
Nutritional and Therapeutic Potential of Stropharia rugosoannulata and Macrolepiota procera: From Composition to Health-Promoting Effect



Abstract
1. Introduction
2. Nutritional Composition
2.1. Moisture
2.2. Protein and Amino Acids
2.3. Carbohydrates
2.4. Lipids
2.5. Minerals
2.5.1. Macroelements
2.5.2. Trace Elements
2.6. Vitamins
3. Bioactive Compounds
3.1. Polysaccharides
3.2. Phenolic Compounds
3.3. Terpenes
3.4. Polypeptides and Enzymes
3.5. Other Bioactive Compounds
4. Preparation of Bioactive Compounds
4.1. Polysaccharide Extraction
4.2. Extraction of Other Bioactive Compounds
5. Omics Analysis
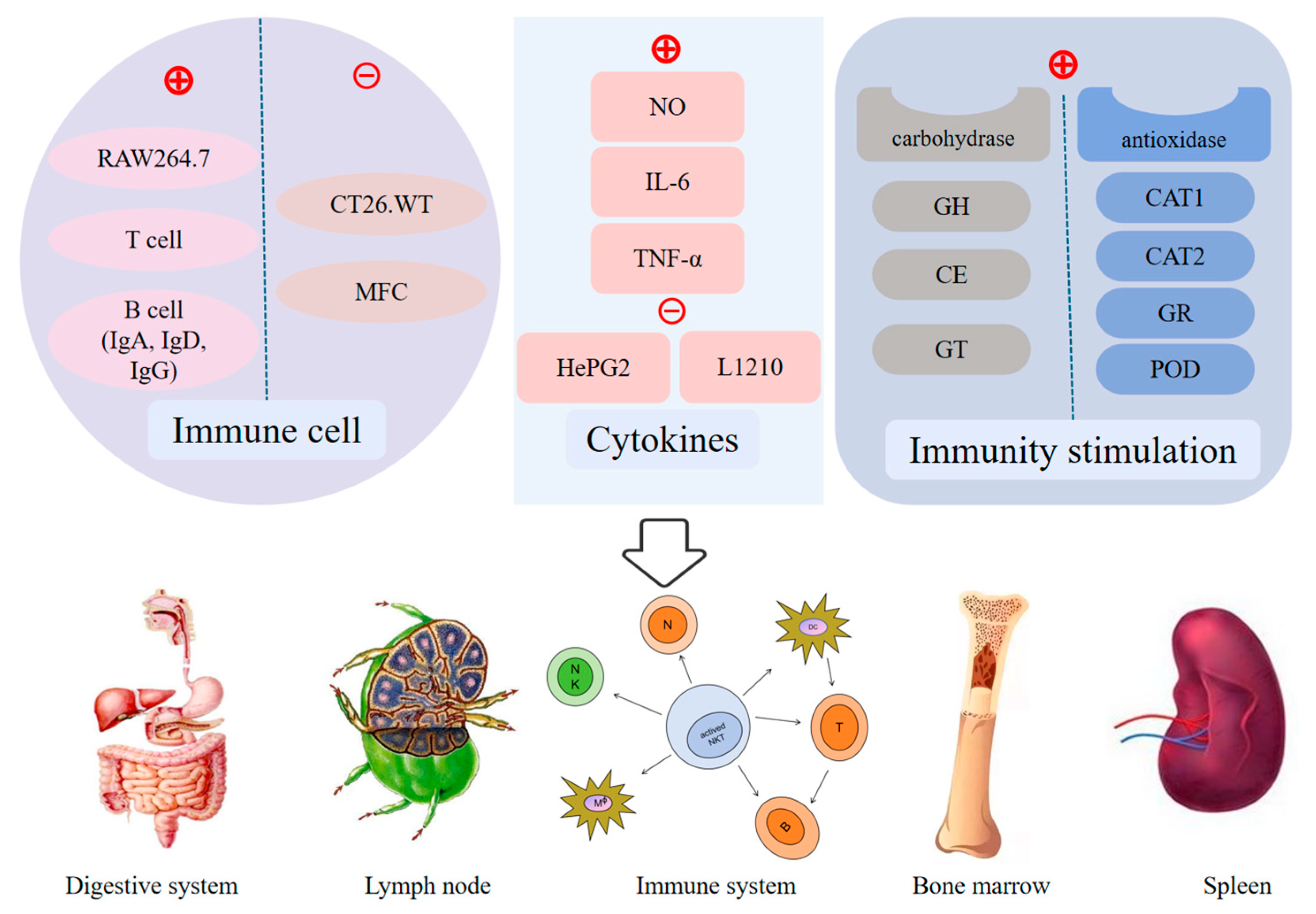
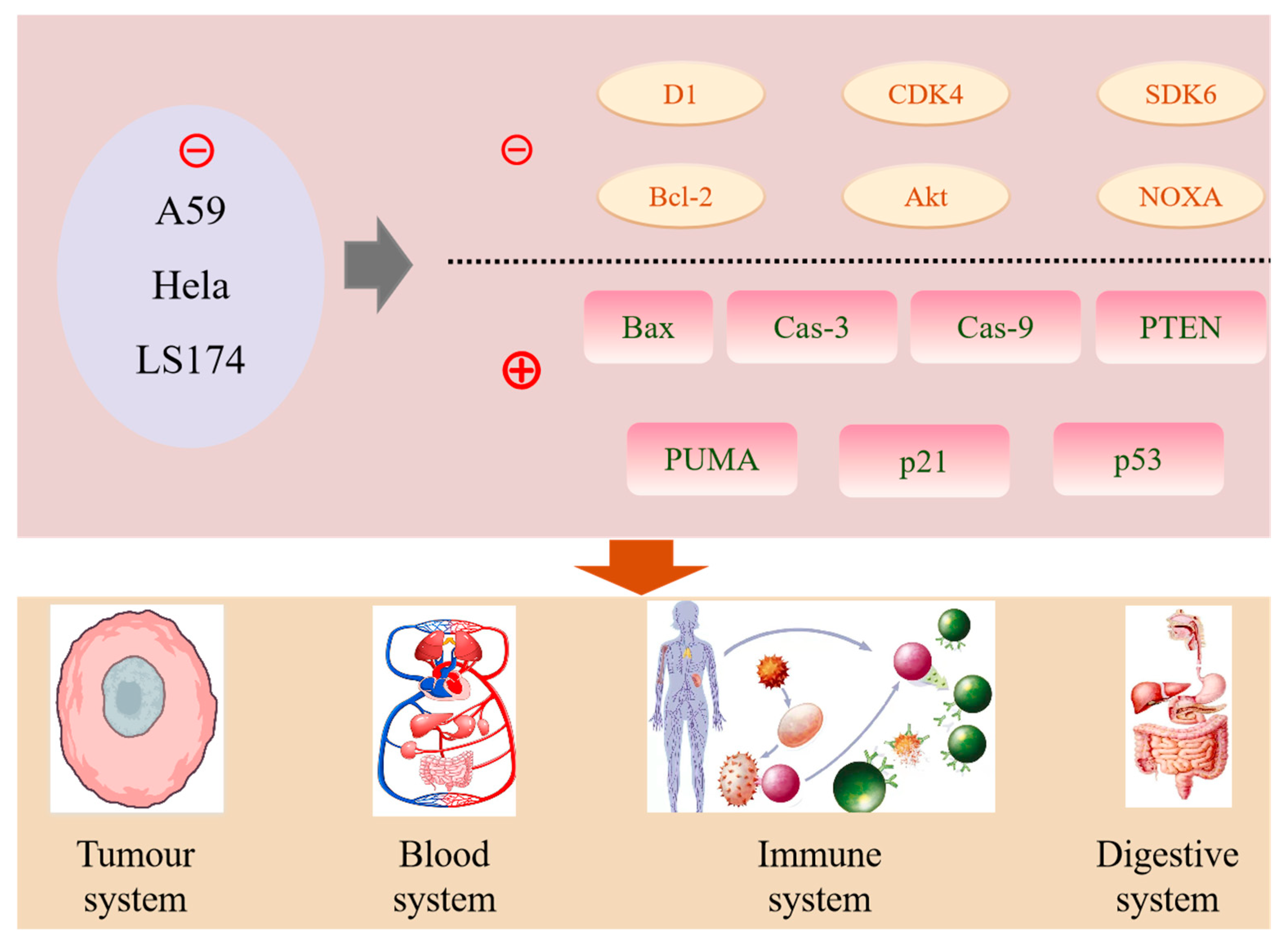
6. Bioactivities and Health-Promoting Benefits
6.1. Antioxidant Properties
6.2. Immunomodulatory Effects
6.3. Anticancer Properties
6.4. Antimicrobial Activity
6.5. Others
7. Safety and Toxicology
7.1. Heavy Metal Accumulation
7.2. Antinutritional Factors
7.3. Improper Storage
7.4. Mitigation Strategies
8. Conclusions and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- El-Ramady, H.; Abdalla, N.; Badgar, K.; Llanaj, X.; Törős, G.; Hajdú, P.; Eid, Y.; Prokisch, J. Edible mushrooms for sustainable and healthy human food: Nutritional and medicinal attributes. Sustainability 2022, 14, 4941. [Google Scholar] [CrossRef]
- Hamza, A.; Mylarapu, A.; Krishna, K.V.; Kumar, D.S. An insight into the nutritional and medicinal value of edible mushrooms: A natural treasury for human health. J. Biotechnol. 2024, 381, 86–99. [Google Scholar]
- Hu, Y.W.; Kakumyan, P.; Bandara, A.R.; Mortimer, P.E. The nutrition, cultivation and biotechnology of Stropharia rugosoannulata. Fungal Biotec 2021, 1, 13–25. [Google Scholar] [CrossRef]
- Dai, Y.C.; Tolgor, B.; Cui, B.K.; Qin, G.F. Illustrations of Medicinal Fungi in China; Northeast Forestry University Press: Harbin, China, 2013. [Google Scholar]
- Ma, L.F.; He, S.L.; Yang, J.J.; Sun, Y.B.; Yang, X.L.; Chang, Y.W. Effects of different Stropharia rugosoannulata culture medium on growth, yield and quality. North. Hortic. 2021, 22, 130–135. [Google Scholar]
- Kamra, D.N.; Zadražil, F. Influence of oxygen and carbon dioxide on lignin degradation in solid state fermentation of wheat straw with Stropharia rugosoannulata. Biotechnol. Lett. 1985, 7, 335–340. [Google Scholar]
- Castellet-Rovira, F.; Lucas, D.; Villagrasa, M.; Rodríguez-Mozaz, S.; Barceló, D.; Sarrà, M. Stropharia rugosoannulata and Gymnopilus luteofolius: Promising fungal species for pharmaceutical biodegradation in contaminated water. J. Environ. Manag. 2018, 207, 396–404. [Google Scholar] [CrossRef]
- Pozdnyakova, N.; Schlosser, D.; Dubrovskaya, E.; Balandina, S.; Sigida, E.; Grinev, V.; Turkovskaya, O. The degradative activity and adaptation potential of the litter-decomposing fungus Stropharia rugosoannulata. World J. Microbiol. Biotechnol. 2018, 34, 133. [Google Scholar] [CrossRef]
- Gong, S.; Chen, C.; Zhu, J.X.; Qi, G.Y.; Jiang, S.X. Effects of wine-cap Stropharia cultivation on soil nutrients and bacterial communities in forestlands of northern China. PeerJ 2018, 6, e5741. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Sharma, S.; Kumar, S. Cultivation of least-exploited commercial mushrooms. Mushroom Biol. Biotechnol. 2007, 213, 167–192. [Google Scholar]
- Domondon, D.; Poppe, J.; Griensven, L. Fruit optimization with wastes used for outdoor cultivation of king Stropharia. Neophilologus 2000, 79, 619–628. [Google Scholar]
- Jin, Z.; Wu, H.; Jiang, J.; Xu, Y. Field experiment on uncooked cultivation of Stropharia rugosoannulata in mountainous areas of southern Zhejiang province. Hans J. Agric. Sci. 2020, 10, 26–34. [Google Scholar]
- Bruhn, J.N.; Abright, N.; Mihail, J.D. Forest farming of wine-cap Stropharia mushrooms. Agrofor. Syst. 2010, 79, 267–275. [Google Scholar] [CrossRef]
- Adamska, I.; Tokarczyk, G.; Spigno, G. Possibilities of using Macrolepiota procera in the production of prohealth food and in medicine. Int. J. Food Sci. 2022, 2022, 5773275. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.W.; Chen, R.C.; Xu, C.; Wang, H.L.; Wang, H.Y.; Ding, Q. Identification, Biological Characteristics and Imitating Wild Cultivation of a Wild Macrolepiota procera Strain. Guizhou Agric. Sci. 2021, 49, 90–91. [Google Scholar]
- Wu, J.; Tokuyama, S.; Nagai, K.; Yasuda, N.; Noguchi, K.; Matsumoto, T.; Hirai, H.; Kawagishi, H. Strophasterols A to D with an unprecedented steroid skeleton: From the mushroom Stropharia rugosoannulata. Angew. Chem. Int. Ed. 2012, 51, 10820–10822. [Google Scholar] [CrossRef]
- Pan, X.J.; Qu, X.H.; Liu, Y.; Wang, S.X. Identification and biological characteristics of wild Macrolepiota procera strain. Jiangsu Agric. Sci. 2016, 44, 199–202. [Google Scholar]
- Pekşen, A.; Kibar, B. Investigations on the cultivation of wild edible mushroom Macrolepiota procera. Uluslararası Tarım Ve Yaban Hayatı Bilim. Derg. 2017, 3, 68–79. [Google Scholar] [CrossRef]
- Pekşen, A.; Kibar, B. Determination of optimum culture conditions for mycelial growth of Macrolepiota procera mushroom. Acta Sci. Pol. Hortorum Cultus 2020, 19, 11–20. [Google Scholar] [CrossRef]
- Jing, B.N.; Chang, X.; Wei, L.; Xie, X.Y.; Zhou, Y.; Wang, Z.Y.; Liu, Y.Q.; Wang, W. Analysis and evaluation of nutrient components, bioactive substances and heavy metal content of Stropharia rugosoannulata in Bo’ai County. Sci. Technol. Food Ind. 2022, 43, 278–285. [Google Scholar]
- Li, X.X.; Zhang, Z.Q.; Wang, L.; Zhao, H.Q.; Jia, Y.H.; Ma, X.; Li, J.Z.; Wang, Y.; Ma, B.J. Three-phase extraction of polysaccharide from Stropharia rugosoannulata: Process optimization, structural characterization and bioactivities. Front. Immunol. 2023, 13, 994706. [Google Scholar] [CrossRef]
- Wu, J.; Suzuki, T.; Choi, J.-H.; Yasuda, N.; Noguchi, K.; Hirai, H.; Kawagishi, H. An unusual sterol from the mushroom Stropharia rugosoannulata. Tetrahedron Lett. 2013, 54, 4900–4902. [Google Scholar] [CrossRef]
- Chen, W.C.; Li, W.; Wu, D.; Zhang, Z.; Chen, H.; Zhang, J.J.; Wang, C.G.; Wu, T.; Yang, Y. Characterization of novel umami-active peptides from Stropharia rugoso-annulata mushroom and in silico study on action mechanism. J. Food Compos. Anal. 2022, 110, 104530. [Google Scholar]
- Palazzolo, E.; Gargano, M.L.; Venturella, G. The nutritional composition of selected wild edible mushrooms from Sicily (southern Italy). Int. J. Food Sci. Nutr. 2012, 63, 79–83. [Google Scholar] [PubMed]
- Lu, Q.; Wang, S.H.; Xue, S.J.; Yang, D.; Li, L. Comparison of non-volatile flavor compounds in Stropharia rugosoannulata soup processed by different methods. J. Food Sci. Technol. 2022, 59, 4025–4036. [Google Scholar] [CrossRef]
- Li, S.R.; Wang, L.; Ni, S.J.; Wang, H.H.; Liu, X.F.; Liu, C.X. The amino acids content of different part of Stropharia rugoso-annulata and their nutrition evaluation. Food Res. Dev. 2017, 38, 95–99. [Google Scholar]
- Wei, J.; Zhao, Q.; Wang, X.; Zhang, G.; Cheng, J.; Chen, Q. Effects of different formulations and cultivation techniques on nutrient quality of Stropharia rugoso-annulata. Vegetables 2022, 8, 51–54. [Google Scholar]
- Aydin, E.; Gurbuz, I.B.; Karahan, H.; Basdar, C. Effect of different processing technologies on chemical properties of wild-grown edible mushroom Macrolepiota procera var. procera (Scop.). J. Food Process. Preserv. 2017, 41, e12802. [Google Scholar] [CrossRef]
- Huang, L.; He, C.M.; Si, C.; Shi, H.Y.; Duan, J. Nutritional, bioactive, and flavor components of giant Stropharia (Stropharia rugoso-annulata): A review. J. Fungi 2023, 9, 792. [Google Scholar] [CrossRef]
- Ćirić, A.; Kruljević, I.; Stojković, D.; Fernandes, Â.; Barros, L.; Calhelha, R.C.; Ferreira, I.C.F.R.; Soković, M.; Glamočlija, J. Comparative investigation on edible mushrooms Macrolepiota mastoidea, M. rhacodes and M. procera: Functional foods with diverse biological activities. Food Funct. 2019, 10, 7678–7686. [Google Scholar] [CrossRef]
- Valverde, M.E.; Hernández-Pérez, T.; Paredes-López, O. Edible Mushrooms: Improving Human Health and Promoting Quality Life. Int. J. Microbiol. 2015, 2015, 376387. [Google Scholar] [CrossRef]
- Niedzielski, P.; Mleczek, M.; Budka, A.; Rzymski, P.; Siwulski, M.; Jasińska, A.; Gąsecka, M.; Budzyńska, S. A screening study of elemental composition in 12 marketable mushroom species accessible in Poland. Eur. Food Res. Technol. 2017, 243, 1759–1771. [Google Scholar] [CrossRef]
- Mleczek, M.; Siwulski, M.; Budka, A.; Niedzielski, P.; Mleczek, P.; Kuczynska-Kippen, N.; Budzynska, S.; Karolewski, Z.; Kalac, P.; Jedryczka, M. Can the concentration of elements in wild-growing mushrooms be deduced from the taxonomic rank? Environ. Res. 2024, 252, 119079. [Google Scholar] [CrossRef] [PubMed]
- Stefanović, V.; Trifković, J.; Djurdjić, S.; Vukojević, V.; Tešić, Ž.; Mutić, J. Study of silver, selenium and arsenic concentration in wild edible mushroom Macrolepiota procera, health benefit and risk. Environ. Sci. Pollut. Res. 2016, 23, 22084–22098. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. Nutrition Components Analyse, Extraction and Antioxidant Properties of Polysaccharide of Stropharia rugosoannulata. Master’s Thesis, Nanjing Normal University, Naijing, China, 2007. [Google Scholar]
- Liu, Y.T.; Sun, J.; Luo, Z.Y.; Rao, S.Q.; Su, Y.J.; Xu, R.R.; Yang, Y.J. Chemical composition of five wild edible mushrooms collected from Southwest China and their antihyperglycemic and antioxidant activity. Food Chem. Toxicol. 2012, 50, 1238–1244. [Google Scholar] [CrossRef]
- Lai, H.H.; Nie, T.T.; Zhang, Y.T.; Chen, Y.; Tao, J.Q.; Lin, T.T.; Ge, T.D.; Li, F.L.; Li, H. Selenium deficiency-induced damage and altered expression of mitochondrial biogenesis markers in the Kidneys of Mice. Biol. Trace Elem. Res. 2020, 199, 185–196. [Google Scholar] [CrossRef]
- Song, Z.; Jia, L.; Xu, F.; Meng, F.; Deng, P.; Fan, K.; Liu, X. Characteristics of Se-Enriched Mycelia by Stropharia rugoso-annulata and its Antioxidant Activities in vivo. Biol. Trace Elem. Res. 2009, 131, 81–89. [Google Scholar] [CrossRef]
- Mazurkiewicz, N.; Podlasińska, J. Bioaccumulation of trace elements in wild-growing edible mushrooms from Lubuskie voivodeship, Poland. Chem. Ecol. 2013, 30, 110–117. [Google Scholar] [CrossRef]
- Fernandes, Â.; Barros, L.; Barreira, J.C.M.; Amilcar, A.L.; Oliveira, M.B.P.P.; Martins, A.; Ferreira, I.C.F.R. Effects of different processing technologies on chemical and antioxidant parameters of Macrolepiota procera wild mushroom. LWT-Food Sci. Technol. 2013, 54, 493–499. [Google Scholar] [CrossRef]
- Ayaz, F.A.; Torun, H.; Özel, A.; Col, M.; Duran, C.; Sesli, E.; Colak, A. Nutritional value of some wild edible mushrooms from Black Sea Region (Turkey). Turk. J. Biochem. 2011, 36, 213–221. [Google Scholar]
- Zhang, B.; Hao, J.; Yin, H.; Duan, C.; Wang, B.; Li, W. Effects of dietary nicotinic acid supplementation on meat quality, carcass characteristics, lipid metabolism, and tibia parameters of Wulong geese. Poult. Sci. 2021, 100, 101430. [Google Scholar] [CrossRef]
- Yan, Q.; Huang, M.; Sun, P.; Cheng, S.; Zhang, Q.; Dai, H. Steroids, fatty acids and ceramide from the mushroom Stropharia rugosoannulata farlow apud murrill. Biochem. Syst. Ecol. 2020, 88, 103963. [Google Scholar] [CrossRef]
- Huang, J.X.; Yuan, S.N.; Pan, J.; Zheng, D.H.; Chen, J.M.; Li, J.; Gui, Q.; Zhou, L.J. Difference of primary nutritional ingredient between Lentinus edodes, Pleurotus ostreatus and Stropharia rugosoannulata grown mainly with rubber wood dust. Chin. J. Trop. Crops 2018, 39, 1625–1629. [Google Scholar]
- Liu, M.M.; Zhang, X.L.; Xu, L.L.; Jiang, P. Analysis of amino acids content and food safety assessment of Stropharia rugosoannulata cultivated in the imitated wild environment under forest. Edible Fungi China 2021, 40, 67–70. [Google Scholar]
- Wei, L.; Jing, B.; Li, X.; Hou, Y.; Xie, X.; Wang, Z.; Liu, Y.; Zhou, Y.; Chang, X.; Wang, W.; et al. Evaluation of nutritional ingredients, biologically active materials, and pharmacological activities of Stropharia rugosoannulata grown under the bamboo forest and in the greenhouse. J. Food Qual. 2021, 2021, 5478227. [Google Scholar] [CrossRef]
- Senatore, F. Chemical constituents of some species of Agaricaceae. Biochem. Syst. Ecol. 1988, 16, 601–604. [Google Scholar]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar]
- Yilmaz, N.; Türkekul, I.; Bulut, S.; Sahin, F.; Bayrak, F.Ö. Fatty acid composition in ten mushroom species collected from Middle Black Sea Region of Turkey. Asian J. Chem. 2013, 25, 1216–1220. [Google Scholar]
- Mleczek, M.; Siwulski, M.; Budka, A.; Mleczek, P.; Budzyńska, S.; Szostek, M.; Kuczyńska-Kippen, N.; Kalač, P.; Niedzielski, P.; Gąsecka, M.; et al. Toxicological risks and nutritional value of wild edible mushroom species -a half-century monitoring study. Chemosphere 2021, 263, 128095. [Google Scholar] [CrossRef]
- Adebola, M.O.; Bello, I.M.; Abioye, J.O.K.; Keturah, B. Myco-chemical and proximate composition of selected mushrooms in lapai niger state nigeria. Int. J. Appl. Sci. 2016, 1, 22–28. [Google Scholar]
- Yan, Q.X. Study on Screening of Active Components and Quality Standard of Stropharia rugosoannulata. Master’s Thesis, Guangxi University of Traditional Chinese Medicine, Nanning, China, 2019. [Google Scholar]
- Zhang, Z.; Kong, F.; Ni, H.; Mo, Z.; Wan, J.B.; Hua, D.; Yan, C. Structural characterization, α-glucosidase inhibitory and DPPH scavenging activities of polysaccharides from guava. Carbohydr. Polym. 2016, 144, 106–114. [Google Scholar]
- Wei, L.; Wang, W.; Xie, X.Y.; Zhou, Y.; Liu, Y.Q.; Ma, Y.N.; Ning, E.J.; Wang, T.; Li, N.J.; Jing, B.N. Optimization of extraction process of polysaccharides from Stropharia rugosoannulata in Bo’ai county by response surface method and evalation of their antibacterial and antioxidant activity. Sci. Technol. Food Ind. 2023, 44, 213–220. [Google Scholar]
- Jiang, L.; Hou, Y.; Ding, X. Structure identification and biological activities of a new polysaccharides from Stropharia rugosoannulata. Lat. Am. J. Pharm. 2020, 39, 1594–1604. [Google Scholar]
- Liu, Y.; Hu, C.F.; Feng, X.; Cheng, L.; Ibrahim, S.A.; Wang, C.T.; Huang, W. Isolation, characterization and antioxidant of polysaccharides from Stropharia rugosoannulata. Int. J. Biol. Macromol. 2020, 155, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Xue, S.J.; Yang, D.; Wang, S.H.; Li, L. Effects of three extraction methods on antioxidant properties of crude polysaccharides from Stropharia rugoso. Food Sci. Technol. 2021, 46, 171–178. [Google Scholar]
- Wang, Q.; Zhao, Y.; Feng, X.; Ibrahim, S.A.; Huang, W.; Liu, Y. Effects of drying on the structural characteristics and antioxidant activities of polysaccharides from Stropharia rugosoannulata. J. Food Sci. Technol. 2021, 58, 3622–3631. [Google Scholar] [CrossRef]
- Jiang, L. The Research on Preparation, Structural Identification and Biological Activity of Polysaccharide from Stropharia rugosoannulata (SR-1) and Polysaccharide from Tricholomalascivum (Fr.) Gillet (TLG-1). Master’s Thesis, China West Normal University, Nanchong, China, 2019. [Google Scholar]
- Kremmyda, A.; MacNaughtan, W.; Arapoglou, D.; Eliopoulos, C.; Metafa, M.; Harding, S.E.; Israilides, C. The detection, purity and structural properties of partially soluble mushroom and cereal β-D-glucans: A solid-state NMR study. Carbohydr. Polym. 2021, 266, 118103. [Google Scholar]
- Georgiev, Y.N.; Vasicek, O.; Dzhambazov, B.; Batsalova, T.G.; Denev, P.N.; Dobreva, L.I.; Danova, S.T.; Simova, S.D.; Wold, C.W.; Ognyanov, M.H.; et al. Structural features and immunomodulatory effects of water-extractable polysaccharides from Macrolepiota procera (Scop.) Singer. J. Fungi 2022, 8, 848. [Google Scholar] [CrossRef]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef]
- Gil-Ramírez, A.; Pavo-Caballero, C.; Baeza, E.; Baenas, N.; Garcia-Viguera, C.; Marín, F.R.; Soler-Rivas, C. Mushrooms do not contain flavonoids. J. Funct. Foods 2016, 25, 1–13. [Google Scholar] [CrossRef]
- Erbiai, E.H.; da Silva, L.P.; Saidi, R.; Lamrani, Z.; Esteves da Silva, J.C.G.; Maouni, A. Chemical composition, bioactive compounds, and antioxidant activity of two wild edible mushrooms Armillaria mellea and Macrolepiota procera from two countries (Morocco and Portugal). Biomolecules 2021, 11, 575. [Google Scholar] [CrossRef]
- Maria-Lidia, P.; Teodora, C.; Ioana, N.; Elena, D.L.; Elena, G.C. Polyphenols contents and antioxidant activity of some romanian wild edible mushrooms. Farmacia 2016, 64, 231–236. [Google Scholar]
- Cateni, F.; Gargano, M.L.; Procida, G.; Venturella, G.; Cirlincione, F.; Ferraro, V. Mycochemicals in wild and cultivated mushrooms: Nutrition and health. Phytochem. Rev. 2021, 21, 339–383. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide—Natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Liu, X.M.; Saburi, T.; Kubota, S.; Huang, P.X.; Wada, Y.J. Thermal stability and oxidation characteristics of α-pinene, β-pinene and α-pinene/β-pinene mixture. RSC Adv. 2021, 11, 20529–20540. [Google Scholar] [CrossRef]
- Nguyen, N.-H.; Ha, T.K.Q.; Yang, J.L.; Pham, H.T.T.; Oh, W.K. Triterpenoids from the genus Gynostemma: Chemistry and pharmacological activities. J. Ethnopharmacol. 2021, 268, 113574. [Google Scholar] [CrossRef]
- Silva, G.d.S.; Marques, J.N.d.J.; Linhares, E.P.M.; Bonora, C.M.; Costa, É.T.; Saraiva, M.F. Review of anticancer activity of monoterpenoids: Geraniol, nerol, geranial and neral. Chem.-Biol. Interact. 2022, 362, 109994. [Google Scholar]
- Jeyakumar, G.E.; Lawrence, R. Mechanisms of bactericidal action of eugenol against Escherichia coli. J. Herb. Med. 2021, 26, 100406. [Google Scholar] [CrossRef]
- Michihiro, S.; Hidekazu, M.; Andikuo, I. A comparison of hypocholesterolemic activity of β-Sitosterol and β-Sitostanol in Rats. J. Nutr. 1977, 107, 2011–2019. [Google Scholar]
- Zheng, M.; Lai, H.; Andrew-Lin, K. Valorization of vanillyl alcohol by pigments: Prussian blue analogue as a highly-effective heterogeneous catalyst for aerobic oxidation of vanillyl alcohol to vanillin. Waste Biomass Valorization 2018, 10, 2933–2942. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.; Ma, H.; Wu, D.; Zhang, Z.; Yang, Y. Structural characterization and angiotensin-converting enzyme (ace) inhibitory mechanism of Stropharia rugosoannulata mushroom peptides prepared by ultrasound. Ultrason. Sonochem 2022, 88, 106074. [Google Scholar]
- Li, W.; Chen, W.C.; Wu, D.; Zhang, Z.; Yang, Y. Taste peptides derived from Stropharia rugosoannulata fermentation mycelium and molecular docking to the taste receptor T1R1/T1R3. Front. Nutr. 2022, 9, 960218. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.B.; Zhang, J.J.; Wu, S.D.; Bai, J.; Zhuo, X.Y.; Zhang, J.X.; Kuai, B.K.; Chen, H. Correction: Transcriptomic analysis of Stropharia rugosoannulata reveals carbohydrate metabolism and cold resistance mechanisms under low-temperature stress. AMB Express 2022, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Akata, I.; Zengin, G.; Picot, C.M.N.; Mahomoodally, M.F. Enzyme inhibitory and antioxidant properties of six mushroom species from the Agaricaceae family. S. Afr. J. Bot. 2019, 120, 95–99. [Google Scholar] [CrossRef]
- Li, W.; Feng, J.; Ma, H.L.; Chen, W.C.; Wu, D.; Zhang, Z.; Yang, Y. Analysis of the characteristic flavor components and flavor characteristics of the fermentation of Stropharia rugoso-annulata based on targeted metabolite assay. J. Food Saf. Qual. 2022, 13, 2736–2744. [Google Scholar]
- Kosanić, M.; Ranković, B.; Rančić, A.; Stanojković, T. Evaluation of metal concentration and antioxidant, antimicrobial, and anticancer potentials of two edible mushrooms Lactarius deliciosus and Macrolepiota procera. J. Food Drug Anal. 2016, 24, 477–484. [Google Scholar] [CrossRef]
- Gao, Y.L.; Abuduaini, G.; Yang, C.H.; Zhang, S.S.; Zhang, Y.R.; Fan, H.X.; Teng, X.; Bao, C.; Liu, H.C.; Wang, D.W.; et al. Isolation, purification, and structural elucidation of Stropharia rugosoannulata polysaccharides with hypolipidemic effect. Front. Nutr. 2022, 9, 1092582. [Google Scholar] [CrossRef]
- Wei, L.; Wang, W.; Hou, Y.; Wang, X.; Liu, Y.; Xie, X.; Li, X.; Wang, T.; Jing, B.; Kumar, Y. Assessment of the quality characteristics of Stropharia rugosoannulata subjected to five different drying methods. J. Food Process. Preserv. 2023, 2023, 2518620. [Google Scholar] [CrossRef]
- Li, S.W.; Zhao, S.X.; Hu, C.H.; Mao, C.Z.; Guo, L.Z.; Yu, H.L.; Yu, H. Whole genome sequence of an edible mushroom Stropharia rugosoannulata (Daqiugaigu). J. Fungi 2022, 8, 99. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.C.; Zhang, Z.; Wu, D.; Liu, P.; Li, Z.P.; Yang, Y. Combined peptidomics and metabolomics analyses to characterize the digestion properties and activity of Stropharia rugosoannulata protein–peptide-based materials. Foods 2024, 13, 2546. [Google Scholar] [CrossRef]
- Wu, X.; Du, Z.; Liu, L.; Chen, Z.; Li, Y.; Fu, S. Integrative analysis of transcriptome and metabolome sheds light on flavonoid biosynthesis in the fruiting body of Stropharia rugosoannulata. J. Fungi 2024, 10, 254. [Google Scholar] [CrossRef]
- Yang, Y.; Meng, G.; Ni, S.; Zhang, H.; Dong, C. Genomic Analysis of Stropharia rugosoannulata Reveals Its Nutritional Strategy and Application Potential in Bioremediation. J. Fungi 2022, 8, 162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tian, G.; Geng, X.; Zhao, Y.; Ng, T.; Zhao, L.; Wang, H. Isolation and characterization of a novel lectin from the edible mushroom Stropharia rugosoannulata. Molecules 2014, 19, 19880–19891. [Google Scholar] [CrossRef] [PubMed]
- El-Enshasy, H.A.; Hatti-Kaul, R. Mushroom immunomodulators: Unique molecules with unlimited applications. Trends Biotechnol. 2013, 31, 668–677. [Google Scholar] [CrossRef]
- Wang, H.; Chen, H.; Zhang, J.; Chen, M. Research progresses on bioactive components in Stropharia rugosoannulata and their pharmacological effects. Acta Edulis Fungi 2018, 25, 115–120. [Google Scholar]
- Seçme, M.; Kaygusuz, O.; Eroglu, C.; Dodurga, Y.; Colak, O.F.; Atmaca, P. Potential anticancer activity of the parasol mushroom, Macrolepiota procera (Agaricomycetes), against the A549 human lung cancer cell line. Int. J. Med. Mushrooms 2018, 11, 1075–1086. [Google Scholar]
- Zara, R.; Rasul, A.; Sultana, T.; Jabeen, F.; Selamoglu, Z. Identification of Macrolepiota procera extract as a novel G6PD inhibitor for the treatment of lung cancer. Saudi J. Biol. Sci. 2022, 29, 3372–3379. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, J.; Lang, Y.; Yao, L.; Xu, H.; Shi, H.; Xu, S. A polysaccharide from Armillaria mellea exhibits strong in vitro anticancer activity via apoptosis-involved mechanisms. Int. J. Biol. Macromol. 2012, 51, 663–667. [Google Scholar] [CrossRef]
- Nowacka, N.; Nowak, R.; Drozd, M.; Olech, M.; Los, R.; Malm, A. Analysis of phenolic constituents, antiradical and antimicrobial activity of edible mushrooms growing wild in Poland. LWT Food Sci. Technol. 2014, 59, 689–694. [Google Scholar] [CrossRef]
- Elsayed, E.A.; El-Enshasy, H.; Wadaan, M.A.M.; Aziz, R. Mushrooms: A potential natural source of anti-inflammatory compounds for medical applications. Mediat. Inflamm. 2014, 2014, 805841. [Google Scholar] [CrossRef]
- Zhai, X.; Zhao, A.; Geng, L.; Xu, C. Fermentation characteristics and hypoglycemic activity of an exopolysaccharide produced by submerged culture of Stropharia rugosoannulata #2. Ann. Microbiol. 2012, 63, 1013–1020. [Google Scholar] [CrossRef]
- Falandysz, J.; Kunito, T.; Kubota, R.; Gucia, M.; Mazur, A.; Falandysz, J.J.; Tanabe, S. Some mineral constituents of parasol mushroom (Macrolepiota procera). J. Environ. Sci. Health Part B 2008, 43, 187–192. [Google Scholar] [CrossRef]
- Kojta, A.; Gucia, M.; Krasińska, G.; Saba, M.; Nnorom, I.C.; Falandysz, J. Mineral constituents of edible field parasol (Macrolepiota procera) mushrooms and the underlying substrate from upland regions of Poland: Bioconcentration potential, intake benefits, and toxicological risk. Pol. J. Environ. Stud. 2016, 25, 2445–2460. [Google Scholar] [CrossRef]
- Bucurica, I.A.; Dulama, I.D.; Radulescu, C.; Banica, A.L.; Stanescu, S.G. Heavy metals and associated risks of wild edible mushrooms consumption: Transfer factor, carcinogenic risk, and health risk index. J. Fungi 2024, 10, 844. [Google Scholar] [CrossRef] [PubMed]
- Golak−Siwulska, I.; Kałużewicz, A.; Spiżewski, T.; Sobieralski, K. Zawartość związków biologicznie aktywnych w grzybach jadalnych dziko rosnących. Sylwan 2018, 162, 238–247. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Components | S. rugosoannulata | M. procera | Refs |
|---|---|---|---|
| Moisture | 90–93% | 88–92% | [3] |
| Proteins | 17.99–25.67% | 7.62–39.68% | [26,27] |
| Carbohydrates | 45.17–54.60% | 40.90–60.30% | [28,29,30] |
| Lipids | 1.33–2.30% (over 77% unsaturated fatty acids, including 57% linoleic acid and 13% palmitic acid) | 0.70–4.23% (rich in unsaturated fatty acids, predominantly linoleic acid) | [29,31] |
| Macroelements | K (1600 mg/100 g), P (75–100 mg/100 g), Mg (20–30 mg/100 g), Ca (70–80 mg/100 g) | K (300–500 mg/100 g), Mg (15–25 mg/100 g), Ca (400–500 mg/100 g) | [32,33,34] |
| Trace elements | Fe (19.5–24.5 mg/100 g), Zn (5.5–10.0 mg/100 g), Se (0.1–0.5 mg/100 g) | Fe (1.5–3 mg/100 g) Zn (0.5–1.2 mg/100 g) | [35,36,37,38,39] |
| Vitamins | B vitamins (B2, B3, B5, B6, B9, and B12), Vitamin C, E, K, Provitamin D2, Beta-carotene | B vitamins (B2, B3, and B5), Vitamin D, E, K, Beta-carotene | [20,40,41,42,43] |
| Essential amino acids | 6.43–11.70%, with isoleucine being the most abundant. | Relatively low | [26,44,45,46] |
| Nonessential amino acids | 2.88–6.84% glutamic acid and 1.72–3.07% aspartic acid | Alanine (1.10 g/100 g) | [27,45,47] |
| Method | Characteristics | Extracted Compounds | Refs |
|---|---|---|---|
| Solvent extraction | Simple operation and high selectivity, but solvent residue and long extraction time are limitations. Optimization of solvent polarity and concentration can improve performance. | Polysaccharides, Gal (62.3% w/w); phenolics, 60–90%, protocatechuic acid 2.23–2.25 µg/g dw, cinnamic acid 8.67 µg/g dw | [79] |
| Hot water extraction | Eco-friendly and non-toxic, but with low efficiency and prolonged duration. Extraction temperature and time can be optimized to enhance outcomes. | Polysaccharides, 95.43% total sugar, mannose:glucose:galactose:methylgalactose = 8:12:58:12 | [57,80] |
| Ultrasound-assisted extraction (UAE) | Increasing extraction efficiency and purity but requiring specialized equipment and showing energy-intensive. Frequency and extraction time can be optimized for better results. | Polysaccharides, 13.25–22.37% | [81] |
| Microwave- assisted extraction (MAE) | Fast and efficient, suitable for thermosensitive compounds. However, uneven heating may impact the extraction process. Extraction power and time can be optimized to mitigate this issue. | Polyphenols, 1.22–6.80%; amino acids, 3.57–23.06%, alanine 0.41%, threonine 1.26%, tryptophan 2.82% | [64,79] |
| Ion exchange chromatography | High selectivity and effective for biomacromolecule separation with strong operability. However, it demands strict sample conditions, is unsuitable for non-ionic compounds, and has a slow separation rate. | Polysaccharides, 90.34–91.23% total sugar, (1→, 6)-α-D-dextran cytoskeleton | [56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Yang, X.; Zhu, J. Nutritional and Therapeutic Potential of Stropharia rugosoannulata and Macrolepiota procera: From Composition to Health-Promoting Effect. J. Fungi 2025, 11, 259. https://doi.org/10.3390/jof11040259
Wang Q, Yang X, Zhu J. Nutritional and Therapeutic Potential of Stropharia rugosoannulata and Macrolepiota procera: From Composition to Health-Promoting Effect. Journal of Fungi. 2025; 11(4):259. https://doi.org/10.3390/jof11040259
Chicago/Turabian StyleWang, Qian, Xiaoyan Yang, and Jiangxiong Zhu. 2025. "Nutritional and Therapeutic Potential of Stropharia rugosoannulata and Macrolepiota procera: From Composition to Health-Promoting Effect" Journal of Fungi 11, no. 4: 259. https://doi.org/10.3390/jof11040259
APA StyleWang, Q., Yang, X., & Zhu, J. (2025). Nutritional and Therapeutic Potential of Stropharia rugosoannulata and Macrolepiota procera: From Composition to Health-Promoting Effect. Journal of Fungi, 11(4), 259. https://doi.org/10.3390/jof11040259