. laticiphilum | CBS 129827 | JQ948290 | JQ948620 | JQ948951 | JQ949281 | JQ949611 | JQ949941 |
| C. limetticola | CBS 114.14 * | JQ948193 | JQ948523 | JQ948854 | JQ949184 | JQ949514 | JQ949844 |
| C. lupini | CBS 129944 | JQ948178 | JQ948508 | JQ948839 | JQ949169 | JQ949499 | JQ949829 |
| melonis | CBS 159.84 * | JQ948194 | JQ948524 | JQ948855 | JQ949185 | JQ949515 | JQ949845 |
| C. nymphaeae | CBS 100064 | JQ948224 | JQ948554 | JQ948885 | JQ949215 | JQ949545 | JQ949875 |
| C.orchidophilum | CBS632.80 * | JQ948151 | JQ948481 | JQ948812 | JQ949142 | JQ949472 | JQ949802 |
| C. pyricola | CBS 128531 | JQ948445 | JQ948776 | JQ949106 | JQ949436 | JQ949766 | JQ950096 |
| C. rhombiforme | CBS 129953 | JQ948457 | JQ948788 | JQ949118 | JQ949448 | JQ949778 | JQ950108 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Tian, Y.; Liu, J.; Li, H.; Lu, J.; Wang, M.; Liu, S. Isolation and Identification of Colletotrichum nymphaeae as a Causal Agent of Leaf Spot on Rhododendron hybridum Ker Gawl and Its Effects on the Ultrastructure of Host Plants. J. Fungi 2025, 11, 392. https://doi.org/10.3390/jof11050392
Sun Y, Tian Y, Liu J, Li H, Lu J, Wang M, Liu S. Isolation and Identification of Colletotrichum nymphaeae as a Causal Agent of Leaf Spot on Rhododendron hybridum Ker Gawl and Its Effects on the Ultrastructure of Host Plants. Journal of Fungi. 2025; 11(5):392. https://doi.org/10.3390/jof11050392
Chicago/Turabian StyleSun, Yajiao, Yunjing Tian, Jian Liu, Huali Li, Junjia Lu, Mengyao Wang, and Shuwen Liu. 2025. "Isolation and Identification of Colletotrichum nymphaeae as a Causal Agent of Leaf Spot on Rhododendron hybridum Ker Gawl and Its Effects on the Ultrastructure of Host Plants" Journal of Fungi 11, no. 5: 392. https://doi.org/10.3390/jof11050392
APA StyleSun, Y., Tian, Y., Liu, J., Li, H., Lu, J., Wang, M., & Liu, S. (2025). Isolation and Identification of Colletotrichum nymphaeae as a Causal Agent of Leaf Spot on Rhododendron hybridum Ker Gawl and Its Effects on the Ultrastructure of Host Plants. Journal of Fungi, 11(5), 392. https://doi.org/10.3390/jof11050392