
Identification of Chromoblastomycosis and Phaeohyphomycosis Agents through ITS-RFLP

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Strains
2.2. Morphological Identification
2.3. DNA Extraction
2.4. Molecular Identification through Sequencing
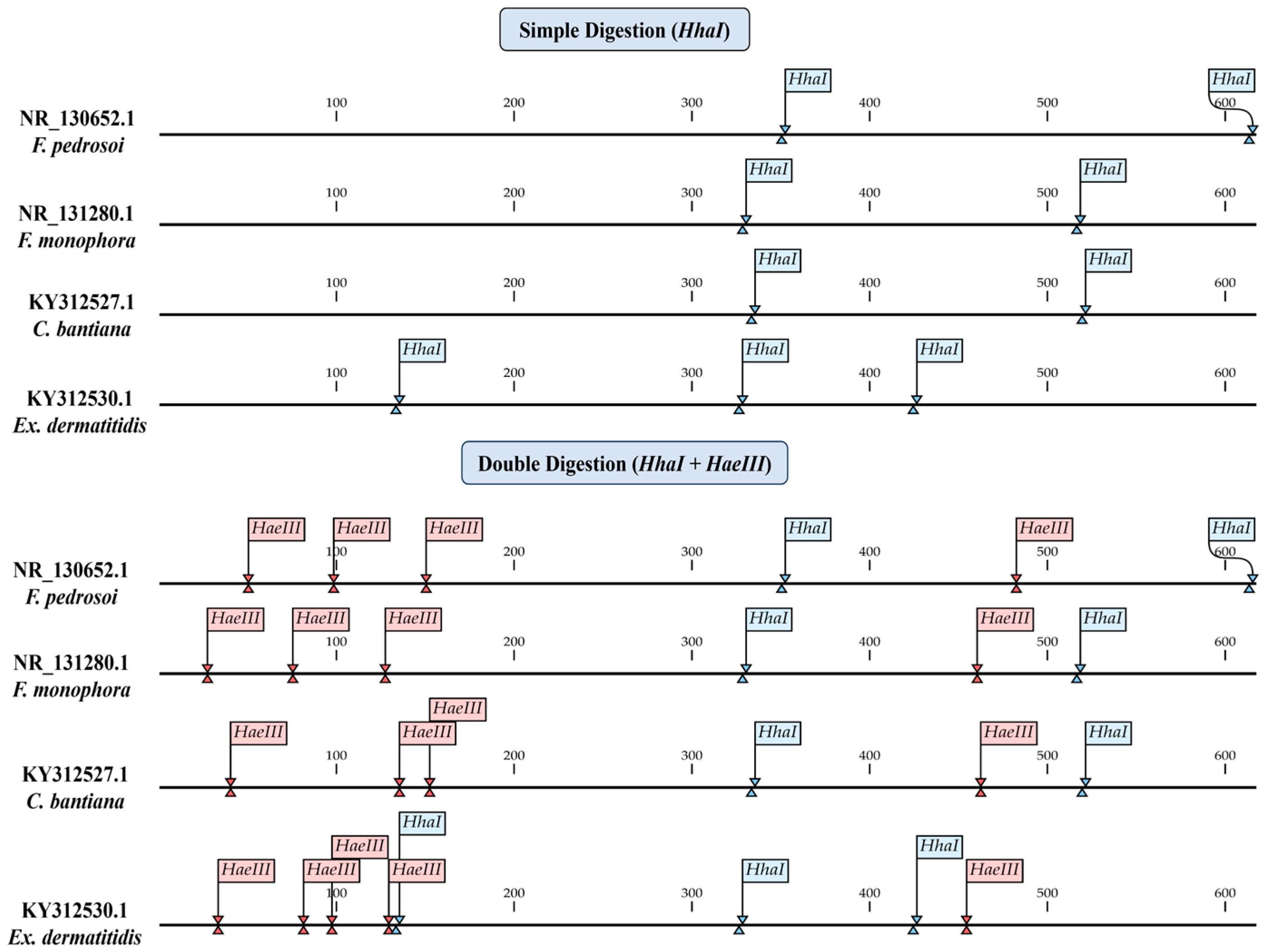
2.5. In Silico ITS-RFLP
2.6. ITS-RFLP
3. Results
3.1. Morphological Identification and Sequencing of the Isolates
3.2. In Silico Analyses
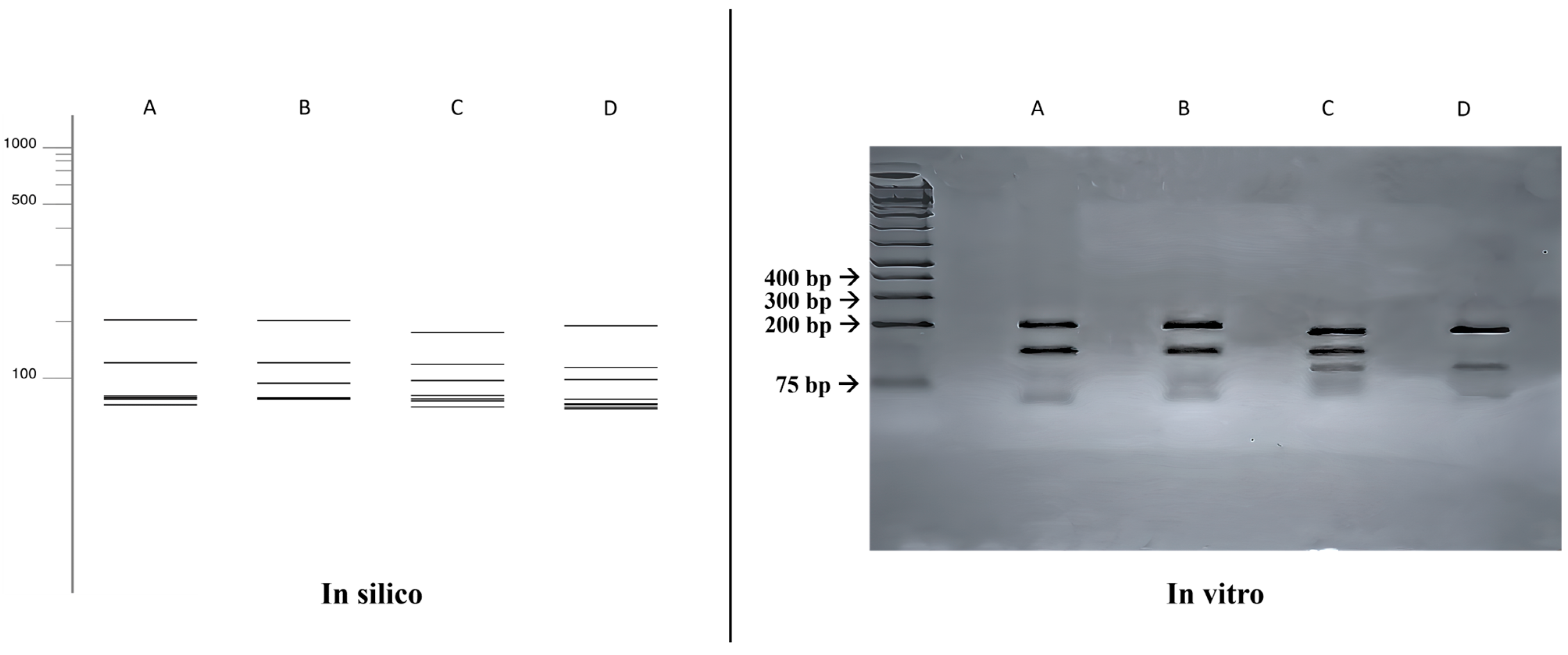
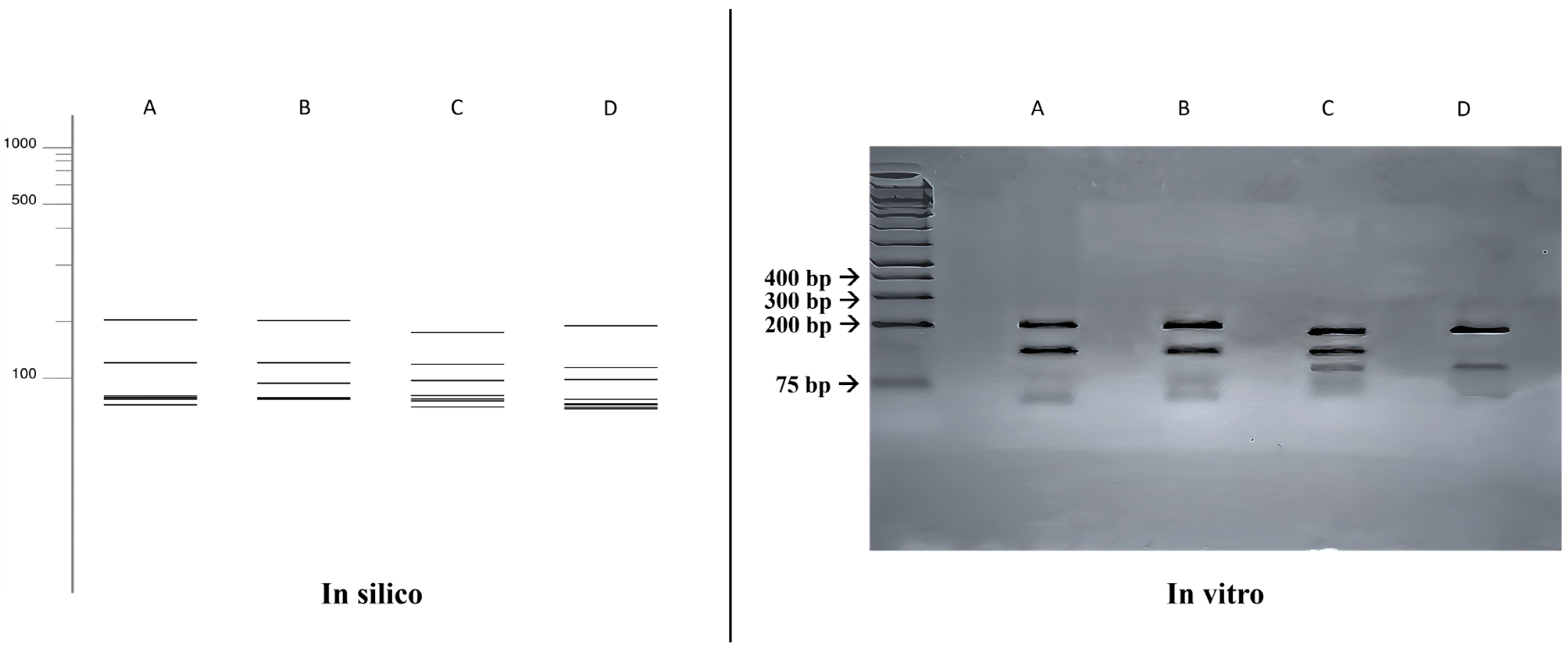
3.3. ITS-RFLP with HhaI
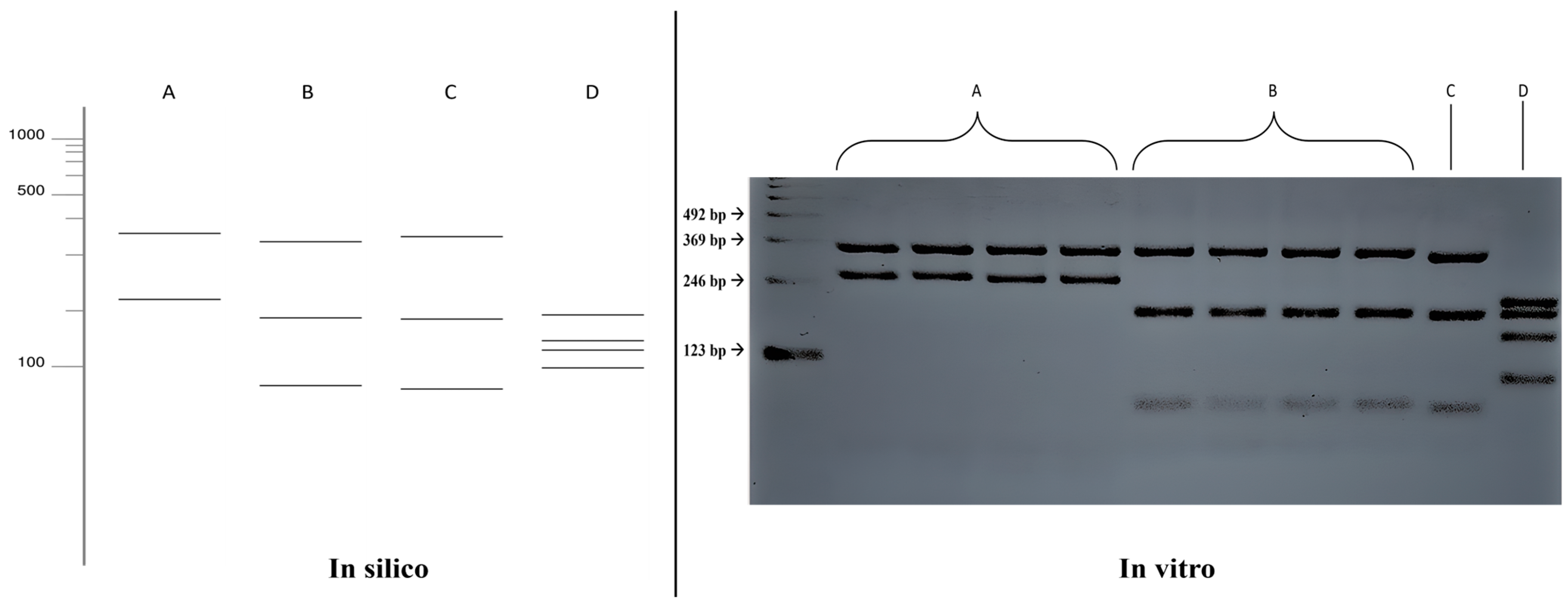
3.4. ITS-RFLP with HhaI and HaeIII
3.5. Comparative Analysis of Identification Methods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arcobello, J.T.; Revankar, S.G. Phaeohyphomycosis. Semin. Respir. Crit. Care Med. 2020, 41, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Queiroz-Telles, F.; de Hoog, S.; Santos, D.W.C.L.; Salgado, C.G.; Vicente, V.A.; Bonifaz, A.; Roilides, E.; Xi, L.; Azevedo, C.d.M.P.e.S.; da Silva, M.B.; et al. Chromoblastomycosis. Clin. Microbiol. Rev. 2017, 30, 233–276. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.W.C.L.; Azevedo, C.d.M.P.e.S.d.; Vicente, V.A.; Queiroz-Telles, F.; Rodrigues, A.M.; de Hoog, G.S.; Denning, D.W.; Colombo, A.L. The Global Burden of Chromoblastomycosis. PLoS Neglected Trop. Dis. 2021, 15, e0009611. [Google Scholar] [CrossRef] [PubMed]
- Enbiale, W.; Bekele, A.; Manaye, N.; Seife, F.; Kebede, Z.; Gebremeskel, F.; van Griensven, J. Subcutaneous Mycoses: Endemic but Neglected among the Neglected Tropical Diseases in Ethiopia. PLoS Neglected Trop. Dis. 2023, 17, e0011363. [Google Scholar] [CrossRef]
- WHO. Ending the Neglect to Attain the Sustainable Development Goals: A Strategic Framework for Integrated Control and Management of Skin-Related Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2022.
- Agarwal, R.; Singh, G.; Ghosh, A.; Verma, K.K.; Pandey, M.; Xess, I. Chromoblastomycosis in India: Review of 169 Cases. PLoS Neglected Trop. Dis. 2017, 11, e0005534. [Google Scholar] [CrossRef]
- Guevara, A.; Siqueira, N.P.; Nery, A.F.; Cavalcante, L.R.D.S.; Hagen, F.; Hahn, R.C. Chromoblastomycosis in Latin America and the Caribbean: Epidemiology over the Past 50 Years. Med. Mycol. 2021, 60, myab062. [Google Scholar] [CrossRef]
- He, Y.; Zheng, H.L.; Mei, H.; Lv, G.X.; Liu, W.D.; Li, X.F. Phaeohyphomycosis in China. Front. Cell. Infect. Microbiol. 2022, 12, 895329. [Google Scholar] [CrossRef]
- Thomas, E.; Bertolotti, A.; Barreau, A.; Klisnick, J.; Tournebize, P.; Borgherini, G.; Zemali, N.; Jaubert, J.; Jouvion, G.; Bretagne, S.; et al. From Phaeohyphomycosis to Disseminated Chromoblastomycosis: A Retrospective Study of Infections Caused by Dematiaceous Fungi. Med. Mal Infect. 2018, 48, 278–285. [Google Scholar] [CrossRef]
- Liu, S.; Zhi, H.; Shen, H.; Lv, W.; Sang, B.; Li, Q.; Zhong, Y.; Liu, Z.; Xia, X. Chromoblastomycosis: A Case Series from Eastern China. PLoS Neglected Trop. Dis. 2022, 16, e0010800. [Google Scholar] [CrossRef]
- Passero, L.F.D.; Cavallone, I.N.; Belda, W. Reviewing the Etiologic Agents, Microbe-Host Relationship, Immune Response, Diagnosis, and Treatment in Chromoblastomycosis. J. Immunol. Res. 2021, 2021, 1–23. [Google Scholar] [CrossRef]
- de Brito, A.C.; Bittencourt, M.d.J.S. Chromoblastomycosis: An Etiological, Epidemiological, Clinical, Diagnostic, and Treatment Update. An. Bras. Dermatol. 2018, 93, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Revankar, S.G.; Sutton, D.A. Melanized Fungi in Human Disease. Clin. Microbiol. Rev. 2010, 23, 884–928. [Google Scholar] [CrossRef] [PubMed]
- Carolina Rojas, O.; León-Cachón, R.B.R.; Pérez-Maya, A.A.; Aguirre-Garza, M.; Moreno-Treviño, M.G.; González, G.M. Phenotypic and Molecular Identification of Fonsecaea Pedrosoi Strains Isolated from Chromoblastomycosis Patients in Mexico and Venezuela. Mycoses 2015, 58, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Bienvenu, A.L.; Picot, S. Mycetoma and Chromoblastomycosis: Perspective for Diagnosis Improvement Using Biomarkers. Molecules 2020, 25, 2594. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Figueras, M.J. Atlas of Clinical Fungi, 2nd ed.; Amer Society for Microbiology: Utrech, The Netherlands; Universitat Rovira i Virgili Reus: Reus, Spain, 2000; pp. 560–680. [Google Scholar]
- Monteiro, R.C. Epidemiologia Molecular e Caracterização Fenotípica de Fungos Demáceos Agentes de Cromoblastomicose no Estado do Pará, com Relato de Fungos Neurotrópicos na Região Amazônica; Dissertação de Mestrado, Universidade Federal do Pará: Belém, PA, Brasil, 2017. [Google Scholar]
- Sun, J.; Najafzadeh, M.J.; Gerrits van den Ende, A.H.G.; Vicente, V.A.; Feng, P.; Xi, L.; de Hoog, G.S. Molecular Characterization of Pathogenic Members of the Genus Fonsecaea Using Multilocus Analysis. PLoS ONE 2012, 7, e41512. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2018, 20, 1160–1166. [Google Scholar] [CrossRef]
- Roberto, T.N.; Rodrigues, A.M.; Hahn, R.C.; De Camargo, Z.P. Identifying Paracoccidioides Phylogenetic Species by PCR-RFLP of the Alpha-Tubulin Gene. Med. Mycol. 2016, 54, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Vincze, T.; Posfai, J.; Roberts, R.J. NEBcutter: A Program to Cleave DNA with Restriction Enzymes. Nucleic Acids Res. 2003, 31, 3688–3691. [Google Scholar] [CrossRef]
- Borman, A.M.; Fraser, M.; Patterson, Z.; Linton, C.J.; Palmer, M.; Johnson, E.M. Fungal Infections of Implantation: More Than Five Years of Cases of Subcutaneous Fungal Infections Seen at the UK Mycology Reference Laboratory. J. Fungi 2022, 8, 343. [Google Scholar] [CrossRef]
- Sendrasoa, F.A.; Razanakoto, N.H.; Rakotoarisaona, M.F.; Andrianarison, M.; Raharolahy, O.; Rasamoelina, T.; Ranaivo, I.M.; Sata, M.; Ratovonjanahary, V.; Maubon, D.; et al. Clinical Aspects of Previously Treated Chromoblastomycosis: A Case Series from Madagascar. Int. J. Infect. Dis. 2020, 101, 228–232. [Google Scholar] [CrossRef]
- Ariani, T.; Rizal, Y.; Ledika Veroci, R. Clinical and Mycological Spectrum of Chromoblastomycosis; 2023; Volume 33, Available online: https://www.jpad.com.pk/index.php/jpad/article/view/2399 (accessed on 18 November 2023).
- Revankar, S.G. Phaeohyphomycosis in Transplant Patients. Fungi 2015, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Xu, J.; Su, Q.; Chen, Y. Exophiala Dermatitis and Exacerbation of Chronic Obstructive Pulmonary Disease. QJM 2019, 112, 869–871. [Google Scholar] [CrossRef]
- An, L.; Jia, G.; Tan, J.; Yang, L.; Wang, Y.; Li, L. Analysis of the Synergistic Antifungal Activity of Everolimus and Antifungal Drugs against Dematiaceous Fungi. Front. Cell. Infect. Microbiol. 2023, 13, 1131416. [Google Scholar] [CrossRef] [PubMed]
- Coelho, R.A.; Brito-Santos, F.; Figueiredo-Carvalho, M.H.G.; Silva, J.V.d.S.; Gutierrez-Galhardo, M.C.; do Valle, A.C.F.; Zancopé-Oliveira, R.M.; Trilles, L.; Meyer, W.; Freitas, D.F.S.; et al. Molecular Identification and Antifungal Susceptibility Profiles of Clinical Strains of Fonsecaea Spp. Isolated from Patients with Chromoblastomycosis in Rio de Janeiro, Brazil. PLoS Neglected Trop. Dis. 2018, 12, e0006675. [Google Scholar] [CrossRef]
- Nascimento, E.; Barião, P.H.G.; Kress, M.R.v.Z.; Vilar, F.C.; Santana, R.d.C.; Gaspar, G.G.; Martinez, R. Cryptococcosis by Cryptococcus Neoformans/Cryptococcus Gattii Species Complexes in Non-Hiv-Infected Patients in Southeastern Brazil. Rev. Soc. Bras. Med. Trop. 2021, 54, E0169. [Google Scholar] [CrossRef]
- da Fonseca, L.A.V.; Araújo, M.A.S.; Silva, D.M.W.; de Albuquerque Maranhão, F.C. ITS-RFLP Optimization for Dermatophyte Identification from Clinical Sources in Alagoas (Brazil) versus Phenotypic Methods. J. Infect. Dev. Ctries 2022, 16, 1773–1777. [Google Scholar] [CrossRef] [PubMed]
- Fontecha, G.; Montes, K.; Ortiz, B.; Galindo, C.; Braham, S. Identification of Cryptic Species of Four Candida Complexes in a Culture Collection. J. Fungi 2019, 5, 117. [Google Scholar] [CrossRef]
- Maubon, D.; Garnaud, C.; Ramarozatovo, L.S.; Fahafahantsoa, R.R.; Cornet, M.; Rasamoelina, T. Molecular Diagnosis of Two Major Implantation Mycoses: Chromoblastomycosis and Sporotrichosis. J. Fungi 2022, 8, 382. [Google Scholar] [CrossRef]
- Fajarningsih, N.D. Internal Transcribed Spacer (ITS) as Dna Barcoding to Identify Fungal Species: A Review. Squalen Bull. Mar. Fish. Postharvest Biotechnol. 2016, 11, 37–44. [Google Scholar] [CrossRef]
- Hoang, M.T.V.; Irinyi, L.; Chen, S.C.A.; Sorrell, T.C.; Meyer, W. Dual DNA Barcoding for the Molecular Identification of the Agents of Invasive Fungal Infections. Front. Microbiol. 2019, 10, 1647. [Google Scholar] [CrossRef]
- Xu, J. Fungal DNA Barcoding1. Genome; Canadian Science Publishing: Ottawa, ON, Canada, 2016; pp. 913–932. [Google Scholar] [CrossRef]
- Réblová, M.; Untereiner, W.A.; Réblová, K. Novel Evolutionary Lineages Revealed in the Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of ITS Secondary Structure. PLoS ONE 2013, 8, e63547. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Moreno, L.F.; Stielow, B.J.; Muszewska, A.; Hainaut, M.; Gonzaga, L.; Abouelleil, A.; Patané, J.S.L.; Priest, M.; Souza, R.; et al. Exploring the Genomic Diversity of Black Yeasts and Relatives (Chaetothyriales, Ascomycota). Stud. Mycol. 2017, 86, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Najafzadeh, M.J.; Gueidan, C.; Badali, H.; Van Den Ende, A.H.G.; Xi, L.; Hoog, G.S. Genetic Diversity and Species Delimitation in the Opportunistic Genus Fonsecaea. Med. Mycol. 2009, 47, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Usuda, D.; Higashikawa, T.; Hotchi, Y.; Usami, K.; Shimozawa, S.; Tokunaga, S.; Osugi, I.; Katou, R.; Ito, S.; Yoshizawa, T.; et al. Exophiala Dermatitidis. World J. Clin. Cases 2021, 9, 7963–7972. [Google Scholar] [CrossRef] [PubMed]
- Castillo Bejarano, J.I.; de los Santos, A.M.; Saldaña, D.C.; Zambrano Lucio, M.; Pérez Cavazos, S.; Espinosa Villaseñor, F.; de la O Cavazos, M.E.; Vaquera Aparicio, D.N. Pediatric Phaeohyphomycosis: A 44-Year Systematic Review of Reported Cases. J. Pediatr. Infect. Dis. Soc. 2022, 12, 10–20. [Google Scholar] [CrossRef]
- Le Govic, Y.; Demey, B.; Cassereau, J.; Bahn, Y.S.; Papon, N. Pathogens Infecting the Central Nervous System. PLoS Pathog. 2022, 18, e1010234. [Google Scholar] [CrossRef]
- Ganesan, V.; Hallur, V.; Velvizhi, S.; Rajendran, T. Cerebral Phaeohyphomycosis Due to Cladophialophora Bantiana: Case Report and Systematic Review of Cases. Infection 2023. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Genbank | Species | Anatomical Site | Host | Geographic Origin |
|---|---|---|---|---|---|
| IEC-CBM02 | KY312523 | F. pedrosoi | Thigh | Human | Pará/Brazil |
| IEC-CBM03 | KY312524 | F. pedrosoi | Foot | Human | Pará/Brazil |
| IEC-CBM04 | KY312525 | F. pedrosoi | Leg | Human | Pará/Brazil |
| IEC-CBM05 | KY312526 | F. pedrosoi | Leg | Human | Pará/Brazil |
| IEC-CBM06 | KY312527 | C. bantiana | CNS † | Human | Pará/Brazil |
| IEC-CBM07 | KY312528 | F. pedrosoi | Hand | Human | Pará/Brazil |
| IEC-CBM08 | KY312529 | F. pedrosoi | Leg | Human | Pará/Brazil |
| IEC-CBM09 | KY312530 | E. dermatitidis | * | Human | Pará/Brazil |
| IEC-CBM10 | KY312531 | F. pedrosoi | Arm | Human | Pará/Brazil |
| IEC-CBM11 | KY312532 | F. pedrosoi | Leg | Human | Pará/Brazil |
| IEC-CBM12 | KY312533 | F. pedrosoi | Foot | Human | Pará/Brazil |
| IEC-CBM13 | KY312534 | F. monophora | Leg | Human | Pará/Brazil |
| IEC-CBM14 | KY312535 | F. pedrosoi | * | Human | Pará/Brazil |
| IEC-CBM15 | KY312536 | F. pedrosoi | * | Human | Pará/Brazil |
| IEC-CBM16 | KY312537 | F. pedrosoi | Thigh | Human | Pará/Brazil |
| IEC-CBM17 | KY312538 | F. monophora | Leg | Human | Pará/Brazil |
| IEC-CBM18 | MF416919 | F. pedrosoi | Thigh | Human | Pará/Brazil |
| IEC-CBM19 | MF416920 | F. monophora | Forearm | Human | Pará/Brazil |
| IEC-CBM21 | MF416922 | F. pedrosoi | Fist | Human | Pará/Brazil |
| IEC-CBM22 | MF416923 | F. pedrosoi | Ankle | Human | Pará/Brazil |
| IEC-CBM23 | MF416924 | F. monophora | * | Human | Pará/Brazil |
| Sample | Morphological Identification | ITS-RFLP (HhaI) | ITS-RFLP (HhaI + HaeIII) | Sequencing Identification |
|---|---|---|---|---|
| IEC-CBM02 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM03 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM04 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM05 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM06 | Cladophialophora sp. | C. bantiana/ F. monophora * | C. bantiana | C. bantiana |
| IEC-CBM07 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM08 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM09 | Exophiala sp. | E. dermatitidis | E. dermatitidis | E. dermatitidis |
| IEC-CBM10 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM11 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM12 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM13 | Fonsecaea sp. | F. monophora | F. monophora | F. monophora |
| IEC-CBM14 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM15 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM16 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM17 | Fonsecaea sp. | F. monophora | F. monophora | F. monophora |
| IEC-CBM18 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM19 | Fonsecaea sp. | F. monophora | F. monophora | F. monophora |
| IEC-CBM21 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM22 | Fonsecaea sp. | F. pedrosoi | F. pedrosoi | F. pedrosoi |
| IEC-CBM23 | Fonsecaea sp. | F. monophora | F. monophora | F. monophora |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, G.S.M.; De Oliveira, R.S.; De Souza, A.B.; Monteiro, R.C.; Santo, E.P.T.E.; Franco Filho, L.C.; Da Silva, S.H.M. Identification of Chromoblastomycosis and Phaeohyphomycosis Agents through ITS-RFLP. J. Fungi 2024, 10, 159. https://doi.org/10.3390/jof10020159
Sousa GSM, De Oliveira RS, De Souza AB, Monteiro RC, Santo EPTE, Franco Filho LC, Da Silva SHM. Identification of Chromoblastomycosis and Phaeohyphomycosis Agents through ITS-RFLP. Journal of Fungi. 2024; 10(2):159. https://doi.org/10.3390/jof10020159
Chicago/Turabian StyleSousa, Gabriel S. M., Rodrigo S. De Oliveira, Alex B. De Souza, Ruan C. Monteiro, Elaine P. T. E. Santo, Luciano C. Franco Filho, and Silvia H. M. Da Silva. 2024. "Identification of Chromoblastomycosis and Phaeohyphomycosis Agents through ITS-RFLP" Journal of Fungi 10, no. 2: 159. https://doi.org/10.3390/jof10020159
APA StyleSousa, G. S. M., De Oliveira, R. S., De Souza, A. B., Monteiro, R. C., Santo, E. P. T. E., Franco Filho, L. C., & Da Silva, S. H. M. (2024). Identification of Chromoblastomycosis and Phaeohyphomycosis Agents through ITS-RFLP. Journal of Fungi, 10(2), 159. https://doi.org/10.3390/jof10020159