
Bioprospection of Tenellins Produced by the Entomopathogenic Fungus Beauveria neobassiana

 , ,
, ,  and
and 
Abstract
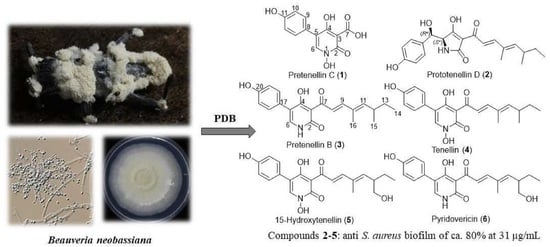
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Fungal Material
2.3. Cultivation, Extraction and Isolation
2.4. Spectral Data
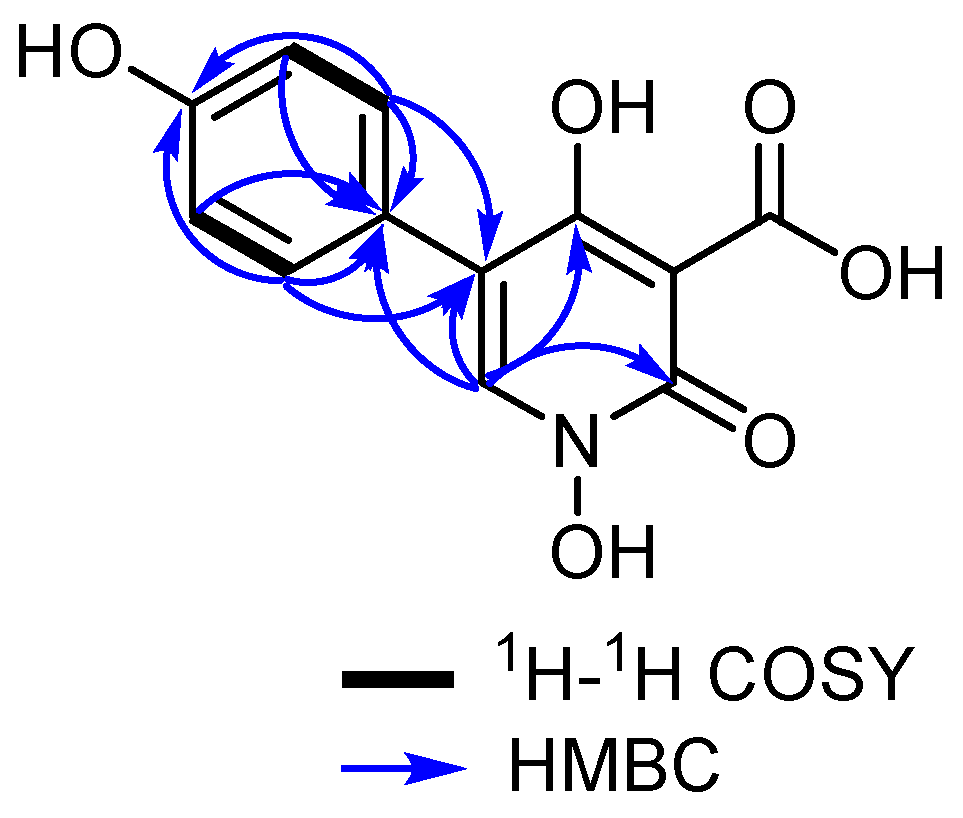
2.4.1. Pretenellin C (1)
2.4.2. Prototenellin D (2)
2.4.3. Pretenellin B (3)
2.4.4. Tenellin (4)
2.4.5. 15-Hydroxytenellin (5)
2.4.6. Pyridovericin (6)
2.5. Biological Assays
3. Results
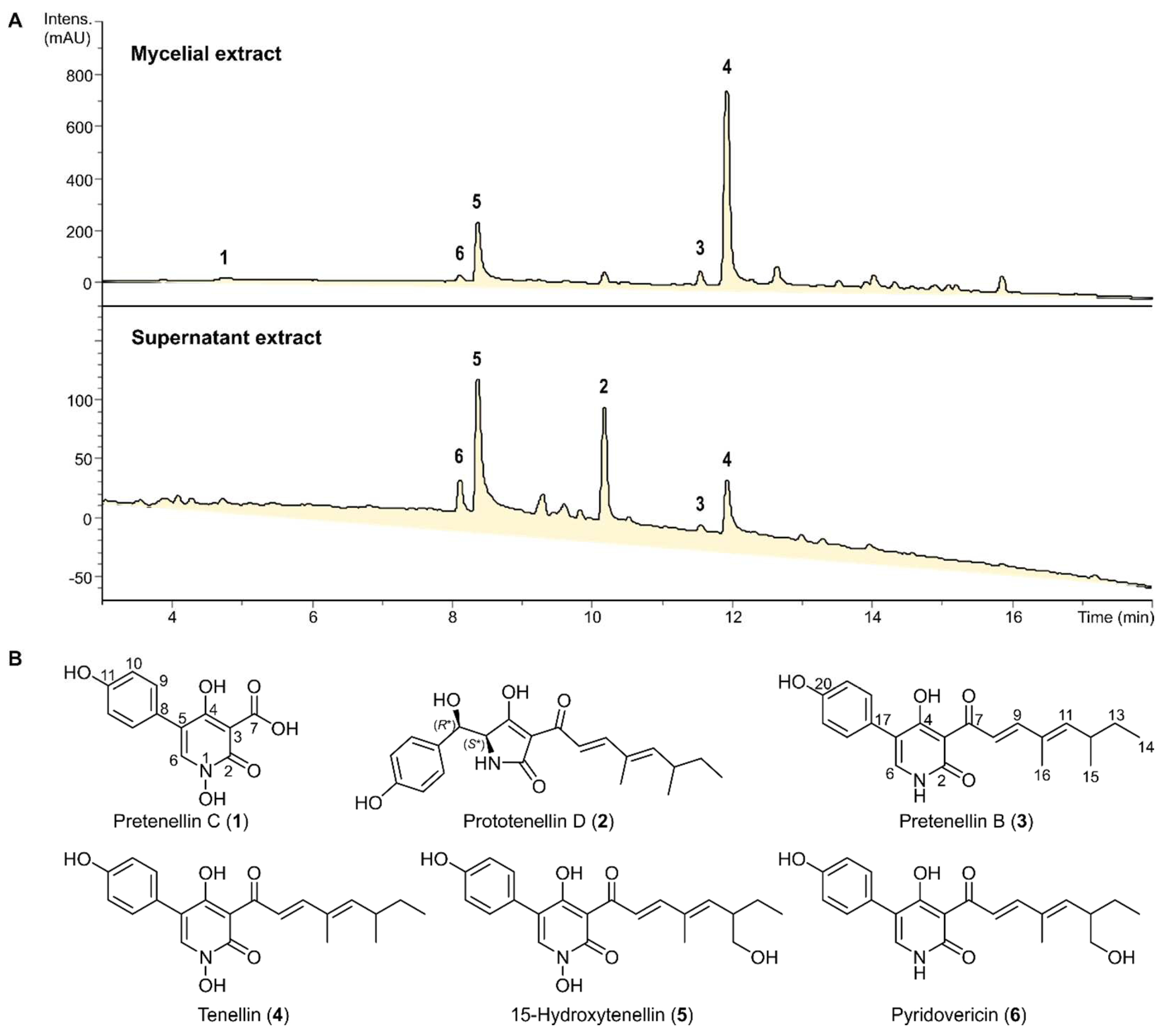
3.1. Isolation and Structure Elucidation of Secondary Metabolites

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pos. | δH a (multi, J [Hz]) | δC, b Type |
|---|---|---|
| 2 | 161.2, CO | |
| 3 | 98.2, C | |
| 4 | 170.5, CO | |
| 5 | 123.6, C | |
| 6 | 8.18 (s, 1H) | 138.7, CH |
| 7 | 13.94 (br) | n.d. c |
| 8 | 115.3, C | |
| 9,13 | 7.42 (d, 8.3, 2H) | 131.2, CH |
| 10,12 | 6.92 (d, 8.3, 2H) | 115.9, CH |
| 11 | 158.4, C |
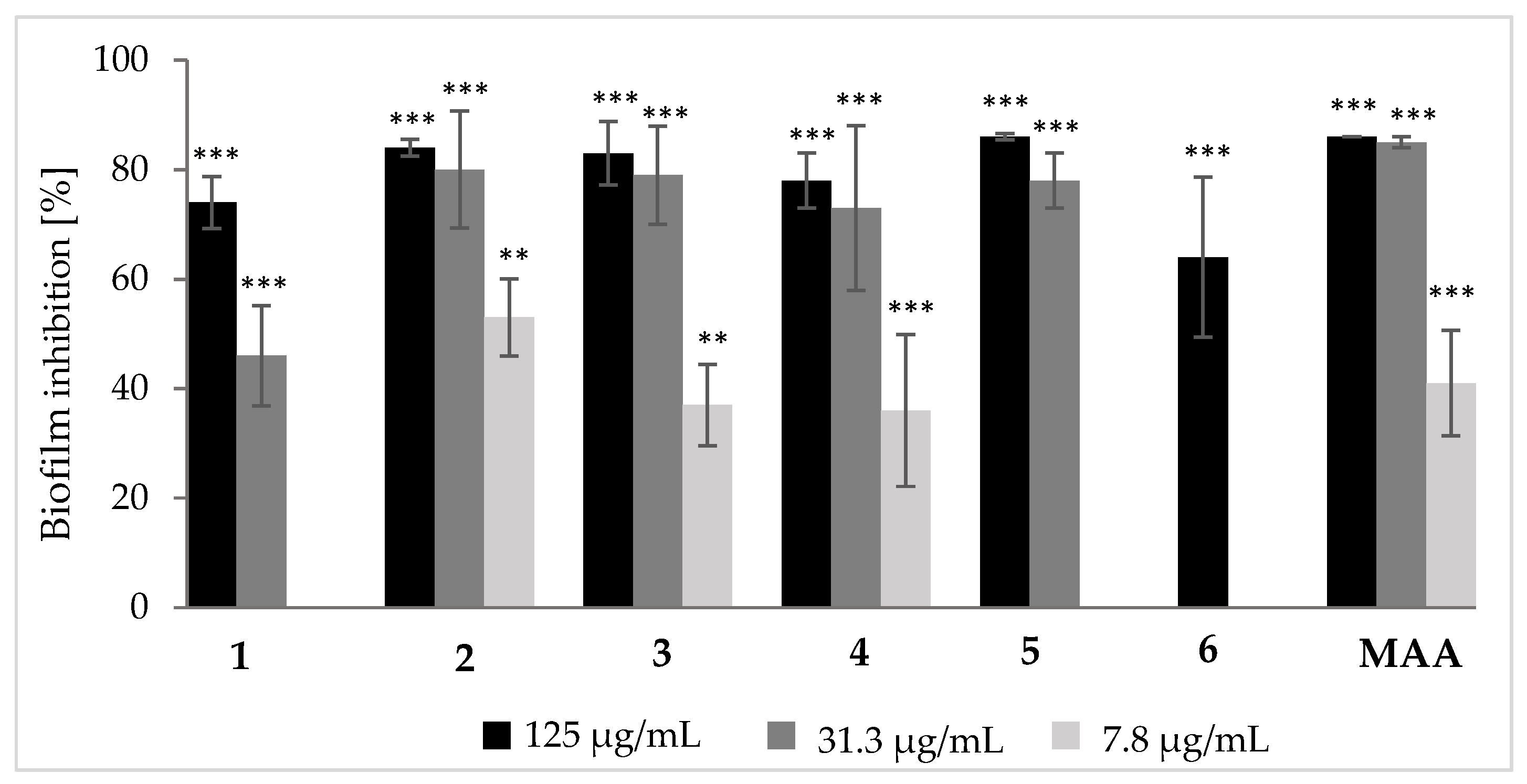
3.2. Biological Effects of Tenellin-Derived Metabolites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mapook, A.; Hyde, K.D.; Hassan, K.; Kemkuignou, B.M.; Čmoková, A.; Surup, F.; Kuhnert, E.; Paomephan, P.; Cheng, T.; de Hoog, S.; et al. Ten decadal advances in fungal biology leading towards human well-being. Fungal Divers. 2022, 116, 547–614. [Google Scholar] [CrossRef]
- Niego, G.A.T.; Lambert, C.; Mortimer, P.; Thongklang, N.; Rapior, S.; Grosse, M.; Schrey, H.; Charria-Girón, E.; Hyde, K.D.; Stadler, M. The contribution of fungi to the global economy. Fungal Divers. 2023, 121, 95–137. [Google Scholar] [CrossRef]
- Mongkolsamrit, S.; Khonsanit, A.; Thanakitpipattana, D.; Tasanathai, K.; Noisripoom, W.; Lamlertthon, S.; Himaman, W.; Houbraken, J.; Samson, R.A.; Luangsa-ard, J. Revisiting Metarhizium and the description of new species from Thailand. Stud. Mycol. 2020, 95, 171–251. [Google Scholar] [CrossRef] [PubMed]
- Kobmoo, N.; Arnamnart, N.; Pootakham, W.; Sonthirod, C.; Khonsanit, A.; Kuephadungphan, W.; Suntivich, R.; Mosunova, O.V.; Giraud, T.; Luangsa-ard, J.J. The integrative taxonomy of Beauveria asiatica and B. bassiana species complexes with whole-genome sequencing, morphometric and chemical analyses. Persoonia 2021, 47, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Du, F.Y.; Li, X.M.; Sun, Z.C.; Meng, L.H.; Wang, B.G. Secondary metabolites with agricultural antagonistic potentials from Beauveria felina, a marine-derived entomopathogenic fungus. J. Agric. Food Chem. 2020, 68, 14824–14831. [Google Scholar] [CrossRef]
- Buchanan, G.O.; Williams, L.A.; Reese, P.B. Biotransformation of cadinane sesquiterpenes by Beauveria bassiana ATCC 7159. Phytochemistry 2000, 54, 39–45. [Google Scholar] [CrossRef]
- Wang, H.; Peng, H.; Li, W.; Cheng, P.; Gong, M. The toxins of Beauveria bassiana and the strategies to improve their virulence to insects. Front. Microbiol. 2021, 12, 705343. [Google Scholar] [CrossRef] [PubMed]
- Halo, L.M.; Heneghan, M.N.; Yakasai, A.A.; Song, Z.; Williams, K.; Bailey, A.M.; Cox, R.J.; Lazarus, C.M.; Simpson, T.J. Late stage oxidations during the biosynthesis of the 2-pyridone tenellin in the entomopathogenic fungus Beauveria bassiana. J. Am. Chem. Soc. 2008, 130, 17988–17996. [Google Scholar] [CrossRef]
- Mirghani, R.; Saba, T.; Khaliq, H.; Mitchell, J.; Do, L.; Chambi, L.; Diaz, K.; Kennedy, T.; Alkassab, K.; Huynh, T.; et al. Biofilms: Formation, drug resistance and alternatives to conventional approaches. AIMS Microbiol. 2022, 8, 239–277. [Google Scholar] [CrossRef]
- Takahashi, S.; Kakinuma, N.; Uchida, K.; Hashimoto, R.; Yanagisawa, T.; Nakagawa, A. Pyridovericin and pyridomacrolidin: Novel metabolites from entomopathogenic fungi Beauveria bassiana. J. Antibiot. 1998, 51, 596–598. [Google Scholar] [CrossRef]
- Becker, K.; Wessel, A.-C.; Luangsa-ard, J.J.; Stadler, M. Viridistratins A-C, antimicrobial and cytotoxic benzo[j]fluoranthenes from stomata of Annulohypoxylon viridistatum (Hypoxylaceae, Ascomycota). Biomolecules 2020, 10, 805. [Google Scholar] [CrossRef] [PubMed]
- Charria-Girón, E.; Stchigel, A.M.; Čmoková, A.; Kolařík, M.; Surup, F.; Marin-Felix, Y. Amesia hispanica sp. nov., producer of the antifungal class of antibiotics dactylfungins. J. Fungi 2023, 9, 463. [Google Scholar] [CrossRef] [PubMed]
- Soliga, K.J.; Bär, S.I.; Oberhuber, N.; Zeng, H.; Schrey, H.; Schobert, R. Synthesis and bioactivity of ancorinoside B, a marine diglycosyl tetramic acid. Mar. Drugs 2021, 19, 583. [Google Scholar] [CrossRef] [PubMed]
- Chepkirui, C.; Yuyama, K.T.; Wanga, L.A.; Decock, C.; Matasyoh, J.C.; Abraham, W.-R.; Stadler, M. Microporenic acids A-G, biofilm inhibitors, and antimicrobial agents from the basidiomycete Microporus Species. J. Nat. Prod. 2018, 81, 778–784. [Google Scholar] [CrossRef]
- Gerth, K.; Bedorf, N.; Höfle, G.; Irschik, H.; Reichenbach, H. Epothilons A and B: Antifungal and cytotoxic compounds from Sorangium cellulosum (Myxobacteria): Production, physico-chemical and biological properties. J. Antibiot. 1996, 49, 560–563. [Google Scholar] [CrossRef]
- Goodin, S.; Kane, M.P.; Rubin, E.H. Epothilones: Mechanism of action and biologic activity. J. Clin. Oncol. 2004, 22, 2015–2025. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, D.; Wu, D.; He, W.; Zuo, M.; Zhu, W.; Xu, Y.; Wang, L. Two New 4-Hydroxy-2-pyridone Alkaloids with antimicrobial and cytotoxic activities from Arthrinium sp. GZWMJZ-606 endophytic with Houttuynia cordata Thunb. Molecules 2023, 28, 2192. [Google Scholar] [CrossRef]
- Bergmann, S.; Schümann, J.; Scherlach, K.; Lange, C.; Brakhage, A.A. Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulans. Nat. Chem. Biol. 2007, 3, 213–217. [Google Scholar] [CrossRef]
- Rizwan-ul-Haq, M.; Hu, Q.B.; Hu, M.Y.; Lin, Q.S.; Zhang, W.L. Biological impact of harmaline, ricinine and their combined effects with Bacillus thuringiensis on Spodoptera exigua (Lepidoptera: Noctuidae). J. Pest Sci. 2009, 82, 327–334. [Google Scholar] [CrossRef]
- Schmidt, K.; Riese, U.; Li, Z.; Hamburger, M. Novel tetramic acids and pyridone alkaloids, militarinones B, C, and D, from the insect pathogenic fungus Paecilomyces militaris. J. Nat. Prod. 2003, 66, 378–383. [Google Scholar] [CrossRef]
- Kornsakulkarn, J.; Pruksatrakul, T.; Surawatanawong, P.; Thangsrikeattigun, C.; Komwijit, S.; Boonyuen, N.; Thongpanchang, C. Antimicrobial, antimalarial, and cytotoxic substances from the insect pathogenic fungus Beauveria asiatica BCC 16812. Phytochem. Lett. 2021, 43, 8–15. [Google Scholar] [CrossRef]
- Colarusso, S.; Narjes, F. Preparation of Pyridine N-oxides as Antiviral Agents. WO2004110442 A1, 23 December 2004. [Google Scholar]
| IC50 (µM) | Positive Control | ||||||
|---|---|---|---|---|---|---|---|
| Test Cell Line | 1 | 2 | 3 | 4 | 5 | 6 | Epothilone B (nM) |
| L929 (fibroblast) | n.a. | 70.1 | 48.2 | 0.79 | 6.8 | 5.7 | 0.65 |
| KB3.1 (ovary) | n.a. | 64.7 | 20.2 | 0.79 | 6.0 | 4.9 | 0.17 |
| A549 (lung) | n.d. | n.d. | n.d. | 0.24 | 2.6 | 24.1 | 0.05 |
| MCF-7 (breast) | n.d. | n.d. | n.d. | 2.0 | 8.1 | 7.3 | 0.07 |
| Test Microorganism | MIC (µg/mL) | Positive Control (µg/mL) | |||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| Staphylococcus aureus | n.i. | n.i. | 66.6 | 16.6 | 66.6 | 66.6 | 0.21 G |
| Escherichia coli | n.i. | n.i. | n.i. | n.i. | n.i. | n.i. | 0.42 G |
| Bacillus subtilis | n.i. | n.i. | 66.6 | 8.3 | 66.6 | 66.6 | 16.6 O |
| Pseudomonas aeruginosa | n.i. | n.i. | n.i. | n.i. | n.i. | n.i. | 0.21 G |
| Wickerhamomyces anomalus | n.d. | n.i. | n.d. | n.i. | n.i. | n.d. | 16.6 N |
| Candida albicans | n.i. | n.i. | n.i. | 66.6 | n.i. | n.i. | 8.3 N |
| Acinetobacter baumannii | n.d. | n.i. | n.d. | n.i. | n.i. | n.d. | 0.52 C |
| Chromobacterium violaceum | n.d. | n.i. | n.i. | n.i. | n.i. | n.d. | 1.70 G |
| Schizosaccharomyces pombe | n.d. | n.i. | n.d. | n.i. | n.i. | n.d. | 4.20 N |
| Mucor hiemalis | n.i. | n.i. | n.i. | 66.6 | n.i. | n.i. | 8.30 N |
| Rhodotorula glutinis | n.d. | n.i. | n.d. | n.i. | n.i. | n.d. | 4.20 N |
| Mycobacterium smegmatis | n.i. | n.i. | n.i. | n.i. | n.i. | n.i. | 1.70 K |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toshe, R.; Charria-Girón, E.; Khonsanit, A.; Luangsa-ard, J.J.; Khalid, S.J.; Schrey, H.; Ebada, S.S.; Stadler, M. Bioprospection of Tenellins Produced by the Entomopathogenic Fungus Beauveria neobassiana. J. Fungi 2024, 10, 69. https://doi.org/10.3390/jof10010069
Toshe R, Charria-Girón E, Khonsanit A, Luangsa-ard JJ, Khalid SJ, Schrey H, Ebada SS, Stadler M. Bioprospection of Tenellins Produced by the Entomopathogenic Fungus Beauveria neobassiana. Journal of Fungi. 2024; 10(1):69. https://doi.org/10.3390/jof10010069
Chicago/Turabian StyleToshe, Rita, Esteban Charria-Girón, Artit Khonsanit, Janet Jennifer Luangsa-ard, Syeda Javariya Khalid, Hedda Schrey, Sherif S. Ebada, and Marc Stadler. 2024. "Bioprospection of Tenellins Produced by the Entomopathogenic Fungus Beauveria neobassiana" Journal of Fungi 10, no. 1: 69. https://doi.org/10.3390/jof10010069
APA StyleToshe, R., Charria-Girón, E., Khonsanit, A., Luangsa-ard, J. J., Khalid, S. J., Schrey, H., Ebada, S. S., & Stadler, M. (2024). Bioprospection of Tenellins Produced by the Entomopathogenic Fungus Beauveria neobassiana. Journal of Fungi, 10(1), 69. https://doi.org/10.3390/jof10010069