

Transcriptome-Wide Identification and Expression Analysis of Genes Encoding Defense-Related Peptides of Filipendula ulmaria in Response to Bipolaris sorokiniana Infection
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Experimental Design
2.3. RNA Isolation
2.4. Library Construction and NGS
2.5. Sequencing Data Analysis
2.6. Identification of CRP and Pep Precursors in F. ulmaria Transcriptomes
2.7. Analysis of DEGs
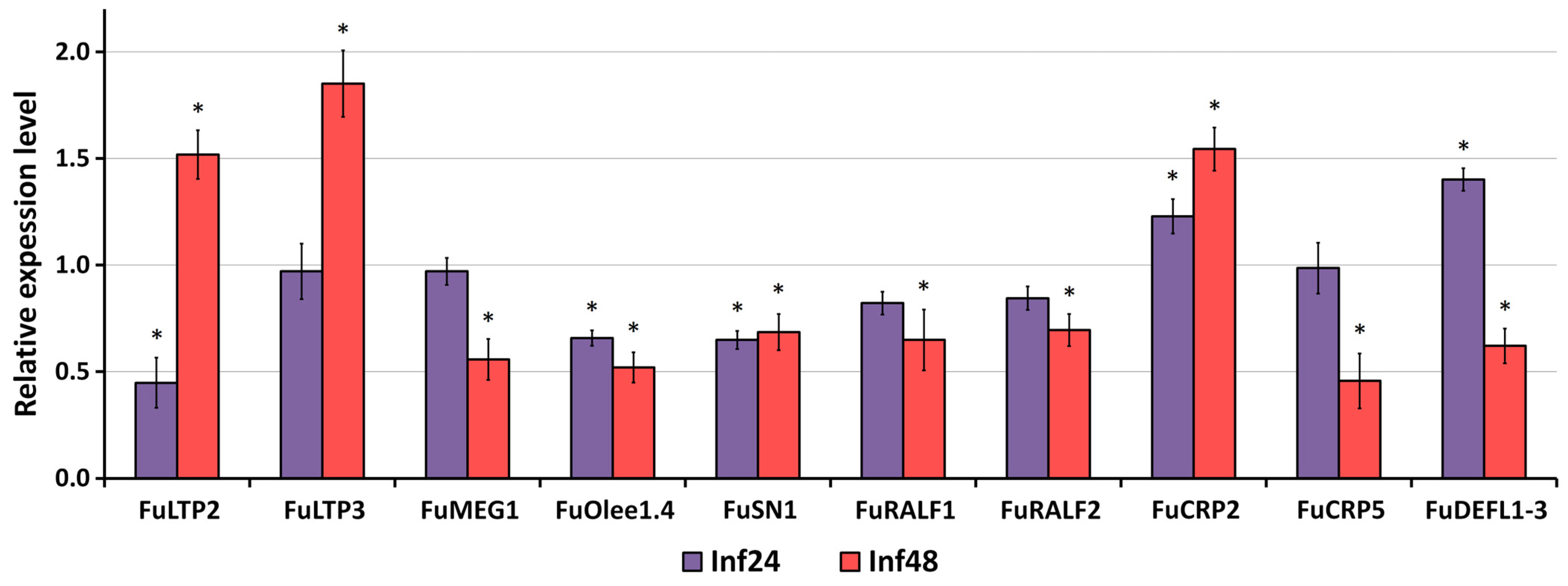
2.8. Validation of RNA-Seq Data by qRT-PCR Analysis
3. Results
3.1. Transcriptome Sequencing
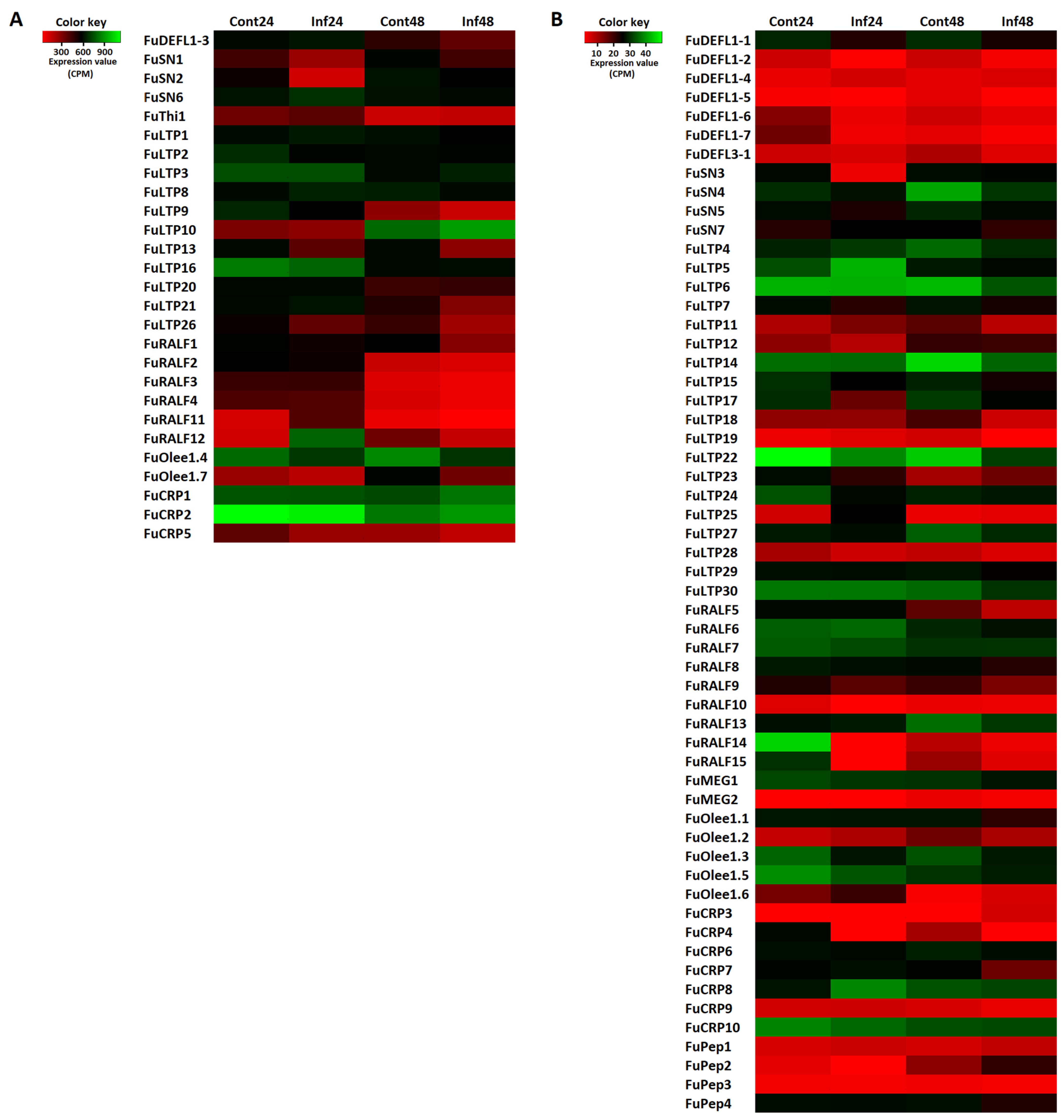
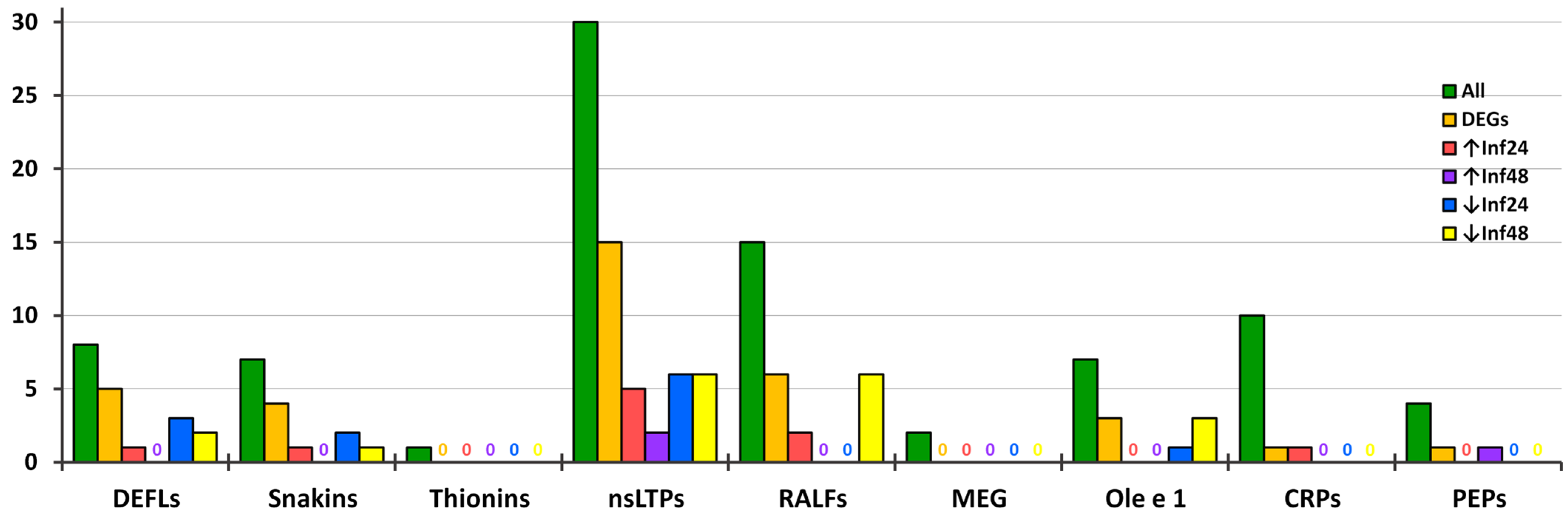
3.2. Identification of Precursors Encoding AMPs and Signaling Peptides
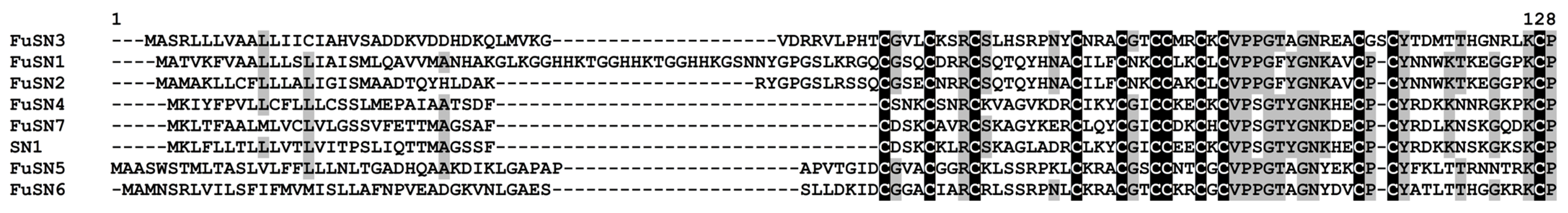
3.2.1. CRPs
- DEFLs
- Snakins
- nsLTPs
- Thionin-Like Peptides
- MEG Peptides
- Ole e 1 Peptides
- RALFs
3.2.2. CRPs with Novel Cysteine Motifs
- 4-Cys Peptides FuCRP5-10
- Other CRPs
3.2.3. Peps
3.3. Validation of RNA-Seq Data by qRT-PCR Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Savary, S.; Ficke, A.; Aubertot, J.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant pathogenic fungi. Microbiol. Spectr. 2017, 5, 701–726. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Li, S.J.; Yan, J.; Tang, Y.; Cheng, J.P.; Gao, A.J.; Yao, X.; Ruan, J.J.; Xu, B.L. Research progress on phytopathogenic fungi and their role as biocontrol agents. Front. Microbiol. 2021, 12, 670135. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Roy, C.; He, X.; Gahtyari, N.C.; Mahapatra, S.; Singh, P.K. Managing spot blotch disease in wheat: Conventional to molecular aspects. Front. Plant Sci. 2023, 14, 1098648. [Google Scholar] [CrossRef]
- Takken, F.; Rep, M. The arms race between tomato and Fusarium oxysporum. Mol. Plant Pathol. 2010, 11, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Zaslof, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 5. [Google Scholar] [CrossRef]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant antimicrobial peptides: An overview about classification, toxicity and clinical applications. Int. J. Biol. Macromol. 2022, 214, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Park, Y.; Hahm, K.S. The role of antimicrobial peptides in preventing multidrug-resistant bacterial infections and biofilm formation. Int. J. Mol. Sci. 2011, 12, 5971–5992. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini review on antimicrobial peptides, sources, mechanism and recent applications. Protein Pept. Lett. 2020, 27, 4–16. [Google Scholar] [CrossRef]
- Kanzaki, H.; Nirasawa, S.; Saitoh, H.; Ito, M.; Nishihara, M.; Terauchi, R.; Nakamura, I. Overexpression of the wasabi defensin gene confers enhanced resistance to blast fungus (Magnaporthe grisea) in transgenic rice. Theor. Appl. Genet. 2002, 105, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Chen, G.H.; Hsu, H.C.; Li, S.S.; Chen, C.S. Cloning and functional expression of a mungbean defensin VrD1 in Pichia pastoris. J. Agric. Food Chem. 2004, 52, 2256–2261. [Google Scholar] [CrossRef]
- Swathi Anuradha, T.; Divya, K.; Jami, S.K.; Kirti, P.B. Transgenic tobacco and peanut plants expressing a mustard defensin show resistance to fungal pathogens. Plant Cell Rep. 2008, 27, 1777–1786. [Google Scholar] [CrossRef] [PubMed]
- Ntui, V.O.; Thirukkumaran, G.; Azadi, P.; Khan, R.S.; Nakamura, I.; Mii, M. Stable integration and expression of wasabi defensin gene in “Egusi” melon (Colocynthis citrullus L.) confers resistance to Fusarium wilt and Alternaria leaf spot. Plant Cell Rep. 2010, 29, 943–954. [Google Scholar] [CrossRef]
- Ghag, S.B.; Shekhawat, U.K.; Ganapathi, T.R. Petunia floral defensins with unique prodomains as novel candidates for development of Fusarium wilt resistance in transgenic banana plants. PLoS ONE 2012, 7, e39557. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Fellers, J.; Adholeya, A.; Velivelli, S.L.; El-Mounadi, K.; Nersesian, N.; Clemente, T.; Shah, D. Expression of apoplast-targeted plant defensin MtDef4.2 confers resistance to leaf rust pathogen Puccinia triticina but does not affect mycorrhizal symbiosis in transgenic wheat. Trans. Res. 2016, 26, 37–49. [Google Scholar] [CrossRef]
- Chan, Y.L.; Prasad, V.; Sanjaya; Chen, K.H.; Liu, P.C.; Chan, M.T.; Cheng, C.P. Transgenic tomato plants expressing an Arabidopsis thionin (Thi2.1) driven by fruit-inactive promoter battle against phytopathogenic attack. Planta 2005, 221, 386–393. [Google Scholar] [CrossRef]
- Hoshikawa, K.; Ishihara, G.; Takahashi, H.; Nakamura, I. Enhanced resistance to gray mold (Botrytis cinerea) in transgenic potato plants expressing thionin genes isolated from Brassicaceae species. Plant Biotechnol. 2012, 29, 87–93. [Google Scholar] [CrossRef]
- Muramoto, N.; Tanaka, T.; Shimamura, T.; Mitsukawa, N.; Hori, E.; Koda, K.; Otani, M.; Hirai, M.; Nakamura, K.; Imaeda, T. Transgenic sweet potato expressing thionin from barley gives resistance to black rot disease caused by Ceratocystis fimbriata in leaves and storage roots. Plant Cell Rep. 2012, 31, 987–997. [Google Scholar] [CrossRef]
- Koo, J.C.; Chun, H.J.; Park, H.C.; Kim, M.C.; Koo, Y.D.; Koo, S.C.; Ok, H.M.; Park, S.J.; Lee, S.H.; Yun, D.J.; et al. Over-expression of a seed specific hevein-like antimicrobial peptide from Pharbitis nil enhances resistance to a fungal pathogen in transgenic tobacco plants. Plant Mol. Biol. 2002, 50, 441–452. [Google Scholar] [CrossRef]
- Shukurov, R.R.; Voblikova, V.D.; Nikonorova, A.K.; Komakhin, R.A.; Komakhina, V.V.; Egorov, T.A.; Grishin, E.V.; Babakov, A.V. Transformation of tobacco and Arabidopsis plants with Stellaria media genes encoding novel hevein-like peptides increases their resistance to fungal pathogens. Transgenic Res. 2012, 21, 313–325. [Google Scholar] [CrossRef]
- Chen, H.; Guo, A.; Lu, Z.; Tan, S.; Wang, J.; Gao, J.; Zhang, S.; Huang, X.; Zheng, J.; Xi, J.; et al. Agrobacterium tumefaciens-mediated transformation of a hevein-like gene into asparagus leads to stem wilt resistance. PLoS ONE 2019, 14, e0223331. [Google Scholar] [CrossRef] [PubMed]
- Almasia, N.I.; Bazzini, A.A.; Hopp, H.E.; Vazquez-Rovere, C. Overexpression of snakin-1 gene enhances resistance to Rhizoctonia solani and Erwinia carotovora in transgenic potato plants. Mol. Plant Pathol. 2008, 9, 329–338. [Google Scholar] [CrossRef]
- Darqui, F.S.; Radonic, L.M.; Trotz, P.M.; López, N.; Vázquez Rovere, C.; Hopp, H.E.; López Bilbao, M. Potato snakin-1 gene enhances tolerance to Rhizoctonia solani and Sclerotinia sclerotiorum in transgenic lettuce plants. J. Biotechnol. 2018, 283, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Conti, G.; Gardella, V.; Vandecaveye, M.A.; Gomez, C.A.; Joris, G.; Hauteville, C.; Burdyn, L.; Almasia, N.I.; Nahirñak, V.; Vazquez-Rovere, C.; et al. Transgenic Citrange troyer rootstocks overexpressing antimicrobial potato Snakin-1 show reduced citrus canker disease symptoms. J. Biotechnol. 2020, 324, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Cao, M.Y.; Zhu, P.X.; Zhang, Q.P.; Lam, H.M. Non-specific LIPID TRANSFER PROTEIN 1 enhances immunity against tobacco mosaic virus in Nicotiana benthamiana. J. Exp. Bot. 2023, 74, 5236–5254. [Google Scholar] [CrossRef]
- Tian, N.; Liu, F.; Wang, P.; Yan, X.; Gao, H.; Zeng, X.; Wu, G. Overexpression of BraLTP2, a lipid transfer protein of Brassica napus, results in increased trichome density and altered concentration of secondary metabolites. Int. J. Mol. Sci. 2018, 19, 1733. [Google Scholar] [CrossRef]
- Jia, Z.; Gou, J.; Sun, Y.; Yuan, L.; Tang, Q.; Yang, X.; Pei, Y.; Luo, K. Enhanced resistance to fungal pathogens in transgenic Populus tomentosa Carr. by overexpression of an nsLTP-like antimicrobial protein gene from motherwort (Leonurus japonicus). Tree Physiol. 2010, 30, 1599–1605. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Tan, H.; Dai, Y.; Li, L.; Huang, Y.; Yao, H.; Cai, Y.; Yu, G. Application of antimicrobial peptides in plant protection: Making use of the overlooked merits. Front. Plant Sci. 2023, 14, 1139539. [Google Scholar] [CrossRef]
- De Coninck, B.; De Smet, I. Plant peptides—Taking them to the next level. J. Exp. Bot. 2016, 67, 4791–4795. [Google Scholar] [CrossRef] [PubMed]
- Luo, L. Plant cytokine or phytocytokine. Plant Signal. Behav. 2012, 7, 1513–1514. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Liu, D.; He, P. Phytocytokines function as immunological modulators of plant immunity. Stress Biol. 2021, 1, 8. [Google Scholar] [CrossRef]
- Huffaker, A.; Pearce, G.; Veyrat, N.; Erb, M.; Turlings, T.C.; Sartor, R.; Shen, Z.; Briggs, S.P.; Vaughan, M.M.; Alborn, H.T.; et al. Plant elicitor peptides are conserved signals regulating direct and indirect antiherbivore defense. Proc. Natl. Acad. Sci. USA 2013, 110, 5707–5712. [Google Scholar] [CrossRef]
- Foix, L.; Nadal, A.; Zagorščak, M.; Ramšak, Ž.; Esteve-Codina, A.; Gruden, K.; Pla, M. Prunus persica plant endogenous peptides PpPep1 and PpPep2 cause PTI-like transcriptome reprogramming in peach and enhance resistance to Xanthomonas arboricola pv. pruni. BMC Genom. 2021, 22, 360. [Google Scholar] [CrossRef]
- Al-Sadi, A.M. Bipolaris sorokiniana-Induced black point, common root rot, and spot blotch diseases of wheat: A review. Front. Cell. Infect. Microbiol. 2021, 11, 584899. [Google Scholar] [CrossRef]
- Kiseleva, M.I.; Zhemchuzhina, N.S.; Dubovoi, V.P.; Lapina, V.V. Identification of root rot pathogens isolated on spring grain crops in republic of Mordovia. Sel’skokhozyaistvennaya Biol. [Agric. Biol.] 2016, 51, 119–127. [Google Scholar] [CrossRef][Green Version]
- Ullah, H.; Yasmin, H.; Mumtaz, S.; Jabeen, Z.; Naz, R.; Nosheen, A.; Hassan, M.N. Multitrait Pseudomonas spp. isolated from monocropped wheat (Triticum aestivum) suppress Fusarium root and crown rot. Phytopathology 2020, 110, 582–592. [Google Scholar] [CrossRef]
- Allali, K.; Goudjal, Y.; Zamoum, M.; Bouznada, K.; Sabaou, N.; Zitouni, A. Nocardiopsis dassonvillei strain MB22 from the Algerian Sahara promotes wheat seedlings growth and potentially controls the common root rot pathogen Bipolaris sorokiniana. J. Plant Pathol. 2019, 101, 1115–1125. [Google Scholar] [CrossRef]
- Campanella, V.; Mandalà, C.; Angileri, V.; Miceli, C. Management of common root rot and Fusarium foot rot of wheat using Brassica carinata break crop green manure. Crop Prot. 2020, 130, 105073. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Kumar, M.; Sahu, P.K.; Singh, H.V.; Kumar, S.; Roy, M.; Imran, M.; Rai, J.P.; et al. Trichoderma harzianum- and methyl jasmonate-induced resistance to Bipolaris sorokiniana through enhanced phenylpropanoid activities in bread wheat (Triticum aestivum L.). Front. Microbiol. 2019, 10, 1697. [Google Scholar] [CrossRef]
- Sharma, S.; Sahu, R.; Navathe, S.; Mishra, V.K.; Chand, R.; Singh, P.K.; Joshi, A.K.; Pandey, S.P. Natural variation in elicitation of defense-signaling associates to field resistance against the spot blotch disease in bread wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 636. [Google Scholar] [CrossRef]
- Shahbaz, M.; Riaz, M.; Ali, S.; Ahmad, F.; Hussain, A.; Nabi, G.; Chaudhry, M.T.; Muhammad, S. Effect of seed dressing chemicals on emergence, yield and against soil & seed born diseases of wheat. Pak. J. Phytopathol. 2018, 30, 183–189. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. Meadowsweet teas as new functional beverages: Comparative analysis of nutrients, phytochemicals and biological effects of four Filipendula species. Molecules 2016, 22, 16. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oliveira, P.; Fraga-Corral, M.; Pereira, A.G.; Lourenço-Lopes, C.; Jimenez-Lopez, C.; Prieto, M.A.; Simal-Gandara, J. Scientific basis for the industrialization of traditionally used plants of the Rosaceae family. Food Chem. 2020, 330, 127197. [Google Scholar] [CrossRef] [PubMed]
- Cwikla, C.; Schmidt, K.; Matthias, A.; Bone, K.M.; Lehmann, R.; Tiralongo, E. Investigations into the antibacterial activities of phytotherapeutics against Helicobacter pylori and Campylobacter jejuni. Phytother. Res. 2010, 24, 649–656. [Google Scholar] [CrossRef]
- Pushkareva, V.I.; Slezina, M.P.; Korostyleva, T.V.; Shcherbakova, L.A.; Istomina, E.A.; Ermolaeva, S.A.; Ogarkova, O.A.; Odintsova, T.I. Antimicrobial activity of wild plant seed extracts against human bacterial and plant fungal pathogens. Am. J. Plant Sci. 2017, 8, 1572–1592. [Google Scholar] [CrossRef]
- Bijttebier, S.; Van der Auwera, A.; Voorspoels, S.; Noten, B.; Hermans, N.; Pieters, L.; Apers, S. A first step in the quest for the active constituents in Filipendula ulmaria (meadowsweet): Comprehensive phytochemical identification by liquid chromatography coupled to quadrupole-orbitrap mass spectrometry. Planta Med. 2016, 82, 559–572. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 7 November 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bushmanova, E.; Antipov, D.; Lapidus, A.; Prjibelski, A.D. rnaSPAdes: A de novo transcriptome assembler and its application to RNA-Seq data. GigaScience 2019, 8, giz100. [Google Scholar] [CrossRef]
- TransDecoder (Find Coding Regions within Transcripts). Available online: https://github.com/TransDecoder/TransDecoder/wiki (accessed on 15 January 2023).
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Kasianov, A.S.; Kovtun, A.S.; Makeev, V.J.; Shcherbakova, L.A.; Kudryavtsev, A.M. Defensin-like peptides in wheat analyzed by whole-transcriptome sequencing: A focus on structural diversity and role in induced resistance. PeerJ 2019, 7, e6125. [Google Scholar] [CrossRef]
- Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Kovtun, A.S.; Kasianov, A.S.; Konopkin, A.A.; Shcherbakova, L.A.; Odintsova, T.I. Molecular insights into the role of cysteine-rich peptides in induced resistance to Fusarium oxysporum infection in tomato based on transcriptome profiling. Int. J. Mol. Sci. 2021, 22, 5741. [Google Scholar] [CrossRef]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Gawde, U.; Chakraborty, S.; Waghu, F.H.; Barai, R.S.; Khanderkar, A.; Indraguru, R.; Shirsat, T.; Idicula-Thomas, S. CAMPR4: A database of natural and synthetic antimicrobial peptides. Nucleic Acids Res. 2023, 51, D377–D383. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, L.P. IPC—Isoelectric Point Calculator. Biol. Direct 2016, 11, 55. [Google Scholar] [CrossRef]
- Eisenhaber, B.; Wildpaner, M.; Schultz, C.J.; Borner, G.H.H.; Dupree, P.; Eisenhaber, F. Glycosylphosphatidylinositol lipid anchoring of plant proteins. Sensitive prediction from sequence- and genome-wide studies for Arabidopsis and rice. Plant Physiol. 2003, 133, 1691–1701. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCT method. Methods 2011, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Moyer, T.B.; Purvis, A.L.; Wommack, A.J.; Hicks, L.M. Proteomic response of Escherichia coli to a membrane lytic and iron chelating truncated Amaranthus tricolor defensin. BMC Microbiol. 2021, 21, 110. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.T.; Velivelli, S.L.S.; Berg, R.H.; Oakley, B.; Shah, D.M. A novel bi-domain plant defensin MtDef5 with potent broad-spectrum antifungal activity binds to multiple phospholipids and forms oligomers. Sci. Rep. 2017, 7, 16157. [Google Scholar] [CrossRef]
- Huffaker, A.; Pearce, G.; Ryan, C.A. An endogenous peptide signal in Arabidopsis activates components of the innate immune response. Proc. Natl. Acad. Sci. USA 2006, 103, 10098–100103. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.R. (Ed.) British Herbal Compendium; British Herbal Medicine Association: Aughton, UK, 1992; Volume 1, pp. 158–160. [Google Scholar]
- Farzaneh, A.; Hadjiakhoondi, A.; Khanavi, M.; Manayi, A.; Bahramsoltani, R.; Kalkhorani, M. Filipendula ulmaria (L.) Maxim. (meadowsweet): A review of traditional uses, phytochemistry and pharmacology. Res. J. Pharmacogn. 2022, 9, 85–106. [Google Scholar] [CrossRef]
- Zeylstra, H. Filipendula ulmaria . Br. J. Phytother. 1998, 5, 8–12. [Google Scholar]
- Sukhikh, S.; Ivanova, S.; Skrypnik, L.; Bakhtiyarova, A.; Larina, V.; Krol, O.; Prosekov, A.; Frolov, A.; Povydysh, M.; Babich, O. Study of the antioxidant properties of Filipendula ulmaria and Alnus glutinosa. Plants 2022, 11, 2415. [Google Scholar] [CrossRef]
- Savina, T.; Lisun, V.; Feduraev, P.; Skrypnik, L. Variation in phenolic compounds, antioxidant and antibacterial activities of extracts from different plant organs of meadowsweet (Filipendula ulmaria (L.) Maxim.). Molecules 2023, 28, 3512. [Google Scholar] [CrossRef]
- Pannakal, S.T.; Eilstein, J.; Hubert, J.; Kotland, A.; Prasad, A.; Gueguiniat-Prevot, A.; Juchaux, F.; Beaumard, F.; Seru, G.; John, S.; et al. Rapid chemical profiling of Filipendula ulmaria using CPC fractionation, 2-D mapping of 13C NMR data, and high-resolution LC-MS. Molecules 2023, 28, 6349. [Google Scholar] [CrossRef]
- Papp, I.; Simándi, B.; Blazics, B.; Alberti, Á.; Héthelyi, É.; Szöke, É.; Kéry, Á. Monitoring volatile and non-volatile salicylates in Filipendula ulmaria by different chromatographic techniques. Chromatographia 2008, 68, 125–129. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Shelenkov, A.A.; Korostyleva, T.V.; Rogozhin, E.A.; Melnikova, N.V.; Kudryavtseva, A.V.; Odintsova, T.I. Defense peptide repertoire of Stellaria media predicted by high throughput next generation sequencing. Biochimie 2017, 135, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Shelenkov, A.A.; Slavokhotova, A.A.; Odintsova, T.I. Cysmotif searcher pipeline for antimicrobial peptide identification in plant transcriptomes. Biochemistry 2018, 83, 1424–1432. [Google Scholar] [CrossRef]
- Yount, N.Y.; Yeaman, M.R. Multidimensional signatures in antimicrobial peptides. Proc. Natl Acad. Sci. USA 2004, 101, 7363–7368. [Google Scholar] [CrossRef] [PubMed]
- Slezina, M.P.; Istomina, E.A.; Odintsova, T.I. Biological diversity of genes encoding wheat defensin homologs. Rus. J. Genet. 2023, 59, 1310–1319. [Google Scholar] [CrossRef]
- Sagaram, U.S.; Pandurangi, R.; Kaur, J.; Smith, T.J.; Shah, D.M. Structure-activity determinants in antifungal plant defensins MsDef1 and MtDef4 with different modes of action against Fusarium graminearum. PLoS ONE 2011, 6, e18550. [Google Scholar] [CrossRef]
- Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Odintsova, T.I. The γ-core motif peptides of plant AMPs as novel antimicrobials for medicine and agriculture. Int. J. Mol. Sci. 2022, 24, 483. [Google Scholar] [CrossRef]
- Slezina, M.P.; Istomina, E.A.; Kulakovskaya, E.V.; Abashina, T.N.; Odintsova, T.I. Synthetic oligopeptides mimicking γ-core regions of cysteine-rich peptides of Solanum lycopersicum possess antimicrobial activity against human and plant pathogens. Curr. Issues Mol. Biol. 2021, 43, 1226–1242. [Google Scholar] [CrossRef]
- Slezina, M.P.; Istomina, E.A.; Kulakovskaya, E.V.; Korostyleva, T.V.; Odintsova, T.I. The γ-core motif peptides of AMPs from grasses display inhibitory activity against human and plant pathogens. Int. J. Mol. Sci. 2022, 23, 8383. [Google Scholar] [CrossRef]
- Pearce, G.; Moura, D.S.; Stratmann, J.; Ryan, C.A., Jr. RALF, a 5-kDa ubiquitous polypeptide in plants, arrests root growth and development. Proc. Natl. Acad. Sci. USA 2001, 98, 12843–12847. [Google Scholar] [CrossRef] [PubMed]
- Germain, H.; Chevalier, E.; Caron, S.; Matton, D.P. Characterization of five RALF-like genes from Solanum chacoense provides support for a developmental role in plants. Planta 2005, 220, 447–454. [Google Scholar] [CrossRef]
- Wu, J.; Kurten, E.L.; Monshausen, G.; Hummel, G.M.; Gilroy, S.; Baldwin, I.T. NaRALF, a peptide signal essential for the regulation of root hair tip apoplastic pH in Nicotiana attenuata, is required for root hair development and plant growth in native soils. Plant J. 2007, 52, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B.; Sussman, M.R. A peptide hormone and its receptor protein kinase regulate plant cell expansion. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Morato do Canto, A.; Ceciliato, P.H.O.; Ribeiro, B.; Ortiz Morea, F.A.; Franco Garcia, A.A.; Silva-Filho, M.C.; Moura, D.S. Biological activity of nine recombinant AtRALF peptides: Implications for their perception and function in Arabidopsis. Plant Physiol. Biochem. 2014, 75, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Mecchia, M.A.; Santos-Fernandez, G.; Duss, N.N.; Somoza, S.C.; Boisson-Dernier, A.; Gagliardini, V.; Martínez-Bernardini, A.; Fabrice, T.N.; Ringli, C.; Muschietti, J.P.; et al. RALF4/19 peptides interact with LRX proteins to control pollen tube growth in Arabidopsis. Science 2017, 358, 1600–1603. [Google Scholar] [CrossRef] [PubMed]
- Loubert-Hudon, A.; Mazin, B.D.; Chevalier, É.; Matton, D.P. The ScRALF3 secreted peptide is involved in sporophyte to gametophyte signalling and affects pollen mitosis I. Plant Biol. 2020, 22, 13–20. [Google Scholar] [CrossRef]
- Blackburn, M.R.; Haruta, M.; Moura, D.S. Twenty years of progress in physiological and biochemical investigation of RALF peptides. Plant Physiol. 2020, 182, 1657–1666. [Google Scholar] [CrossRef]
- Stegmann, M.; Monaghan, J.; Smakowska-Luzan, E.; Rovenich, H.; Lehner, A.; Holton, N.; Belkhadir, Y.; Zipfel, C. The receptor kinase FER is a RALF-regulated scaffold controlling plant immune signaling. Science 2017, 355, 287–289. [Google Scholar] [CrossRef]
- Matos, J.L.; Fiori, C.S.; Silva-Filho, M.C.; Moura, D.S. A conserved dibasic site is essential for correct processing of the peptide hormone AtRALF1 in Arabidopsis thaliana. FEBS Lett. 2008, 582, 3343–3347. [Google Scholar] [CrossRef]
- Srivastava, R.; Liu, J.X.; Guo, H.; Yin, Y.; Howell, S.H. Regulation and processing of a plant peptide hormone, AtRALF23, in Arabidopsis. Plant J. 2009, 59, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.; Lori, M.; Mbengue, M.; van Verk, M.; Klauser, D.; Hander, T.; Böni, R.; Robatzek, S.; Boller, T. The family of Peps and their precursors in Arabidopsis: Differential expression and localization but similar induction of pattern-triggered immune responses. J. Exp. Bot. 2013, 64, 5309–5321. [Google Scholar] [CrossRef]
- Pearce, G.; Yamaguchi, Y.; Munske, G.; Ryan, C.A. Structure-activity studies of AtPep1, a plant peptide signal involved in the innate immune response. Peptides 2008, 29, 2083–2089. [Google Scholar] [CrossRef]
- Lori, M.; van Verk, M.C.; Hander, T.; Schatowitz, H.; Klauser, D.; Flury, P.; Gehring, C.A.; Boller, T.; Bartels, S.J. Evolutionary divergence of the plant elicitor peptides (Peps) and their receptors: Interfamily incompatibility of perception but compatibility of downstream signalling. J. Exp. Bot. 2015, 66, 5315–5325. [Google Scholar] [CrossRef]
- Krol, E.; Mentzel, T.; Chinchilla, D.; Boller, T.; Felix, G.; Kemmerling, B.; Postel, S.; Arents, M.; Jeworutzki, E.; Al-Rasheid, K.A.; et al. Perception of the Arabidopsis danger signal peptide 1 involves the pattern recognition receptor AtPEPR1 and its close homologue AtPEPR2. J. Biol. Chem. 2010, 285, 13471–13479. [Google Scholar] [CrossRef]
- Bartels, S.; Boller, T. Quo vadis, Pep? Plant elicitor peptides at the crossroads of immunity, stress, and development. J. Exp. Bot. 2015, 66, 5183–5193. [Google Scholar] [CrossRef]
- Huffaker, A.; Dafoe, N.J.; Schmelz, E.A. ZmPep1, an ortholog of Arabidopsis elicitor peptide 1, regulates maize innate immunity and enhances disease resistance. Plant Physiol. 2011, 155, 1325–1338. [Google Scholar] [CrossRef]
- Zelman, A.K.; Berkowitz, G.A. Plant elicitor peptide (Pep) signaling and pathogen defense in tomato. Plants 2023, 12, 2856. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Lilley, C.J.; Urwin, P.E. Identification of genes involved in the response of Arabidopsis to simultaneous biotic and abiotic stresses. Plant Physiol. 2013, 162, 2028–2041. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.; Dennison, S.R.; Lea, B.; Snape, T.J.; Nicoll, L.D.; Radecka, I.; Harris, F. Anionic antimicrobial and anticancer peptides from plants. Crit. Rev. Plant Sci. 2013, 32, 303–320. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Number of Raw Assembled Contigs | Number of CDS * |
|---|---|---|
| Number of sequences | 158,615 | 46,063 |
| N50, bp | 1365 | 1221 |
| Median length, bp | 297 | 699 |
| Average length, bp | 687 | 933 |
| Maximal length, bp | 16,604 | 14,925 |
| Assembly | Complete BUSCOs (%) | Complete and Single-Copy BUSCOs (%) | Complete and Duplicated BUSCOs (%) | Fragmented BUSCOs (%) | Missing BUSCOs (%) |
|---|---|---|---|---|---|
| 1a,1b,2a,2b | 237 (55.1) | 209 (48.6) | 28 (6.5) | 116 (27.0) | 77 (17.9) |
| 3a,3b,4a,4b | 383 (89.1) | 335 (77.9) | 48 (11.2) | 29 (6.7) | 18 (4.2) |
| 1a,1b,2a,2b, 3a,3b,4a,4b | 351 (81.6) | 264 (61.4) | 87 (20.2) | 55 (12.8) | 24 (5.6) |
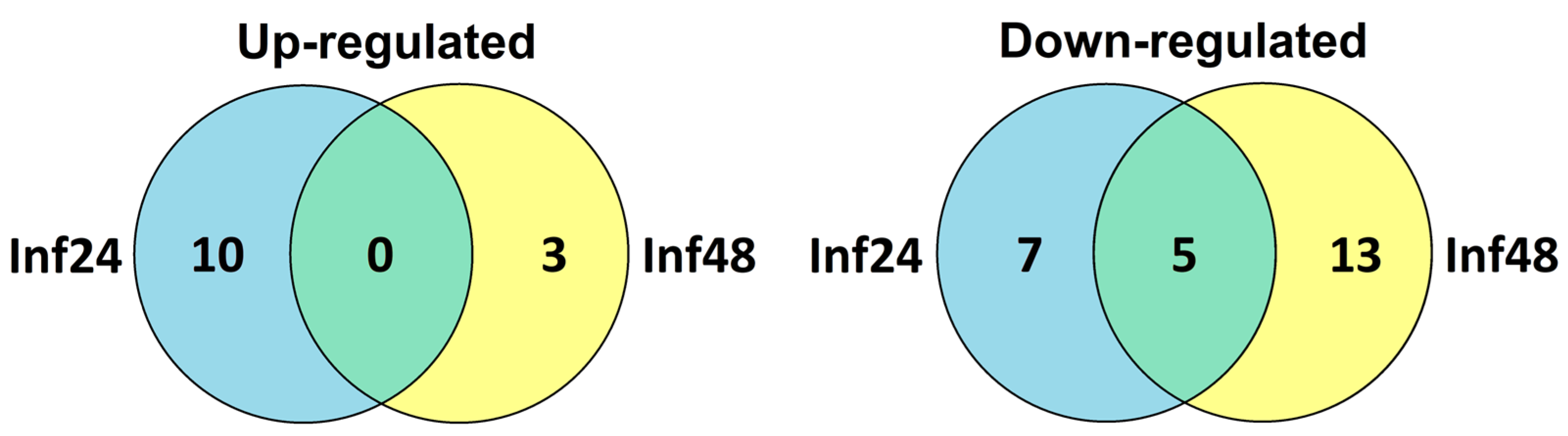
| Up-Regulated | Down-Regulated | ||
|---|---|---|---|
| at 24 hpi | at 48 hpi | at 24 hpi | at 48 hpi |
| FuDEFL1-4 | FuDEFL1-1 | FuDEFL1-1 | |
| FuDEFL1-6 | FuDEFL3-1 | ||
| FuDEFL1-7 | |||
| FuSN6 | FuSN2 | FuSN4 | |
| FuSN3 | |||
| FuLTP5 | FuLTP3 | FuLTP2 | FuLTP11 |
| FuLTP8 | FuLTP23 | FuLTP9 | FuLTP13 |
| FuLTP11 | FuLTP13 | FuLTP15 | |
| FuLTP19 | FuLTP15 | FuLTP17 | |
| FuLTP25 | FuLTP17 | FuLTP18 | |
| FuLTP24 | FuLTP22 | ||
| FuRALF11 | FuRALF1 | ||
| FuRALF12 | FuRALF5 | ||
| FuRALF11 | |||
| FuRALF12 | |||
| FuRALF14 | |||
| FuRALF15 | |||
| FuOlee1.3 | FuOlee1.1 | ||
| FuOlee1.3 | |||
| FuOlee1.4 | |||
| FuCRP8 | |||
| FuPEP2 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Istomina, E.A.; Korostyleva, T.V.; Kovtun, A.S.; Slezina, M.P.; Odintsova, T.I. Transcriptome-Wide Identification and Expression Analysis of Genes Encoding Defense-Related Peptides of Filipendula ulmaria in Response to Bipolaris sorokiniana Infection. J. Fungi 2024, 10, 258. https://doi.org/10.3390/jof10040258
Istomina EA, Korostyleva TV, Kovtun AS, Slezina MP, Odintsova TI. Transcriptome-Wide Identification and Expression Analysis of Genes Encoding Defense-Related Peptides of Filipendula ulmaria in Response to Bipolaris sorokiniana Infection. Journal of Fungi. 2024; 10(4):258. https://doi.org/10.3390/jof10040258
Chicago/Turabian StyleIstomina, Ekaterina A., Tatyana V. Korostyleva, Alexey S. Kovtun, Marina P. Slezina, and Tatyana I. Odintsova. 2024. "Transcriptome-Wide Identification and Expression Analysis of Genes Encoding Defense-Related Peptides of Filipendula ulmaria in Response to Bipolaris sorokiniana Infection" Journal of Fungi 10, no. 4: 258. https://doi.org/10.3390/jof10040258
APA StyleIstomina, E. A., Korostyleva, T. V., Kovtun, A. S., Slezina, M. P., & Odintsova, T. I. (2024). Transcriptome-Wide Identification and Expression Analysis of Genes Encoding Defense-Related Peptides of Filipendula ulmaria in Response to Bipolaris sorokiniana Infection. Journal of Fungi, 10(4), 258. https://doi.org/10.3390/jof10040258