
Redundant and Distinct Roles of Two 14-3-3 Proteins in Fusarium sacchari, Pathogen of Sugarcane Pokkah Boeng Disease



Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culturing Conditions
2.2. Generation of Mutant Strains
2.3. RNA Extraction and Quantitative Real-Time RT-PCR
2.4. Induction of Macrospores
2.5. Microscopy
2.6. Pathogenicity Assays
2.7. Transcriptomic Analysis
2.8. Yeast Two-Hybrid Assays
2.9. Statistical Analysis
3. Results
3.1. Conservation of 14-3-3 Proteins in Fusarium Species
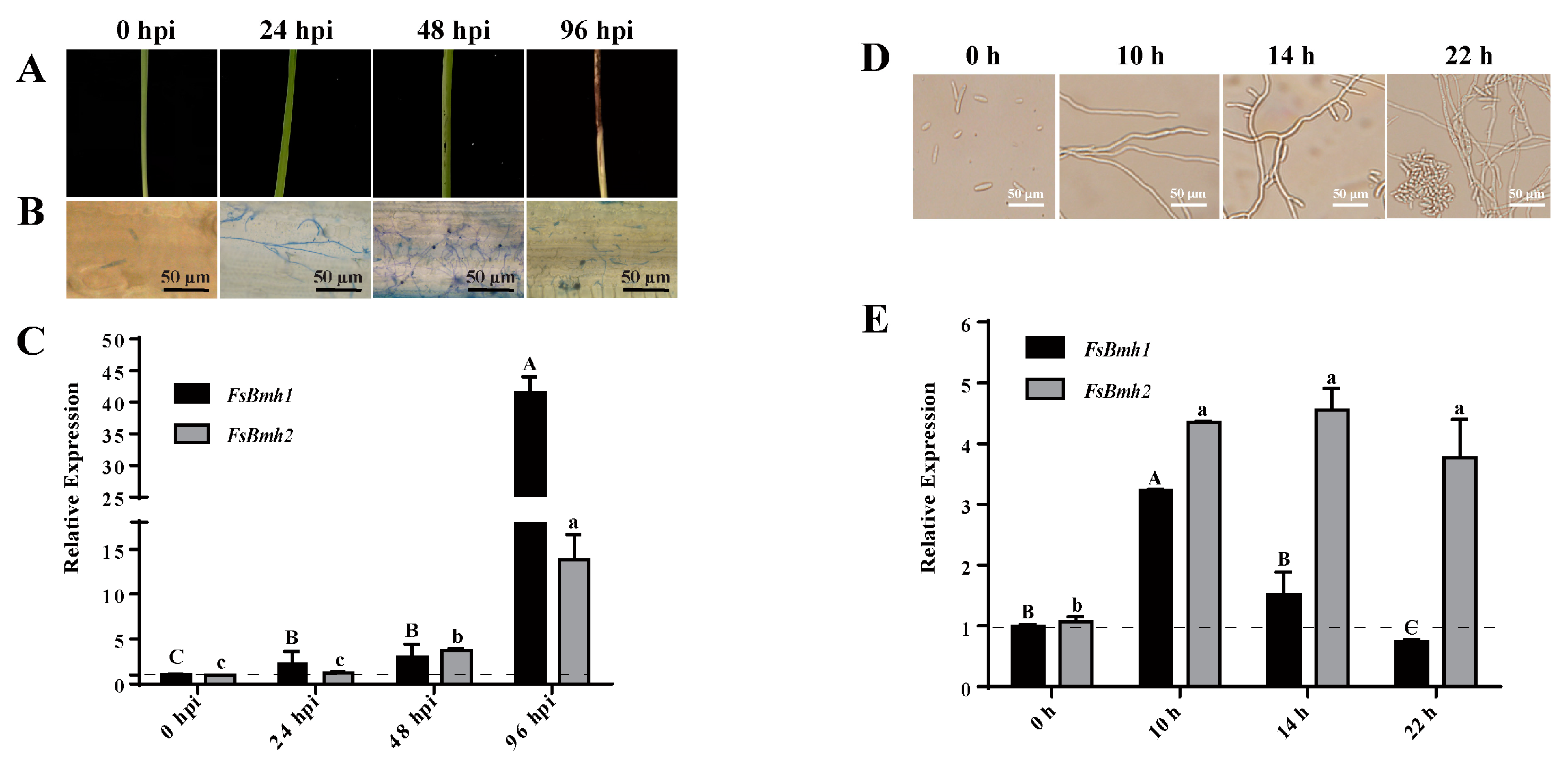
3.2. Expression Patterns of FsBmh1 and FsBmh2 during Infection and Vegetative Growth
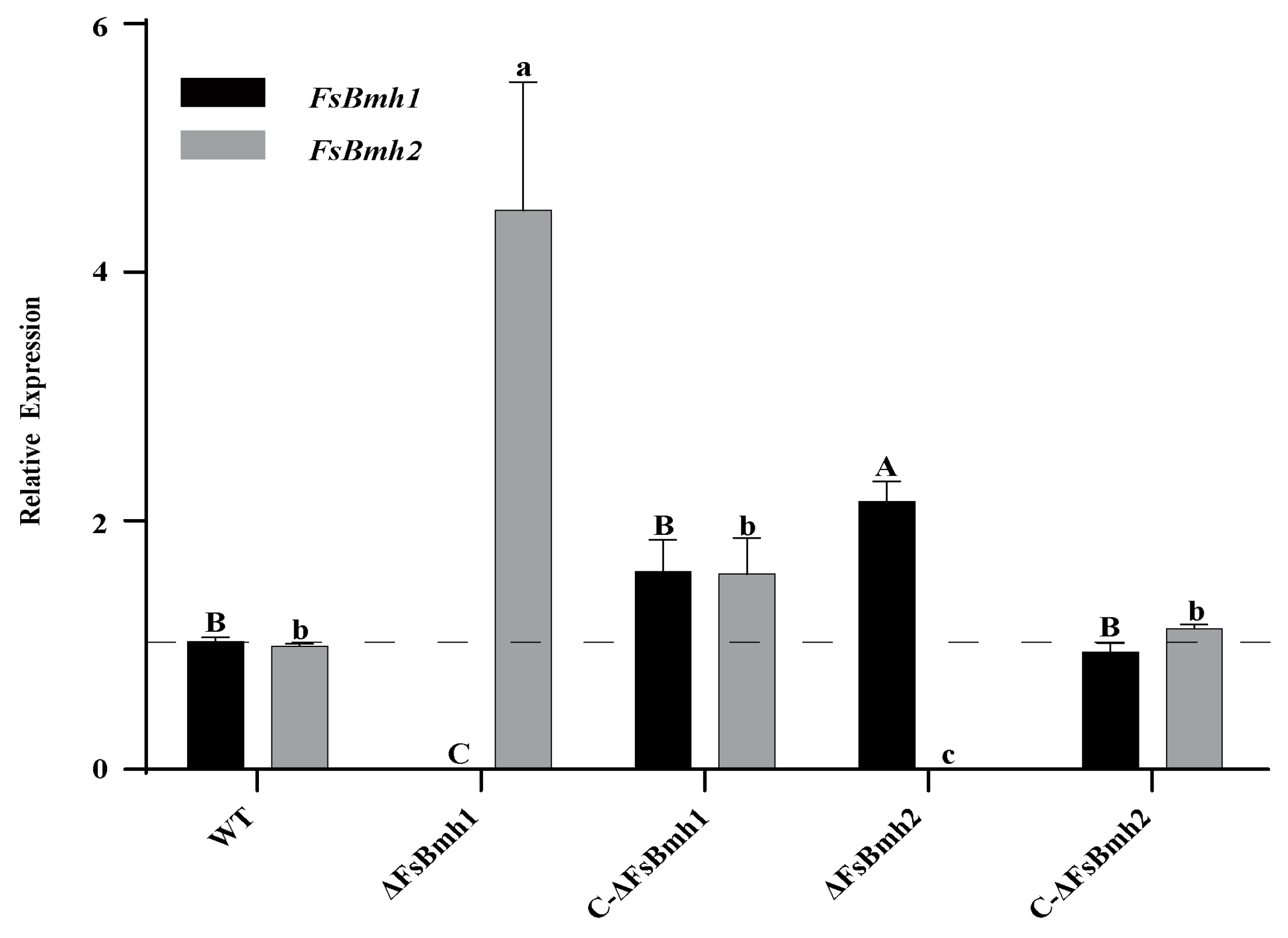
3.3. The Expression of FsBmh1 and FsBmh2 Are Mutually Compensated in Cells
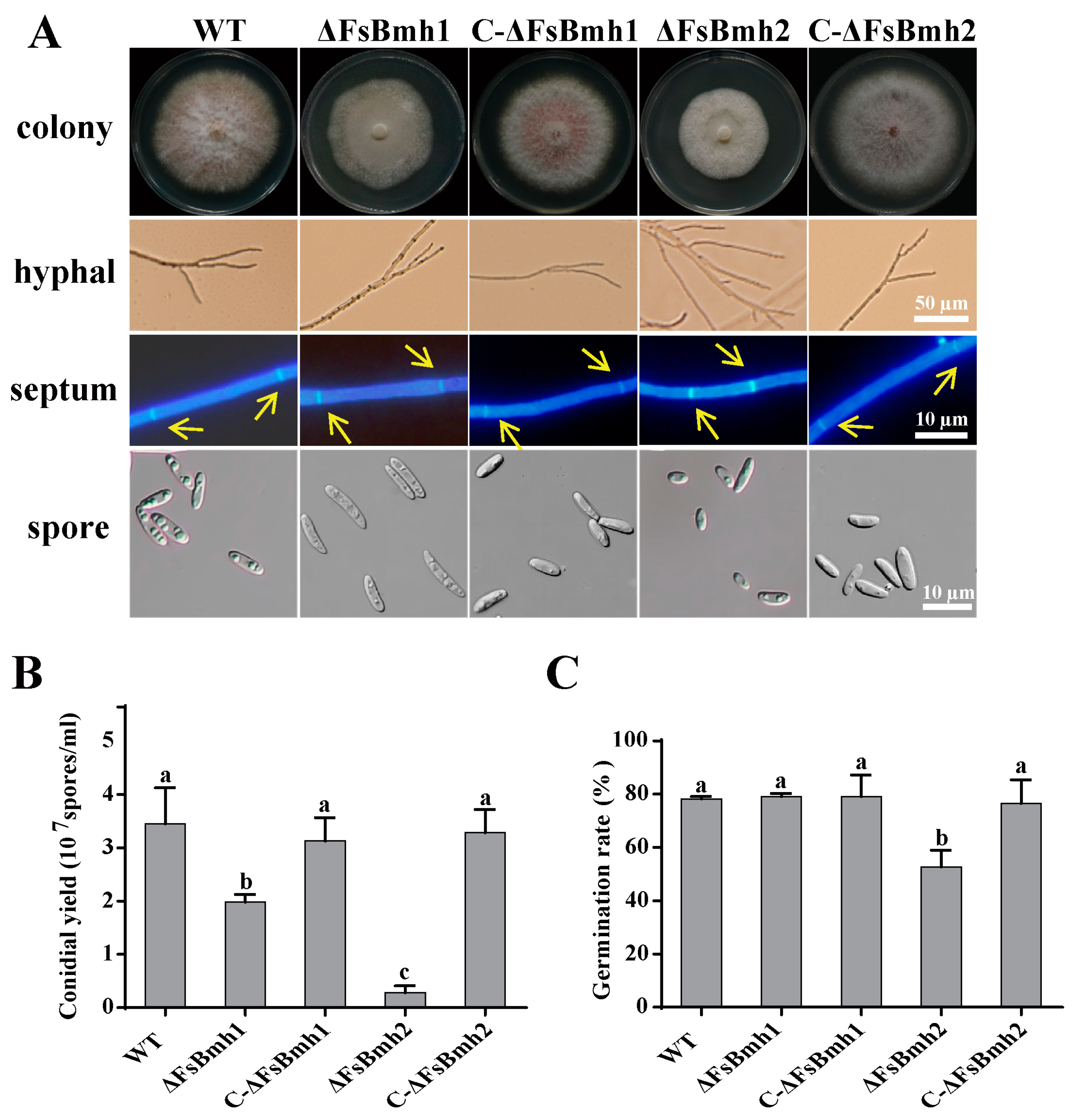
3.4. Diverse Contributions of FsBmh1 and FsBmh2 to Fungal Phenotypes
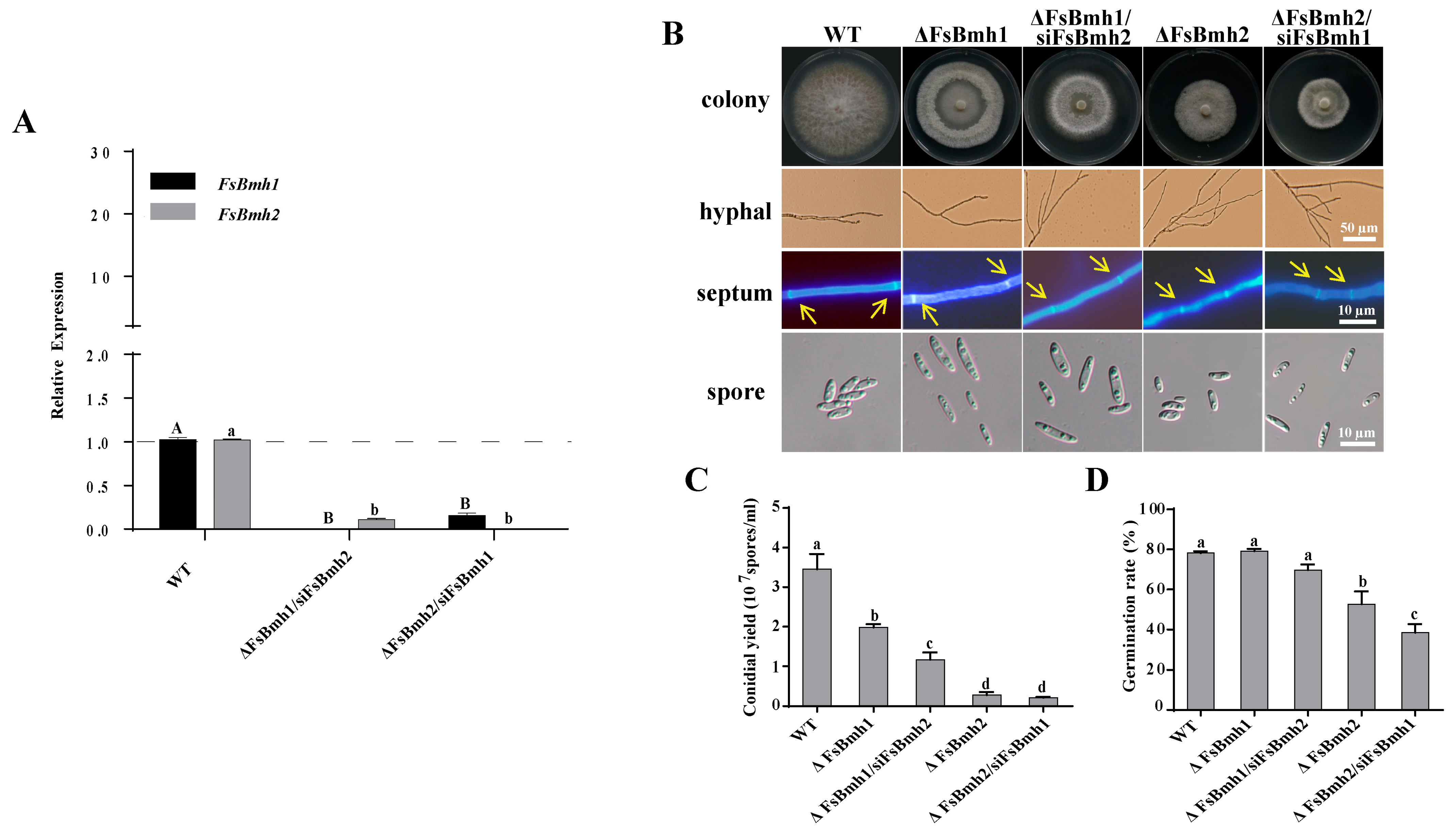
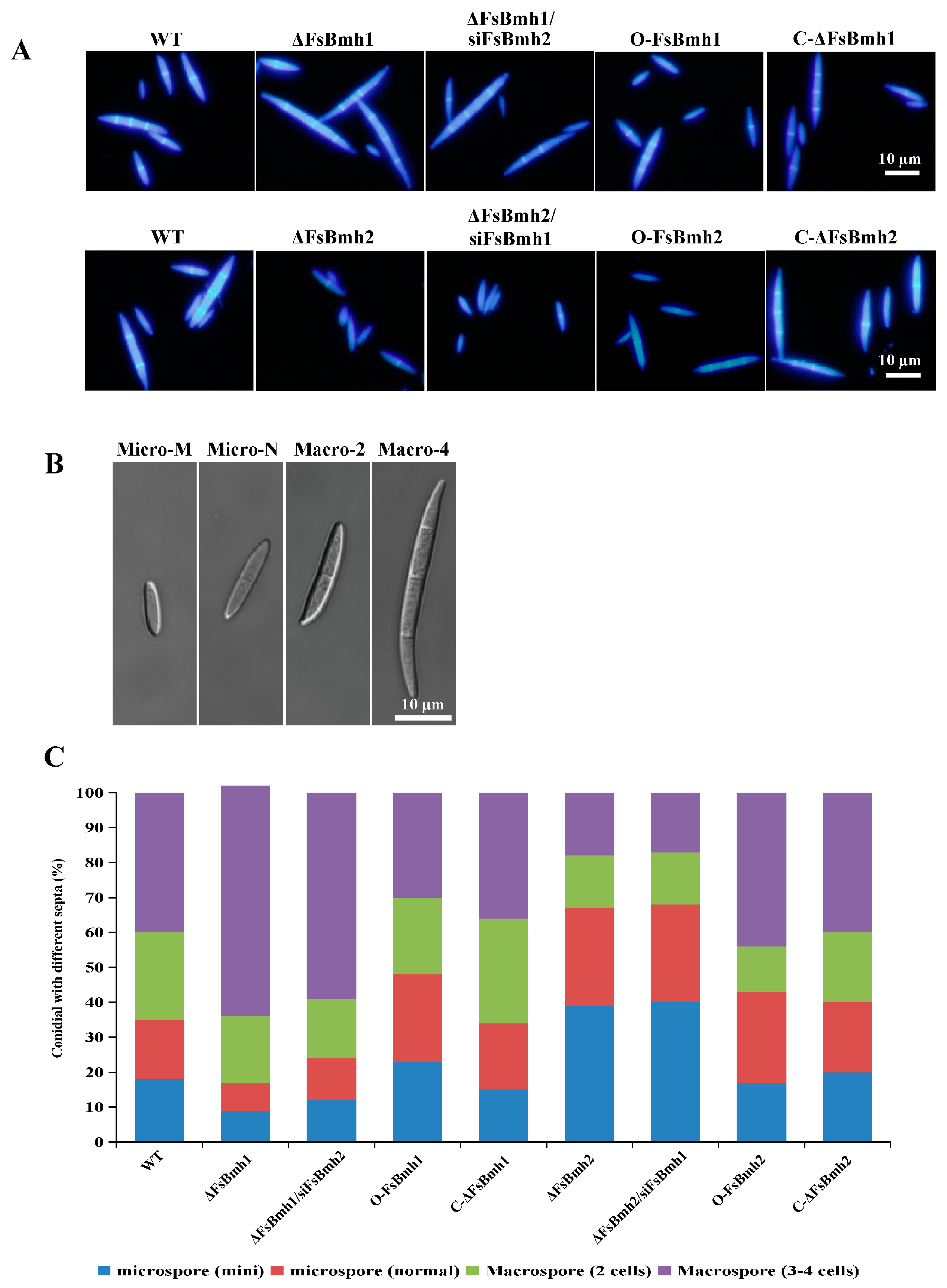
3.5. FsBmh1 and FsBmh2 Interactively Regulate Spore Morphology in F. sacchari
3.6. Overexpression of FsBmh Does Not Alter the Phenotype of F. sacchari
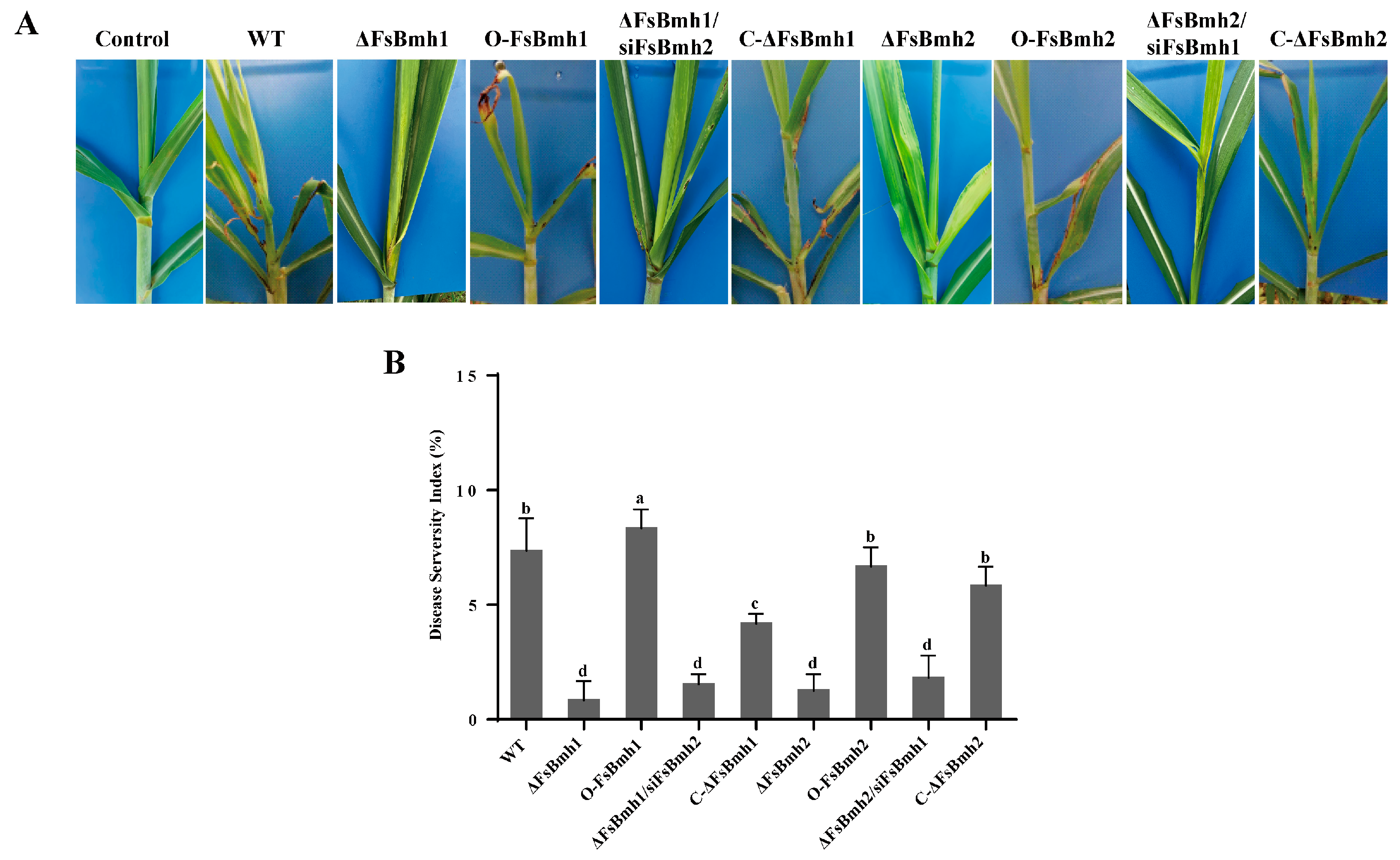
3.7. FsBmh1 and FsBmh2 Are Essential for F. sacchari Virulence
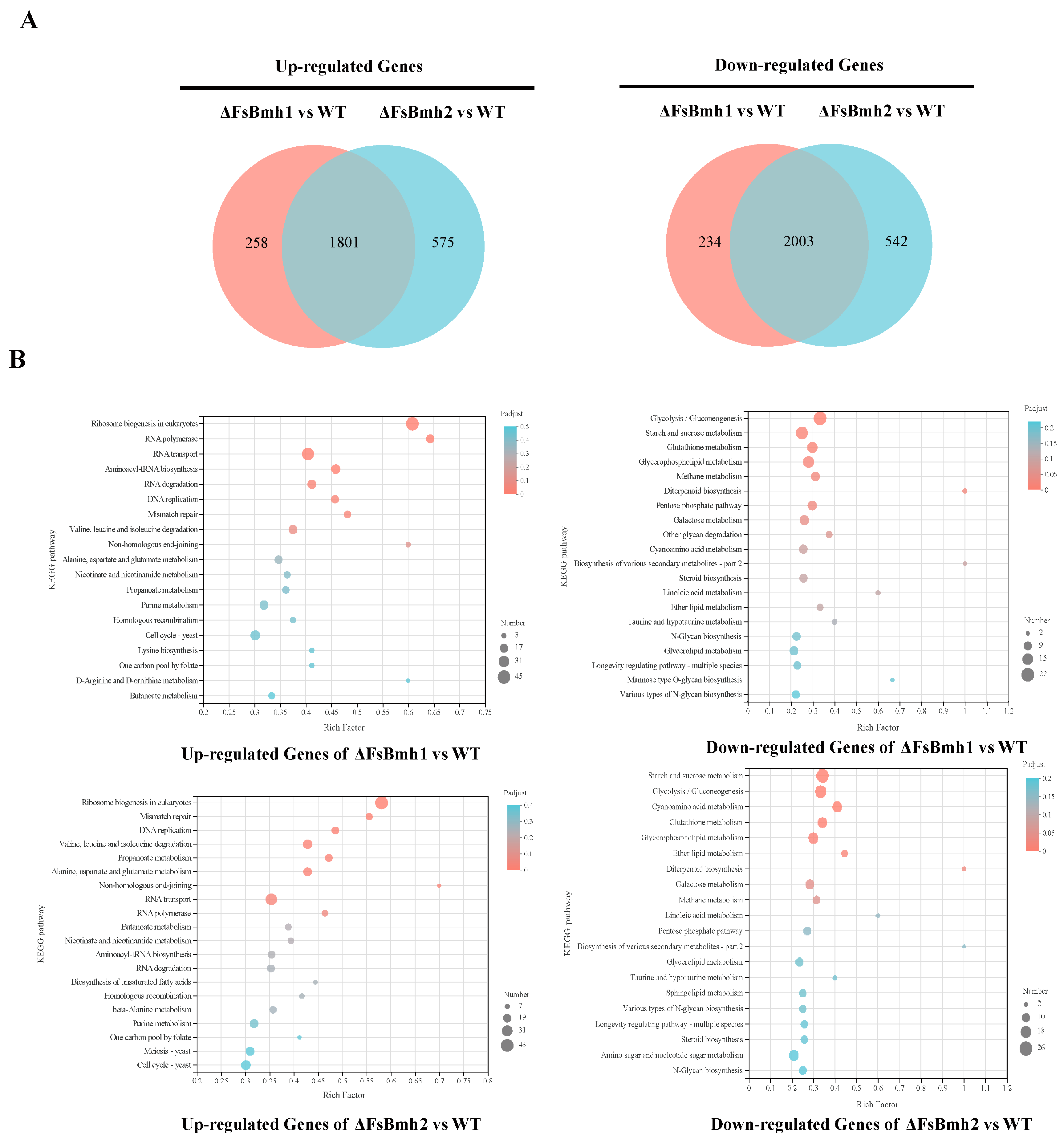
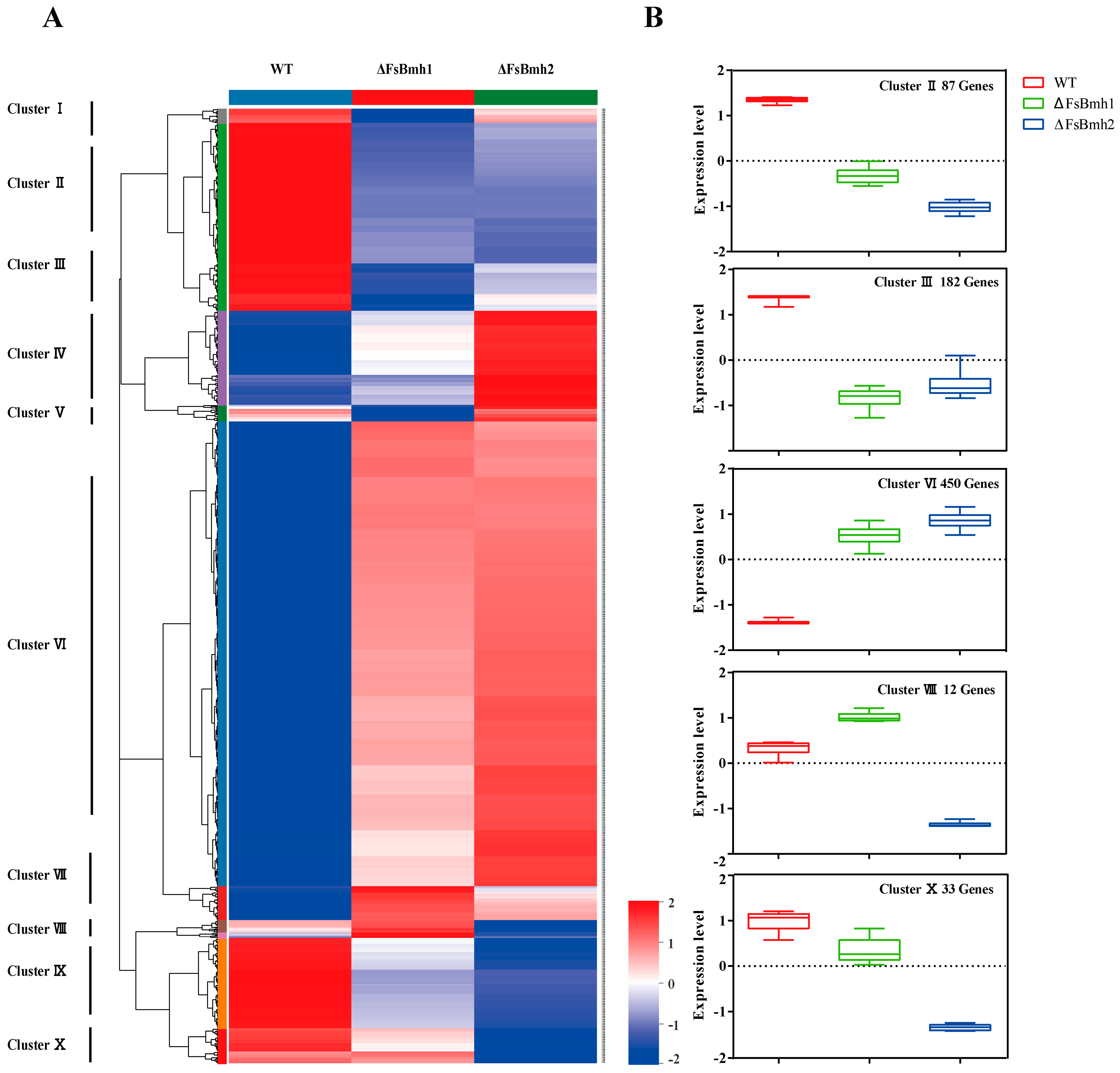
3.8. Transcriptomic Analysis Reveals the Functional Basis of FsBmh1 and FsBmh2
3.9. Identification of Direct Targets of FsBmh1 and FsBmh2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vishwakarma, S.K.; Kumar, P.; Nigam, A.; Singh, A.; Kumar, A. Pokkah Boeng: An Emerging Disease of Sugarcane. J. Plant Pathol. Microbiol. 2017, 4, 1000170. [Google Scholar]
- Lin, Z.; Xu, S.; Que, Y.; Wang, J.; Comstock, J.C.; Wei, J.; McCord, P.H.; Chen, B.; Chen, R.; Zhang, M. Species-specific detection and identification of Fusarium species complex, the causal agent of sugarcane pokkah boeng in China. PLoS ONE 2014, 9, e104195. [Google Scholar] [CrossRef] [PubMed]
- Hilton, A.; Zhang, H.; Yu, W.; Shim, W. Identification and characterization of pathogenic and endophytic fungal species associated with pokkah boeng disease of sugarcane. Plant Pathol. J. 2017, 33, 238–248. [Google Scholar] [CrossRef]
- Meng, J.R.; Huang, H.J.; Li, Y.X.; Li, Y.J.; Li, J.Q.; Chen, B.S. First report of Fusarium sacchari causing sugarcane pokkah boeng in china. Plant Dis. 2020, 104, 1553–1554. [Google Scholar] [CrossRef]
- Wang, Z.P.; Sun, H.J.; Guo, Q.; Xu, S.Q.; Wang, J.H.; Lin, S.H.; Zhang, M.Q. Artificial inoculation method of pokkah boeng disease of sugarcane and screening of resistant germplasm resources in subtropical china. Sugar Tech. 2017, 19, 283–292. [Google Scholar] [CrossRef]
- Lin, Z.; Wang, J.; Bao, Y.; Guo, Q.; Powell, C.A.; Xu, S.; Chen, B.; Zhang, M. Deciphering the transcriptomic response of Fusarium verticillioides in relation to nitrogen availability and the development of sugarcane pokkah boeng disease. Sci. Rep. 2016, 6, 29692. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Li, H.; Zhou, Y.; Bao, Y.; Duan, Z.; Wang, C.; Powell, C.A.; Chen, B.; Zhang, M.; Yao, W. Predication of the effector proteins secreted by Fusarium sacchari using genomic analysis and heterogenous expression. J. Fungi 2022, 8, 59. [Google Scholar] [CrossRef]
- Van Heusden, G.P.; Steensma, H.Y. Yeast 14-3-3 proteins. Yeast 2006, 23, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M.; Leptourgidou, F.; Camerini, S.; Pozio, E.; Skoulakis, E.M. Interkingdom complementation reveals structural conservation and functional divergence of 14-3-3 proteins. PLoS ONE 2013, 8, e78090. [Google Scholar] [CrossRef]
- Aitken, A. Post-translational modification of 14-3-3 isoforms and regulation of cellular function. Semin. Cell Dev. Biol. 2011, 22, 673–680. [Google Scholar] [CrossRef]
- Cheah, P.S.; Ramshaw, H.S.; Thomas, P.Q.; Toyo-Oka, K.; Xu, X.; Martin, S.; Coyle, P.; Guthridge, M.A.; Stomski, F.; van den Buuse, M.; et al. Neurodevelopmental and neuropsychiatric behaviour defects arise from 14-3-3zeta deficiency. Mol. Psychiatry 2012, 17, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Obsilova, V.; Kopecka, M.; Kosek, D.; Kacirova, M.; Kylarova, S.; Rezabkova, L.; Obsil, T. Mechanisms of the 14-3-3 protein function: Regulation of protein function through conformational modulation. Physiol. Res. 2014, 63, S155–S164. [Google Scholar] [CrossRef] [PubMed]
- Teunissen, J.H.M.; Crooijmans, M.E.; Teunisse, P.P.P.; van Heusden, G.P.H. Lack of 14-3-3 proteins in Saccharomyces cerevisiae results in cell-to-cell heterogeneity in the expression of Pho4-regulated genes SPL2 and PHO84. BMC Genom. 2017, 18, 701. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, M.B.; Rittinger, K.; Volinia, S.; Caron, P.R.; Aitken, A.; Leffers, H.; Gamblin, S.J.; Smerdon, S.J.; Cantley, L.C. The Structural Basis for 14-3-3: Phosphopeptide Binding Specificity. Cell 1997, 91, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shakes, D.C. Molecular evolution of the 14-3-3 protein family. J. Mol. Evol. 1996, 43, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R. An account of fungal 14-3-3 proteins. Eur. J. Cell Biol. 2017, 96, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Ren, A.; Zhu, J.; Yu, H.; Jiang, A.; Zheng, H.; Zhao, M. 14-3-3 Proteins: A window for a deeper understanding of fungal metabolism and development. World J. Microbiol. Biotechnol. 2019, 35, 24. [Google Scholar] [CrossRef]
- Camoni, L.; Visconti, S.; Aducci, P.; Marra, M. 14-3-3 proteins in plant hormone signaling: Doing several things at once. Front. Plant Sci. 2018, 9, 297. [Google Scholar] [CrossRef]
- Bertram, P.G.; Zeng, C.; Thorson, J.; Shaw, A.S.; Zheng, X.F.S. The 14-3-3 proteins positively regulate rapamycin-sensitive signaling. Curr. Biol. 1998, 8, 1259, S1–S1267. [Google Scholar] [CrossRef]
- Wang, Y.K.; Das, B.; Huber, D.H.; Wellington, M.; Kabir, M.A.; Sherman, F.; Rustchenko, E. Role of the 14–3–3 protein in carbon metabolism of the pathogenic yeast Candida albicans. Yeast 2004, 21, 685–702. [Google Scholar] [CrossRef]
- Pham, C.D.; Yu, Z.; Sandrock, B.; Bölker, M.; Gold, S.E.; Perlin, M.H. Ustilago maydis Rho1 and 14-3-3 homologues participate in pathways controlling cell separation and cell polarity. Eukaryot Cell 2009, 8, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Brauer, E.K.; Manes, N.; Bonner, C.; Subramaniam, R. Two 14-3-3 proteins contribute to nitrogen sensing through the TOR and glutamine synthetase-dependent pathways in Fusarium graminearum. Fungal Genet. Biol. 2020, 134, 103277. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liu, L.; Li, Y.; Dong, X.; Zhang, H.; Chen, H.; Zheng, X.; Zhang, Z. The FgVps39-FgVam7-FgSso1 complex mediates vesicle trafficking and is important for the development and virulence of Fusarium graminearum. Mol. Plant-Microbe Interact.® 2017, 30, 410–422. [Google Scholar] [CrossRef]
- Yu, J.H.; Hamari, Z.; Han, K.H.; Seo, J.A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Zou, C.; Zhou, H.; Wang, J.; Lu, L.; Li, Y.; Chen, B. Δ1-pyrroline-5-carboxylate/glutamate biogenesis is required for fungal virulence and sporulation. PLoS ONE 2013, 8, e73483. [Google Scholar] [CrossRef]
- Kim, E.J.; Kramer, S.F.; Hebert, C.G.; Valdes, J.J.; Bentley, W.E. Metabolic engineering of the baculovirus-expression system via inverse “shotgun” genomic analysis and RNA interference (dsRNA) increases product yield and cell longevity. Biotechnol. Bioeng. 2007, 98, 645–654. [Google Scholar] [CrossRef]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2−ΔΔCT method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Vega Gutiérrez, T.A.; Tirado Ramírez, M.A.; Molina Cárdenas, L.; López Urquídez, G.A.; López Orona, C.A. Fusarium verticillioides Causing Root and Stem Rot in Papaya (Carica papaya) in Mexico. Plant Dis. 2023, 107, 2517. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, L.I.; Liu, C.; Sun, S.; Liu, A.; Liang, Y.; Yu, J. The roles of FgPEX2 and FgPEX12 in virulence and lipid metabolism in Fusarium graminearum. Fungal Genet. Biol. 2020, 135, 103288. [Google Scholar] [CrossRef]
- Raja, S.; Subhashini, P.; Thangaradjou, T. Differential methods of localisation of fungal endophytes in the seagrasses. Mycology 2016, 7, 112–123. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- White, M.A. The yeast two-hybrid system: Forward and reverse. Proc. Natl. Acad. Sci. USA 1996, 93, 10001–10003. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Song, J.; Xin, X.; Xie, X.; Zhao, B. Arbuscular Mycorrhizal Fungal 14-3-3 Proteins Are Involved in Arbuscule Formation and Responses to Abiotic Stresses During AM Symbiosis. Front. Microbiol. 2018, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, B.A.; Lohmar, J.M.; Satterlee, T.; McDonald, T.; Cary, J.W.; Calvo, A.M. The 14-3-3 Protein Homolog ArtA Regulates Development and Secondary Metabolism in the Opportunistic Plant Pathogen Aspergillus flavus. Appl. Environ. Microbiol. 2018, 84, e02241-17. [Google Scholar] [CrossRef] [PubMed]
- Zahrádka, J.; van Heusden, G.P.; Sychrová, H. Yeast 14-3-3 proteins participate in the regulation of cell cation homeostasis via interaction with Nha1 alkali-metal-cation/proton antiporter. Biochim. Biophys. Acta 2012, 1820, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.L.; Mösch, H.U.; Fink, G.R. 14-3-3 proteins are essential for RAS/MAPK cascade signaling during pseudohyphal development in S. cerevisiae. Cell 1997, 89, 1055–1065. [Google Scholar] [CrossRef]
- Li, J.; Chang, Y.C.; Wu, C.H.; Liu, J.; Kwon-Chung, K.J.; Huang, S.H.; Shimada, H.; Fante, R.; Fu, X.; Jong, A. The 14-3-3 Gene Function of Cryptococcus neoformans Is Required for its Growth and Virulence. J. Microbiol. Biotechnol. 2016, 26, 918–927. [Google Scholar] [CrossRef]
- Nowak, M.A.; Boerlijst, M.C.; Cooke, J.; Smith, J.M. Evolution of genetic redundancy. Nature 1997, 388, 167–171. [Google Scholar] [CrossRef]
- Liu, Q.; Li, J.G.; Ying, S.H.; Wang, J.J.; Sun, W.L.; Tian, C.G.; Feng, M.G. Unveiling equal importance of two 14-3-3 proteins for morphogenesis, conidiation, stress tolerance and virulence of an insect pathogen. Environ. Microbiol. 2015, 17, 1444–1462. [Google Scholar] [CrossRef]
- Zhang, T.J.; Shi, L.; Chen, D.D.; Liu, R.; Shi, D.K.; Wu, C.G.; Sun, Z.H.; Ren, A.; Zhao, M.W. 14-3-3 proteins are involved in growth, hyphal branching, ganoderic acid biosynthesis, and response to abiotic stress in Ganoderma lucidum. Microbiol. Biotechnol. 2018, 102, 1769–1782. [Google Scholar] [CrossRef] [PubMed]
- Boulette, M.L.; Baynham, P.J.; Jorth, P.A.; Kukavica-Ibrulj, I.; Longoria, A.; Barrera, K.; Levesque, R.C.; Whiteley, M. Characterization of alanine catabolism in Pseudomonas aeruginosa and its importance for proliferation in vivo. J. Bacteriol. 2009, 191, 6329–6334. [Google Scholar] [CrossRef]
- Trevisson, E.; Burlina, A.; Doimo, M.; Pertegato, V.; Casarin, A.; Cesaro, L.; Navas, P.; Basso, G.; Sartori, G.; Salviati, L. Functional complementation in yeast allows molecular characterization of missense argininosuccinate lyase mutations. J. Biol. Chem. 2009, 284, 28926–28934. [Google Scholar] [CrossRef] [PubMed]
- Morel, M.; Buée, M.; Chalot, M.; Brun, A. NADP-dependent glutamate dehydrogenase: A dispensable function in ectomycorrhizal fungi. New Phytol. 2006, 169, 179–189. [Google Scholar] [CrossRef] [PubMed]
- King, W.R.; Singer, J.; Warman, M.; Wilson, D.; Hube, B.; Lager, I.; Patton-Vogt, J. The glycerophosphocholine acyltransferase Gpc1 contributes to phosphatidylcholine biosynthesis, long-term viability, and embedded hyphal growth in Candida albicans. J. Biol. Chem. 2024, 300, 105543. [Google Scholar] [CrossRef] [PubMed]
- Sosso, D.; van der Linde, K.; Bezrutczyk, M.; Schuler, D.; Schneider, K.; Kämper, J.; Walbot, V. Sugar Partitioning between Ustilago maydis and Its Host Zea mays L during Infection. Plant Physiol. 2019, 179, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14-3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Dengler, L.; Örd, M.; Schwab, L.M.; Loog, M.; Ewald, J.C. Regulation of trehalase activity by multi-site phosphorylation and 14-3-3 interaction. Sci. Rep. 2021, 11, 962. [Google Scholar] [CrossRef] [PubMed]
- Panni, S.; Montecchi-Palazzi, L.; Kiemer, L.; Cabibbo, A.; Paoluzi, S.; Santonico, E.; Landgraf, C.; Volkmer-Engert, R.; Bachi, A.; Castagnoli, L.; et al. Combining peptide recognition specificity and context information for the prediction of the 14-3-3-mediated interactome in S. cerevisiae and H. sapiens. Proteomics 2011, 11, 128–143. [Google Scholar] [CrossRef]
- May, T.; Soll, J. 14-3-3 proteins form a guidance complex with chloroplast precursor proteins in plants. Plant Cell 2000, 12, 53–64. [Google Scholar] [CrossRef]
- Wu, L.; Yuan, Z.; Wang, P.; Mao, X.; Zhou, M.; Hou, Y. The plasma membrane H+-ATPase FgPMA1 regulates the development, pathogenicity, and phenamacril sensitivity of Fusarium graminearum by interacting with FgMyo-5 and FgBmh2. Mol. Plant Pathol. 2022, 23, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Baltussen, T.J.H.; Coolen, J.P.M.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Gene co-expression analysis identifies gene clusters associated with isotropic and polarized growth in Aspergillus fumigatus conidia. Fungal Genet. Biol. 2018, 116, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Thakur, R.; Goel, G.; Shankar, J. Nano-LC-Q-TOF Analysis of Proteome Revealed Germination of Aspergillus flavus Conidia is Accompanied by MAPK Signalling and Cell Wall Modulation. Mycopathologia 2016, 181, 769–786. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, M.; Wang, W.; Li, X.; Bai, N.; Xie, M.; Yang, J. AoMae1 Regulates Hyphal Fusion, Lipid Droplet Accumulation, Conidiation, and Trap Formation in Arthrobotrys oligospora. J. Fungi 2023, 9, 496. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, B.; Liu, L.; Chen, H.; Zhang, H.; Zheng, X.; Zhang, Z. FgMon1, a guanine nucleotide exchange factor of FgRab7, is important for vacuole fusion, autophagy and plant infection in Fusarium graminearum. Sci. Rep. 2015, 5, 18101. [Google Scholar] [CrossRef]
- Gai, Y.; Li, L.; Ma, H.; Riely, B.K.; Liu, B.; Li, H. The critical role of MetR/MetB/MetC/MetX in cysteine and methionine metabolism, fungal development and virulence of Alternaria alternata. Appl. Environ. Microbiol. 2021, 87, e01911-20. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Zhang, Q.; Scharfenstein, L.; Mack, B.; Yoshimi, A.; Miyazawa, K.; Abe, K. Aspergillus flavus GPI-anchored protein-encoding ecm33 has a role in growth, development, aflatoxin biosynthesis, and maize infection. Appl. Microbiol. Biotechnol. 2018, 102, 5209–5220. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhang, X.; Zhang, Y.; Yi, G.; Xie, J.; Viljoen, A.; Wang, W.; Mostert, D.; Fu, G.; Peng, C.; et al. FocECM33, a GPI-anchored protein, regulates vegetative growth and virulence in Fusarium oxysporum f. sp. cubense tropical race 4. Fungal Biol. 2022, 126, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Wei, X.Y.; Wang, S.J.; Wang, J.J. Characterization of trehalose-6-phosphate phosphatase in trehalose biosynthesis, asexual development, stress resistance and virulence of an insect mycopathogen. Pestic. Biochem. Physiol. 2020, 163, 185–192. [Google Scholar] [CrossRef]
- Song, X.S.; Li, H.P.; Zhang, J.B.; Song, B.; Huang, T.; Du, X.M.; Gong, A.D.; Liu, Y.K.; Feng, Y.N.; Agboola, R.S.; et al. Trehalose 6-phosphate phosphatase is required for development, virulence and mycotoxin biosynthesis apart from trehalose biosynthesis in Fusarium graminearum. Fungal Genet. Biol. 2014, 63, 24–41. [Google Scholar] [CrossRef]
- Chen, X.; Abubakar, Y.S.; Yang, C.; Wang, X.; Miao, P.; Lin, M.; Wen, Y.; Wu, Q.; Zhong, H.; Fan, Y.; et al. Trehalose Phosphate Synthase Complex-Mediated Regulation of Trehalose 6-Phosphate Homeostasis Is Critical for Development and Pathogenesis in Magnaporthe oryzae. mSystems 2021, 6, e0046221. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Protein ID | Reference | Targeted by | Functional Annotation | Identity/% | E-Value |
|---|---|---|---|---|---|---|
| 1 | FVER_00270 | XP_031079814.1 | FsBmh1 & FsBmh2 | alpha-glucosidase (maltase) | 94.44 | 0 |
| 2 | FVER_01513 | KAF5563502.1 | FsBmh1 & FsBmh2 | Arginosuccinate lyase | 98.72 | 0 |
| 3 | FVER_02281 | XP_018749177.1 | FsBmh1 & FsBmh2 | 40S ribosomal protein S25 | 100 | 0 |
| 4 | FVER_02466 | KAG5750504.1 | FsBmh1 & FsBmh2 | glycerol-3-phosphate O-acyltransferase | 98.83 | 0 |
| 5 | FVER_03743 | KAG7434925.1 | FsBmh1 & FsBmh2 | Glyceraldehyde-3phosphate dehydrogenase | 99.41 | 0 |
| 6 | FVER_04097 | AMD38891.1 | FsBmh1 & FsBmh2 | Translation elongation factor 1-alpha | 100 | 0 |
| 7 | FVER_04309 | KAI1018791.1 | FsBmh1 & FsBmh2 | Glutamate decarboxylase | 99.48 | 0 |
| 8 | FVER_04904 | XP_023433270.1 | FsBmh1 & FsBmh2 | Trehalase | 99.13 | 0 |
| 9 | FVER_06138 | KAI1031434.1 | FsBmh1 & FsBmh2 | Bifunctional purine biosynthesis protein | 99.05 | 0 |
| 10 | FVER_06338 | XP_023428086.1 | FsBmh1 & FsBmh2 | F-type H+-transporting ATPase subunit gamma | 99.67 | 0 |
| 11 | FVER_07898 | AMD38871.1 | FsBmh1 & FsBmh2 | Alpha, alpha-trehalose-phosphate synthase | 100 | 0 |
| 12 | FVER_12178 | XP_041687834.1 | FsBmh1 & FsBmh2 | Phosphotyrosine-specific protein phosphatase | 99.27 | 0 |
| 13 | FVER_09248 | XP_023430651.1 | FsBmh1 | GPI-anchored cell wall organization protein | 98.74 | 0 |
| 14 | FVER_10685 | KAF5704680.1 | FsBmh1 | S-(hydroxymethyl) glutathione dehydrogenase | 99.48 | 0 |
| 15 | FVER_13904 | KAF5653012.1 | FsBmh1 | Pyruvate decarboxylase | 99.65 | 0 |
| 16 | FVER_02283 | KAF5614968.1 | FsBmh2 | hsp70-like protein | 85.99 | 0 |
| 17 | FVER_05845 | XP_023435700.1 | FsBmh2 | uricase | 100 | 0 |
| 18 | FVER_08037 | XP_023423671.1 | FsBmh2 | meiosis 5 (Mei5) protein | 94.78 | 0 |
| 19 | FVER_11800 | KAI1029965.1 | FsBmh2 | related to helix-loop-helix protein | 96.48 | 0 |
| 20 | FVER_11869 | KAI1041413.1 | FsBmh2 | trehalose-6-phosphate phosphatase | 99.77 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Yao, Z.; Zhao, L.; Yu, M.; Chen, B.; Zou, C. Redundant and Distinct Roles of Two 14-3-3 Proteins in Fusarium sacchari, Pathogen of Sugarcane Pokkah Boeng Disease. J. Fungi 2024, 10, 257. https://doi.org/10.3390/jof10040257
Chen Y, Yao Z, Zhao L, Yu M, Chen B, Zou C. Redundant and Distinct Roles of Two 14-3-3 Proteins in Fusarium sacchari, Pathogen of Sugarcane Pokkah Boeng Disease. Journal of Fungi. 2024; 10(4):257. https://doi.org/10.3390/jof10040257
Chicago/Turabian StyleChen, Yuejia, Ziting Yao, Lixian Zhao, Mei Yu, Baoshan Chen, and Chengwu Zou. 2024. "Redundant and Distinct Roles of Two 14-3-3 Proteins in Fusarium sacchari, Pathogen of Sugarcane Pokkah Boeng Disease" Journal of Fungi 10, no. 4: 257. https://doi.org/10.3390/jof10040257
APA StyleChen, Y., Yao, Z., Zhao, L., Yu, M., Chen, B., & Zou, C. (2024). Redundant and Distinct Roles of Two 14-3-3 Proteins in Fusarium sacchari, Pathogen of Sugarcane Pokkah Boeng Disease. Journal of Fungi, 10(4), 257. https://doi.org/10.3390/jof10040257