
Parasitism of Hirsutella rhossiliensis on Different Nematodes and Its Endophytism Promoting Plant Growth and Resistance against Root-Knot Nematodes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Microbial Strains and Culture Conditions
2.2. Nematodes
2.3. Arabidopsis thaliana
2.4. Genetic Transformation of H. rhossiliensis with Green and Red Fluorescent Proteins
2.5. Observation of the Fungal Infection Process on Nematodes
2.6. Parasitism Assay of H. rhossiliensis HR02 on Nematodes
2.7. Observation of H. rhossiliensis Colonizing Arabidopsis Roots
2.8. Measurement of Fungal Effects on A. thaliana Growth and Resistance to Root-Knot Nematodes
2.9. Data Analysis
3. Results
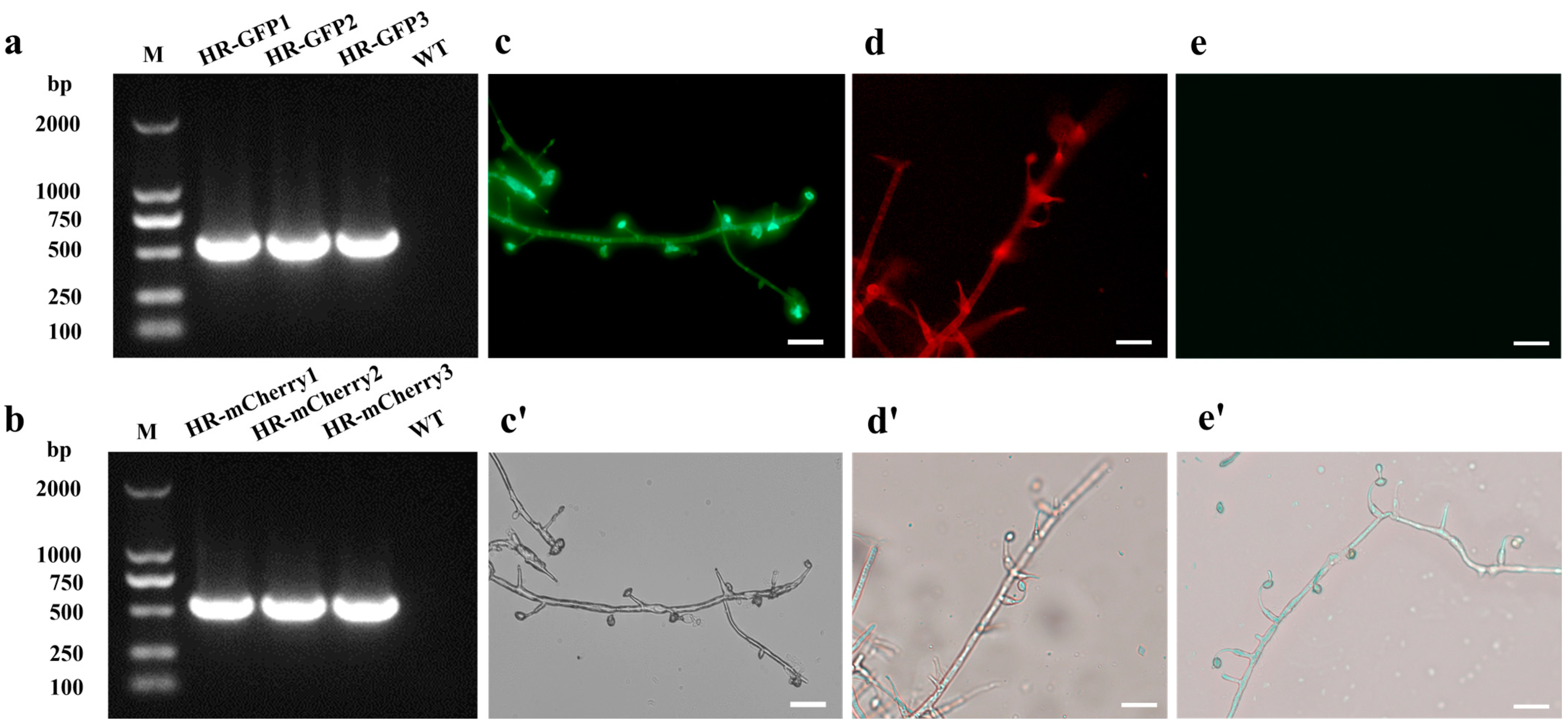
3.1. Acquisition of GFP-Tagged and mCherry-Tagged Transformants of H. rhossiliensis HR02
3.2. Observation of the Fungal Infection Process on Nematodes
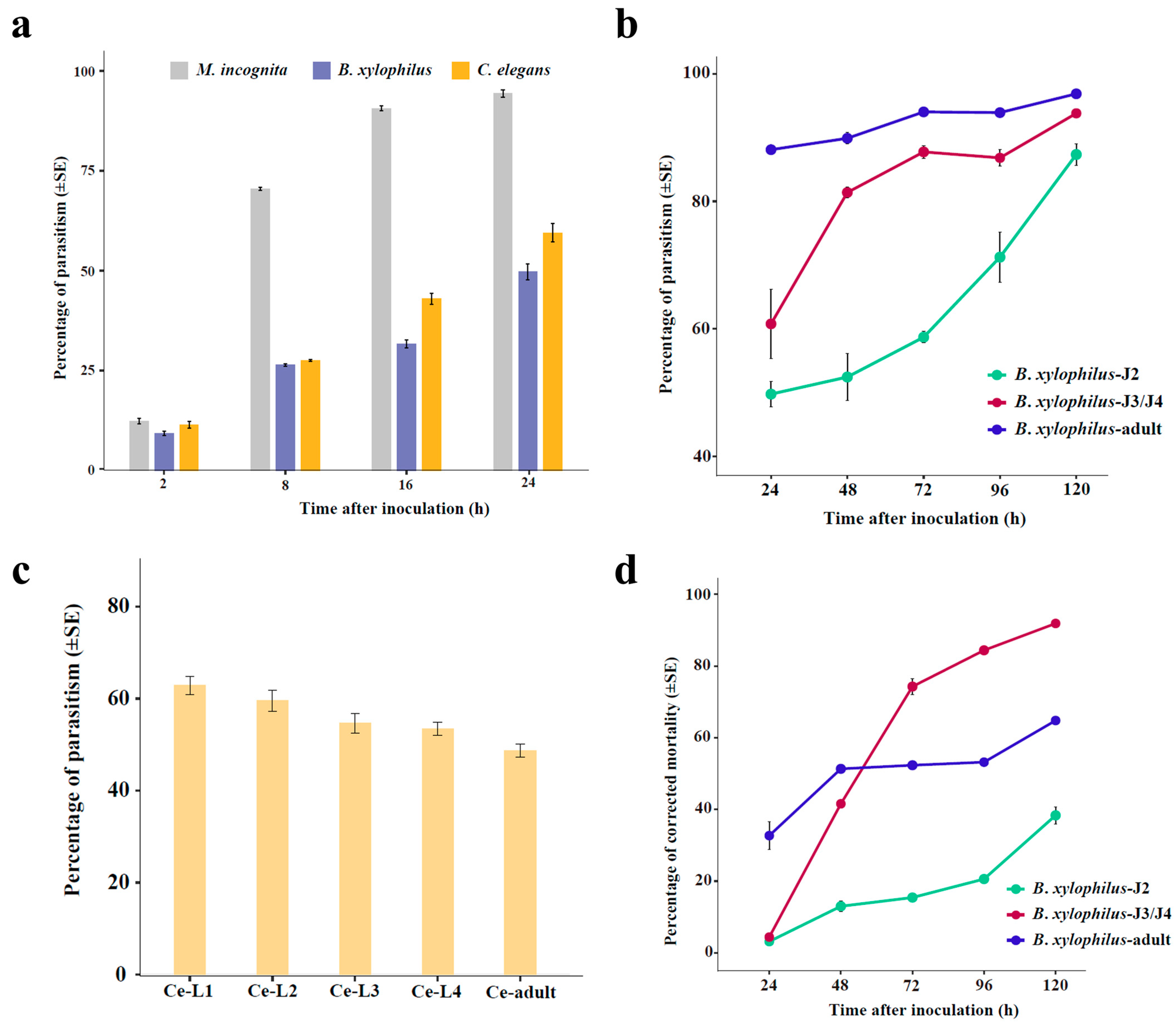
3.3. Comparison of the Fungal Parasitism on Different Nematodes
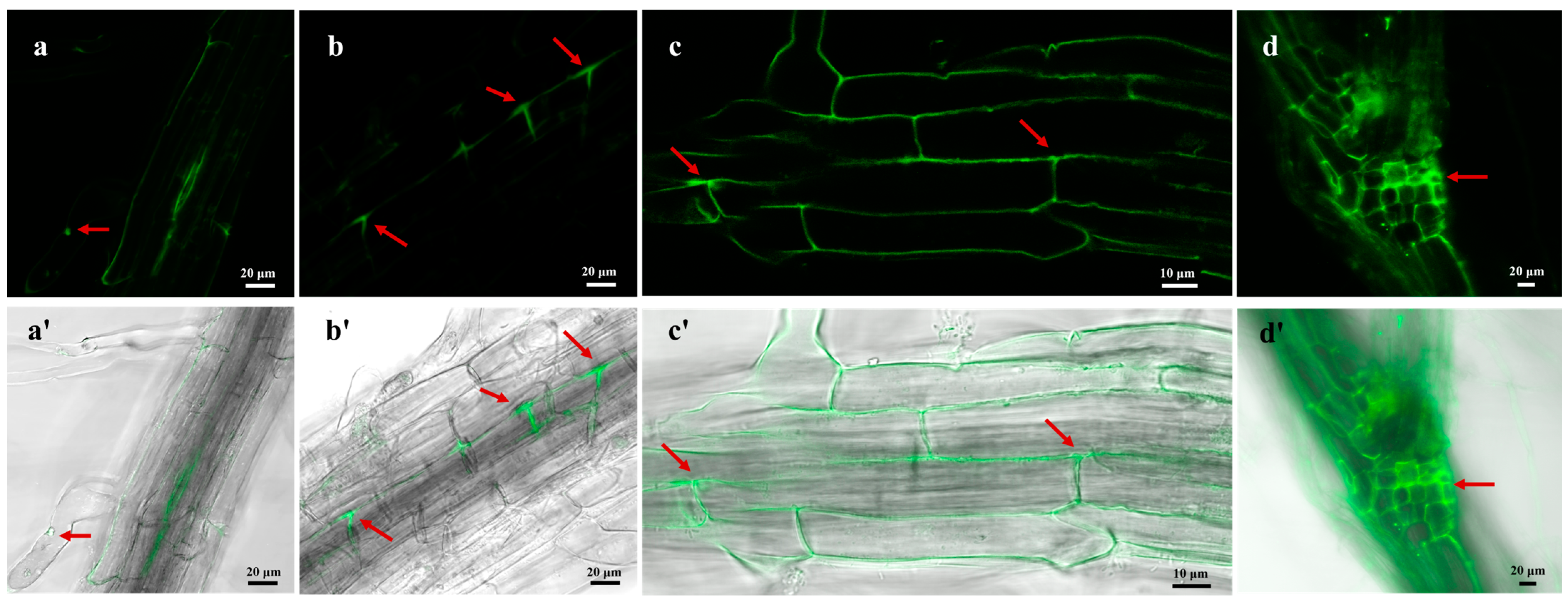
3.4. Endophytic Colonization of H. rhossiliensis HR02 on Arabidopsis Roots
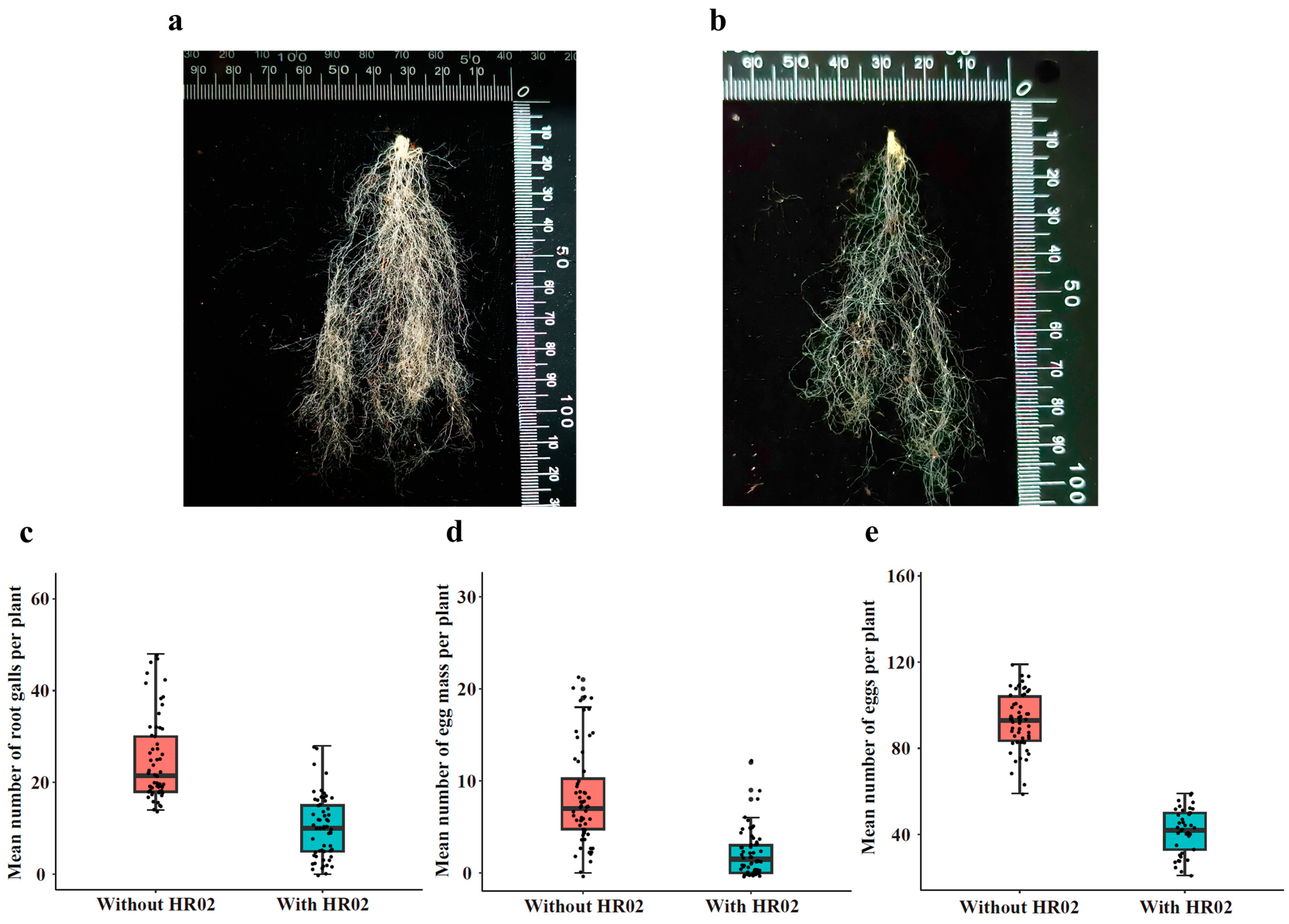
3.5. Effects of Endophytic H. rhossiliensis on Promoting Plant Growth
3.6. Effects of Endophytic H. rhossiliensis on Improving Plant Resistance to Root-Knot Nematodes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sturhan, D.; Schneider, R. Hirsutella heteroderae, a new nematode parasitic fungus. Phytopathol. Z. 1980, 99, 105–115. [Google Scholar] [CrossRef]
- Liu, X.Z.; Chen, S.Y. Parasitism of Heterodera glycines by Hirsutella spp. in Minnesota soybean fields. Biol. Control 2000, 19, 161–166. [Google Scholar] [CrossRef]
- Ma, R.; Liu, X.; Jian, H.; Li, S. Detection of Hirsutella spp. and Pasteuria sp. parasitizing second-stage juveniles of Heterodera glycines in soybean fields in China. Biol. Control 2005, 33, 223–229. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, Y.; Jiang, X.; Shu, C.; Hamid, M.I.; Hussain, M.; Chen, S.; Xu, J.; Xiang, M.; Liu, X. Population genetics of Hirsutella rhossiliensis, a dominant parasite of cyst nematode juveniles on a continental scale. Appl. Environ. Microbiol. 2016, 82, 6317–6325. [Google Scholar] [CrossRef]
- Lackey, B.A.; Muldoon, A.E.; Jaffee, B.A. Alginate pellet formulation of Hirsutella rhossiliensis for biological control of plant-parasitic nematodes. Biol. Control 1993, 3, 155–160. [Google Scholar] [CrossRef]
- Jaffee, B.A.; Gaspard, J.T.; Ferris, H. Density-dependent parasitism of the soil-borne nematode Criconemella xenoplax by the nematophagous fungus Hirsutella rhossiliensis. Microb. Ecol. 1989, 17, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Jaffee, B.A.; Ferris, H.; Stapleton, J.J.; Norton, M.V.K.; Muldoon, A.E. Parasitism of nematodes by the fungus Hirsutella rhossiliensis as affected by certain organic amendments. J. Nematol. 1994, 26, 152–161. [Google Scholar]
- Xiang, M.-C.; Yang, X.-H.; Wang, Z.-X.; Liu, X.-Z.; Chen, S.-Y.; Xiao, Q.-M. Variability of morphology, parasitism, and nucleotide sequences among isolates and species of nematophagous Hirsutella. Biol. Control 2007, 41, 110–119. [Google Scholar] [CrossRef]
- Jaffee, B.A.; Gaspard, J.T.; Ferris, H.; Muldoon, A.E. Quantification of parasitism of the soil-borne nematode Criconemella xenoplax by the nematophagous fungus Hirsutella rhossiliensis. Soil Biol. Biochem. 1988, 20, 631–636. [Google Scholar] [CrossRef]
- Cayrol, J.C.; Combettes, S. Étude de l’agressivité de quelques champignons nématophages vis-a-vis d’Anguina agrostis. Rev. Nématol. 1983, 6, 153–154. [Google Scholar]
- Ciancio, A.; Logrieco, A.; Lamberti, F. Parasitism of Xiphinema diversicaudatum by the fungus Hirsutella rhossiliensis. Nematol. Mediterr. 1986, 14, 187–192. [Google Scholar]
- Timper, P.; Kaya, H.K.; Jaffee, B.A. Survival of entomogenous nematodes in soil infested with the nematode-parasitic fungus Hirsutella rhossiliensis (Deuteromycotina: Hyphomycetes). Biol. Control. 1991, 1, 42–50. [Google Scholar] [CrossRef]
- Sun, J.; Xie, H.; Qiu, J.; Liu, X.; Xiang, M. Parasitism of secondary-stage juvenile of Heterodera glycines and four larva stages of Caenorhabditis elegans by Hirsutella spp. Exp. Parasitol. 2013, 135, 96–101. [Google Scholar] [CrossRef]
- Cayrol, J.C.; Castet, R.; Samson, R.A. Comparative activity of different Hirsutella species towards three plant parasitic nematodes. Rev. Nématol. 1986, 9, 412–414. [Google Scholar]
- Lopez-Llorca, L.V.; Bordallo, J.J.; Salinas, J.; Monfort, E.; López-Serna, M.L. Use of light and scanning electron microscopy to examine colonisation of barley rhizosphere by the nematophagous fungus Verticillium chlamydosporium. Micron 2002, 33, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Llorca, L.V.; Jansson, H.B.; Maciá-Vicente, J.G.; Salinas, J. Nematophagous fungi as root endophytes. In Microbial Root Endophytes; Schulz, B., Boyle, C., Sieber, T.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 191–206. [Google Scholar]
- Macia-Vicente, J.G.; Jansson, H.-B.; Lopez-Llorca, L.V. Understanding biological importance of endophytism of nematophagous fungi. In Prospects and Applications for Plant-Associated Microbes. A Laboratory Manual, Part B: Fungi; Pirttilä, A.M., Sorvari, S., Eds.; BioBien Innovations: Piikkiö, Finland, 2011; pp. 260–265. [Google Scholar]
- Bordallo, J.J.; Lopez-Llorca, L.V.; Jasson, H.-B.; Salinas, J.; Persmark, L.; Asensio, L. Colonization of plant roots by egg-parasitic and nematode-trapping fungi. New Phytol. 2002, 154, 491–499. [Google Scholar] [CrossRef]
- Monfort, E.; Lopez-Llorca, L.V.; Jansson, H.B.; Salinas, J.; Park, J.O.; Sivasithamparam, K. Colonisation of seminal roots of wheat and barley by egg-parasitic nematophagous fungi and their effects on Gaeumannomyces graminis var. tritici and development of root-rot. Soil Biol. Biochem. 2005, 37, 1229–1235. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Akhtar, M.S. Synergistic effects of antagonistic fungi and a plant growth promoting rhizobacterium, an arbuscular mycorrhizal fungus, or composted cow manure on populations of Meloydogyne incognita and growth of tomato. Biocontrol. Sci. Technol. 2008, 18, 207–290. [Google Scholar] [CrossRef]
- Escudero, N.; Lopez-Llorca, L.V. Effects on plant growth and root-knot nematode infection of an endophytic GFP transformant of the nematophagous fungus Pochonia chlamydosporia. Symbiosis 2012, 57, 33–42. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, W.; Huan, H.; Xia, W.; Chen, Y.; Wang, P.; Liu, Y. Construction of an integrated mCherry red fluorescent protein expression system for labeling and tracing in Lactiplantibacillus plantarum WCFS1. Front. Microbiol. 2021, 12, 690270. [Google Scholar] [CrossRef]
- Atkins, S.D.; Mauchline, T.H.; Kerry, B.R.; Hirsch, P.R. Development of a transformation system for the nematophagous fungus Pochonia chlamydosporia. Mycol. Res. 2004, 108, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Zavala-Gonzalez, E.A.; Rodríguez-Cazorla, E.; Escudero, N.; Aranda-Martinez, A.; Martínez-Laborda, A.; Ramírez-Lepe, M.; Vera, A.; Lopez-Llorca, L.V. Arabidopsis thaliana root colonization by the nematophagous fungus Pochonia chlamydosporia is modulated by jasmonate signaling and leads to accelerated flowering and improved yield. New Phytol. 2017, 213, 351–364. [Google Scholar] [CrossRef]
- Maciá-Vicente, J.G.; Jansson, H.B.; Talbot, N.J.; Lopez-Llorca, L.V. Real-time PCR quantification and live-cell imaging of endophytic colonization of barley (Hordeum vulgare) roots by Fusarium equiseti and Pochonia chlamydosporia. New Phytol. 2009, 182, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.-S.; Jeong, P.-Y.; Joo, H.-J.; Lee, J.-E.; Moon, Y.-S.; Cheon, H.-M.; Kim, J.-H.; Lee, Y.-U.; Shim, Y.-H.; Paik, Y.-K. Identification and characterization of a dual-acting antinematodal agent against the pinewood nematode, Bursaphelenchus xylophilus. PLoS ONE 2009, 4, e7593. [Google Scholar] [CrossRef] [PubMed]
- Vrain, T.; Wakarchuk, D.; Levesque, A.; Hamilton, R. Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fundam. Appl. Nematol. 1992, 15, 563–573. [Google Scholar]
- Janssen, T.; Karssen, G.; Verhaeven, M.; Coyne, D.; Bert, W. Mitochondrial coding genome analysis of tropical root-knot nematodes (Meloidogyne) supports haplotype based diagnostics and reveals evidence of recent reticulate evolution. Sci. Rep. 2016, 6, 22591. [Google Scholar] [CrossRef] [PubMed]
- Randig, O.; Bongiovanni, M.; Carneiro, R.M.; Castagnone-Sereno, P. Genetic diversity of root-knot nematodes from Brazil and development of SCAR marker specific for the coffee damaging species. Genome 2002, 45, 862–870. [Google Scholar] [CrossRef]
- Herndon, L.A.; Wolkow, C.A.; Driscoll, M.; Hall, D.H. Effects of ageing on the basic biology and anatomy of C. elegans. In Ageing: Lessons from C. elegans. Healthy Ageing and Longevity; Olsen, A., Gill, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 9–39. [Google Scholar]
- Porta-de-la-Riva, M.; Fontrodona, L.; Villanueva, A.; Cerón, J. Basic Caenorhabditis elegans methods: Synchronization and observation. J. Vis. Exp. 2012, 64, e4019. [Google Scholar] [CrossRef]
- Zhuang, X.; Zhao, J.-L.; Bai, M.; Ping, X.-X.; Li, Y.-L.; Yang, Y.-H.; Mao, Z.-C.; Yang, G.-S.; Xie, B.-Y. Pochonia chlamydosporia isolate PC-170-induced expression of marker genes for defense pathways in tomatoes challenged by different pathogens. Microorganisms 2021, 9, 1882. [Google Scholar] [CrossRef]
- Zou, M.; Xin, B.; Sun, X.; Lin, R.; Lu, J.; Qi, J.; Xie, B.; Cheng, X. URA3 as a selectable marker for disruption and functional assessment of PacC gene in the entomopathogenic fungus Isaria javanica. J. Fungi 2023, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, H.T. A protocol describing pharynx counts and a review of other assays of apoptotic cell death in the nematode worm Caenorhabditis elegans. Nat. Protoc. 2007, 2, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Jaffee, B.A.; Zehr, E.I. Sporulation of the fungus Hirsutella rhossiliensis from the nematode Criconemella xenoplax. Plant Dis. 1983, 67, 1265–1267. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE Publications, Inc.: London, UK, 2018. [Google Scholar]
- Tedford, E.C.; JaVee, B.A.; Muldoon, A.E. Effect of soil moisture and texture on transmission of the nematophagous fungus Hirsutella rhossiliensis to cyst and root-knot nematodes. Phytopathology 1992, 82, 1002–1007. [Google Scholar] [CrossRef]
- Tedford, E.C.; JaVee, B.A. In vitro parasitism of Rotylenchus robustus by isolates of Hirsutella rhossiliensis. J. Nematol. 1995, 22, 486–489. [Google Scholar]
- Mesny, F.; Miyauchi, S.; Thiergart, T.; Pickel, B.; Atanasova, L.; Karlsson, M.; Hüttel, B.; Barry, K.W.; Haridas, S.; Chen, C.; et al. Genetic determinants of endophytism in the Arabidopsis root mycobiome. Nat. Comm. 2021, 12, 7227. [Google Scholar] [CrossRef]
- Poveda, J.; Eugui, D.; Abril-Urias, P.; Velasco, P. Endophytic fungi as direct plant growth promoters for sustainable agricultural production. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Liao, J.; Lu, J.; Lin, R.; Zou, M.; Xie, B.; Cheng, X. Parasitism of Hirsutella rhossiliensis on Different Nematodes and Its Endophytism Promoting Plant Growth and Resistance against Root-Knot Nematodes. J. Fungi 2024, 10, 68. https://doi.org/10.3390/jof10010068
Sun X, Liao J, Lu J, Lin R, Zou M, Xie B, Cheng X. Parasitism of Hirsutella rhossiliensis on Different Nematodes and Its Endophytism Promoting Plant Growth and Resistance against Root-Knot Nematodes. Journal of Fungi. 2024; 10(1):68. https://doi.org/10.3390/jof10010068
Chicago/Turabian StyleSun, Xin, Jiaqian Liao, Junru Lu, Runmao Lin, Manling Zou, Bingyan Xie, and Xinyue Cheng. 2024. "Parasitism of Hirsutella rhossiliensis on Different Nematodes and Its Endophytism Promoting Plant Growth and Resistance against Root-Knot Nematodes" Journal of Fungi 10, no. 1: 68. https://doi.org/10.3390/jof10010068
APA StyleSun, X., Liao, J., Lu, J., Lin, R., Zou, M., Xie, B., & Cheng, X. (2024). Parasitism of Hirsutella rhossiliensis on Different Nematodes and Its Endophytism Promoting Plant Growth and Resistance against Root-Knot Nematodes. Journal of Fungi, 10(1), 68. https://doi.org/10.3390/jof10010068