

Poly ADP-Ribosylation in a Plant Pathogenic Oomycete Phytophthora infestans: A Key Controller of Growth and Host Plant Colonisation
 , ,
, ,  , and
, and {kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. DNA Extraction, Purification and Sequencing
2.2. Production and Purification of dsRNA
2.3. Treatment of Mycelium with dsRNA and PARP Inhibitor 3-Aminobenzamide (3AB)
2.4. Real-Time Quantitative RT-PCR (RT-qPCR)
2.5. Immunological Detection of Poly ADP-Ribose (PAR)
2.6. Test on Detached Leaves
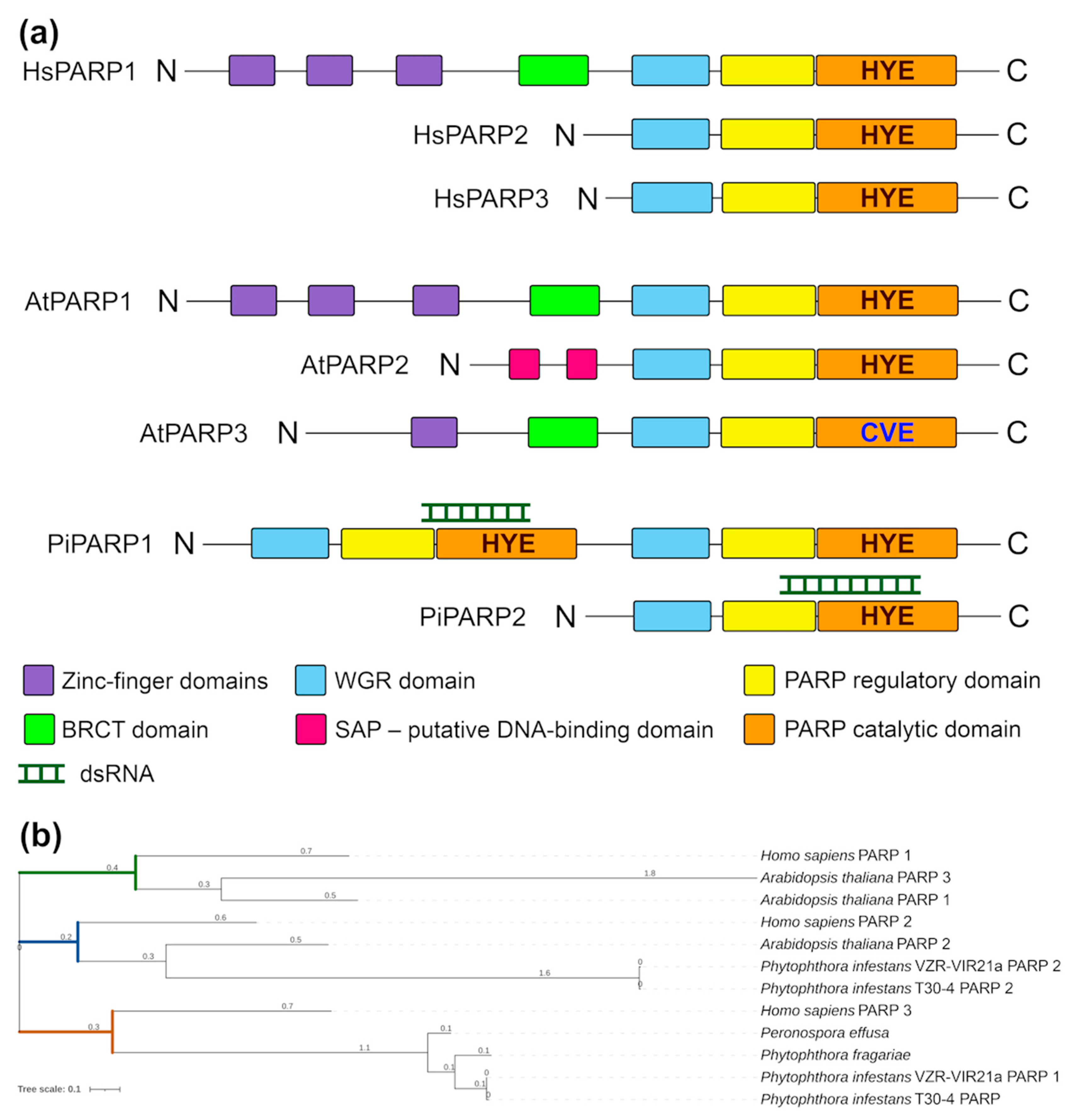
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Angmo, D.; Sharma, S.P.; Kalia, A. Breeding Strategies for Late Blight Resistance in Potato Crop: Recent Developments. Mol. Biol. Rep. 2023, 50, 7879–7891. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Jiao, F. Effectors of Phytophthora Pathogens Are Powerful Weapons for Manipulating Host Immunity. Planta 2019, 250, 413–425. [Google Scholar] [CrossRef]
- Citarelli, M.; Teotia, S.; Lamb, R.S. Evolutionary History of the Poly(ADP-Ribose) Polymerase Gene Family in Eukaryotes. BMC Evol. Biol. 2010, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Perina, D.; Mikoč, A.; Ahel, J.; Ćetković, H.; Žaja, R.; Ahel, I. Distribution of Protein Poly(ADP-Ribosyl)Ation Systems across All Domains of Life. DNA Repair. 2014, 23, 4–16. [Google Scholar] [CrossRef]
- Rissel, D.; Peiter, E. Poly(ADP-Ribose) Polymerases in Plants and Their Human Counterparts: Parallels and Peculiarities. Int. J. Mol. Sci. 2019, 20, 1638. [Google Scholar] [CrossRef]
- Dasovich, M.; Leung, A.K.L. PARPs and ADP-Ribosylation: Deciphering the Complexity with Molecular Tools. Mol. Cell 2023, 83, 1552–1572. [Google Scholar] [CrossRef] [PubMed]
- Spechenkova, N.; Samarskaya, V.O.; Kalinina, N.O.; Zavriev, S.K.; MacFarlane, S.; Love, A.J.; Taliansky, M. Plant Poly(ADP-Ribose) Polymerase 1 Is a Potential Mediator of Cross-Talk between the Cajal Body Protein Coilin and Salicylic Acid-Mediated Antiviral Defence. Viruses 2023, 15, 1282. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, N.O.; Spechenkova, N.; Ilina, I.; Samarskaya, V.O.; Bagdasarova, P.; Zavriev, S.K.; Love, A.J.; Taliansky, M. Disruption of Poly(ADP-Ribosyl)Ation Improves Plant Tolerance to Methyl Viologen-Mediated Oxidative Stress via Induction of ROS Scavenging Enzymes. Int. J. Mol. Sci. 2024, 25, 9367. [Google Scholar] [CrossRef] [PubMed]
- Jubin, T.; Kadam, A.; Jariwala, M.; Bhatt, S.; Sutariya, S.; Gani, A.R.; Gautam, S.; Begum, R. The PARP Family: Insights into Functional Aspects of Poly (ADP-ribose) Polymerase-1 in Cell Growth and Survival. Cell Prolif. 2016, 49, 421. [Google Scholar] [CrossRef]
- Carreras-Puigvert, J.; Zitnik, M.; Jemth, A.-S.; Carter, M.; Unterlass, J.E.; Hallström, B.; Loseva, O.; Karem, Z.; Calderón-Montaño, J.M.; Lindskog, C.; et al. A Comprehensive Structural, Biochemical and Biological Profiling of the Human NUDIX Hydrolase Family. Nat. Commun. 2017, 8, 1541. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Chesarone-Cataldo, M.; Todorova, T.; Huang, Y.-H.; Chang, P. A Systematic Analysis of the PARP Protein Family Identifies New Functions Critical for Cell Physiology. Nat. Commun. 2013, 4, 2240. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Zheng, J.-Y.; Huang, L.-L.; Wang, Y.-H.; Yao, D.-F.; Dai, H.-B. Human PARP1 Substrates and Regulators of Its Catalytic Activity: An Updated Overview. Front. Pharmacol. 2023, 14, 1137151. [Google Scholar] [CrossRef]
- Langelier, M.-F.; Eisemann, T.; Riccio, A.A.; Pascal, J.M. PARP Family Enzymes: Regulation and Catalysis of the Poly(ADP-Ribose) Posttranslational Modification. Curr. Opin. Struct. Biol. 2018, 53, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Vainonen, J.P.; Shapiguzov, A.; Vaattovaara, A.; Kangasjärvi, J. Plant PARPs, PARGs and PARP-like Proteins. Curr. Protein Pept. Sci. 2016, 17, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Pan, W.; Chen, W.; Lian, Q.; Wu, Q.; Lv, Z.; Cheng, X.; Ge, X. New Perspectives on the Plant PARP Family: Arabidopsis PARP3 Is Inactive, and PARP1 Exhibits Predominant Poly (ADP-Ribose) Polymerase Activity in Response to DNA Damage. BMC Plant Biol. 2019, 19, 364. [Google Scholar] [CrossRef] [PubMed]
- Maluchenko, N.V.; Koshkina, D.O.; Feofanov, A.V.; Studitsky, V.M.; Kirpichnikov, M.P. Poly(ADP-Ribosyl) Code Functions. Acta Nat. 2021, 13, 58–69. [Google Scholar] [CrossRef]
- Spechenkova, N.; Kalinina, N.O.; Zavriev, S.K.; Love, A.J.; Taliansky, M. ADP-Ribosylation and Antiviral Resistance in Plants. Viruses 2023, 15, 241. [Google Scholar] [CrossRef] [PubMed]
- Jubin, T.; Kadam, A.; Saran, S.; Begum, R. Poly (ADP-Ribose) Polymerase1 Regulates Growth and Multicellularity in D. Discoideum. Differentiation 2016, 92, 10–23. [Google Scholar] [CrossRef]
- Semighini, C.P.; Savoldi, M.; Goldman, G.H.; Harris, S.D. Functional Characterization of the Putative Aspergillus Nidulans Poly(ADP-Ribose) Polymerase Homolog PrpA. Genetics 2006, 173, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Kothe, G.O.; Kitamura, M.; Masutani, M.; Selker, E.U.; Inoue, H. PARP Is Involved in Replicative Aging in Neurospora crassa. Fungal Genet. Biol. 2010, 47, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, Y.; Xiong, X.; Yan, Y.; Lou, J.; Noman, M.; Li, D.; Song, F. The Ser/Thr Protein Kinase FonKin4-Poly(ADP-Ribose) Polymerase FonPARP1 Phosphorylation Cascade Is Required for the Pathogenicity of Watermelon Fusarium Wilt Fungus Fusarium Oxysporum f. Sp. Niveum. Front. Microbiol. 2024, 15, 1397688. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Gao, G.; Zheng, W.; Liu, H.; Pan, W.; Xia, X.; Zhang, D.; Lin, W.; Wang, Z.; Feng, B. PARylation of 14-3-3 Proteins Controls the Virulence of Magnaporthe Oryzae. Nat. Commun. 2024, 15, 8047. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid Isolation of High Molecular Weight Plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Horn, T.; Boutros, M. E-RNAi: A Web Application for the Multi-Species Design of RNAi Reagents—2010 Update. Nucleic Acids Res. 2010, 38, W332–W339. [Google Scholar] [CrossRef] [PubMed]
- Samarskaya, V.O.; Spechenkova, N.; Markin, N.; Suprunova, T.P.; Zavriev, S.K.; Love, A.J.; Kalinina, N.O.; Taliansky, M. Impact of Exogenous Application of Potato Virus Y-Specific dsRNA on RNA Interference, Pattern-Triggered Immunity and Poly(ADP-Ribose) Metabolism. Int. J. Mol. Sci. 2022, 23, 7915. [Google Scholar] [CrossRef]
- Samarskaya, V.O.; Spechenkova, N.; Ilina, I.; Suprunova, T.P.; Kalinina, N.O.; Love, A.J.; Taliansky, M.E. A Non-Canonical Pathway Induced by Externally Applied Virus-Specific dsRNA in Potato Plants. Int. J. Mol. Sci. 2023, 24, 15769. [Google Scholar] [CrossRef]
- Ivanov, A.A.; Tyapkin, A.V.; Golubeva, T.S. How Does the Sample Preparation of Phytophthora Infestans Mycelium Affect the Quality of Isolated RNA? Curr. Issues Mol. Biol. 2023, 45, 3517–3524. [Google Scholar] [CrossRef]
- Méchin, V.; Damerval, C.; Zivy, M. Total Protein Extraction with TCA-Acetone. In Plant Proteomics: Methods and Protocols; Thiellement, H., Zivy, M., Damerval, C., Méchin, V., Eds.; Humana Press: Totowa, NJ, USA, 2007; pp. 1–8. ISBN 978-1-59745-227-4. [Google Scholar]
- Abramoff, M.D.; Magelhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Internat. 2004, 11, 36–42. [Google Scholar]
- Sundaresha, S.; Bairwa, A.; Tomar, M.; Kumar, R.; Venkatasalam, E.P.; Sagar, V.; Bhardwaj, V.; Sharma, S. In Vitro Method for Synthesis of Large-Scale dsRNA Molecule as a Novel Plant Protection Strategy. In Plant Gene Silencing: Methods and Protocols; Mysore, K.S., Senthil-Kumar, M., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2022; pp. 211–226. ISBN 978-1-07-161875-2. [Google Scholar]
- Chaudhary, D.; Jeena, A.S.; Rohit; Gaur, S.; Raj, R.; Mishra, S.; Kajal; Gupta, O.P.; Meena, M.R. Advances in RNA Interference for Plant Functional Genomics: Unveiling Traits, Mechanisms, and Future Directions. Appl. Biochem. Biotechnol. 2024, 196, 5681–5710. [Google Scholar] [CrossRef]
- Adams-Phillips, L.; Briggs, A.G.; Bent, A.F. Disruption of Poly(ADP-Ribosyl)Ation Mechanisms Alters Responses of Arabidopsis to Biotic Stress. Plant Physiol. 2010, 152, 267–280. [Google Scholar] [CrossRef]
- Rissel, D.; Heym, P.P.; Thor, K.; Brandt, W.; Wessjohann, L.A.; Peiter, E. No Silver Bullet-Canonical Poly(ADP-Ribose) Polymerases (PARPs) Are No Universal Factors of Abiotic and Biotic Stress Resistance of Arabidopsis Thaliana. Front. Plant Sci. 2017, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Keppler, B.D.; Song, J.; Nyman, J.; Voigt, C.A.; Bent, A.F. 3-Aminobenzamide Blocks MAMP-Induced Callose Deposition Independently of Its Poly(ADPribosyl)Ation Inhibiting Activity. Front. Plant Sci. 2018, 9, 1907. [Google Scholar] [CrossRef]
- Adams-Phillips, L.; Wan, J.; Tan, X.; Dunning, F.M.; Meyers, B.C.; Michelmore, R.W.; Bent, A.F. Discovery of ADP-Ribosylation and Other Plant Defense Pathway Elements Through Expression Profiling of Four Different Arabidopsis–Pseudomonas R-Avr Interactions. MPMI 2008, 21, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Briggs, A.G.; Adams-Phillips, L.C.; Keppler, B.D.; Zebell, S.G.; Arend, K.C.; Apfelbaum, A.A.; Smith, J.A.; Bent, A.F. A Transcriptomics Approach Uncovers Novel Roles for Poly(ADP-Ribosyl)Ation in the Basal Defense Response in Arabidopsis Thaliana. PLoS ONE 2017, 12, e0190268. [Google Scholar] [CrossRef] [PubMed]
- Krishnan Muthaiah, V.P.; Kaliyappan, K.; Mahajan, S.D. Poly ADP-Ribose Polymerase-1 Inhibition by 3-Aminobenzamide Recuperates HEI-OC1 Auditory Hair Cells from Blast Overpressure-Induced Cell Death. Front. Cell Dev. Biol. 2023, 11, 1047308. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samarskaya, V.O.; Koblova, S.; Suprunova, T.; Rogozhin, E.A.; Spechenkova, N.; Yakunina, S.; Love, A.J.; Kalinina, N.O.; Taliansky, M. Poly ADP-Ribosylation in a Plant Pathogenic Oomycete Phytophthora infestans: A Key Controller of Growth and Host Plant Colonisation. J. Fungi 2025, 11, 29. https://doi.org/10.3390/jof11010029
Samarskaya VO, Koblova S, Suprunova T, Rogozhin EA, Spechenkova N, Yakunina S, Love AJ, Kalinina NO, Taliansky M. Poly ADP-Ribosylation in a Plant Pathogenic Oomycete Phytophthora infestans: A Key Controller of Growth and Host Plant Colonisation. Journal of Fungi. 2025; 11(1):29. https://doi.org/10.3390/jof11010029
Chicago/Turabian StyleSamarskaya, Viktoriya O., Sofya Koblova, Tatiana Suprunova, Eugene A. Rogozhin, Nadezhda Spechenkova, Sofiya Yakunina, Andrew J. Love, Natalia O. Kalinina, and Michael Taliansky. 2025. "Poly ADP-Ribosylation in a Plant Pathogenic Oomycete Phytophthora infestans: A Key Controller of Growth and Host Plant Colonisation" Journal of Fungi 11, no. 1: 29. https://doi.org/10.3390/jof11010029
APA StyleSamarskaya, V. O., Koblova, S., Suprunova, T., Rogozhin, E. A., Spechenkova, N., Yakunina, S., Love, A. J., Kalinina, N. O., & Taliansky, M. (2025). Poly ADP-Ribosylation in a Plant Pathogenic Oomycete Phytophthora infestans: A Key Controller of Growth and Host Plant Colonisation. Journal of Fungi, 11(1), 29. https://doi.org/10.3390/jof11010029