
Proliferation and Apoptosis of Cat (Felis catus) Male Germ Cells during Breeding and Non-Breeding Seasons



{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Sample Collection
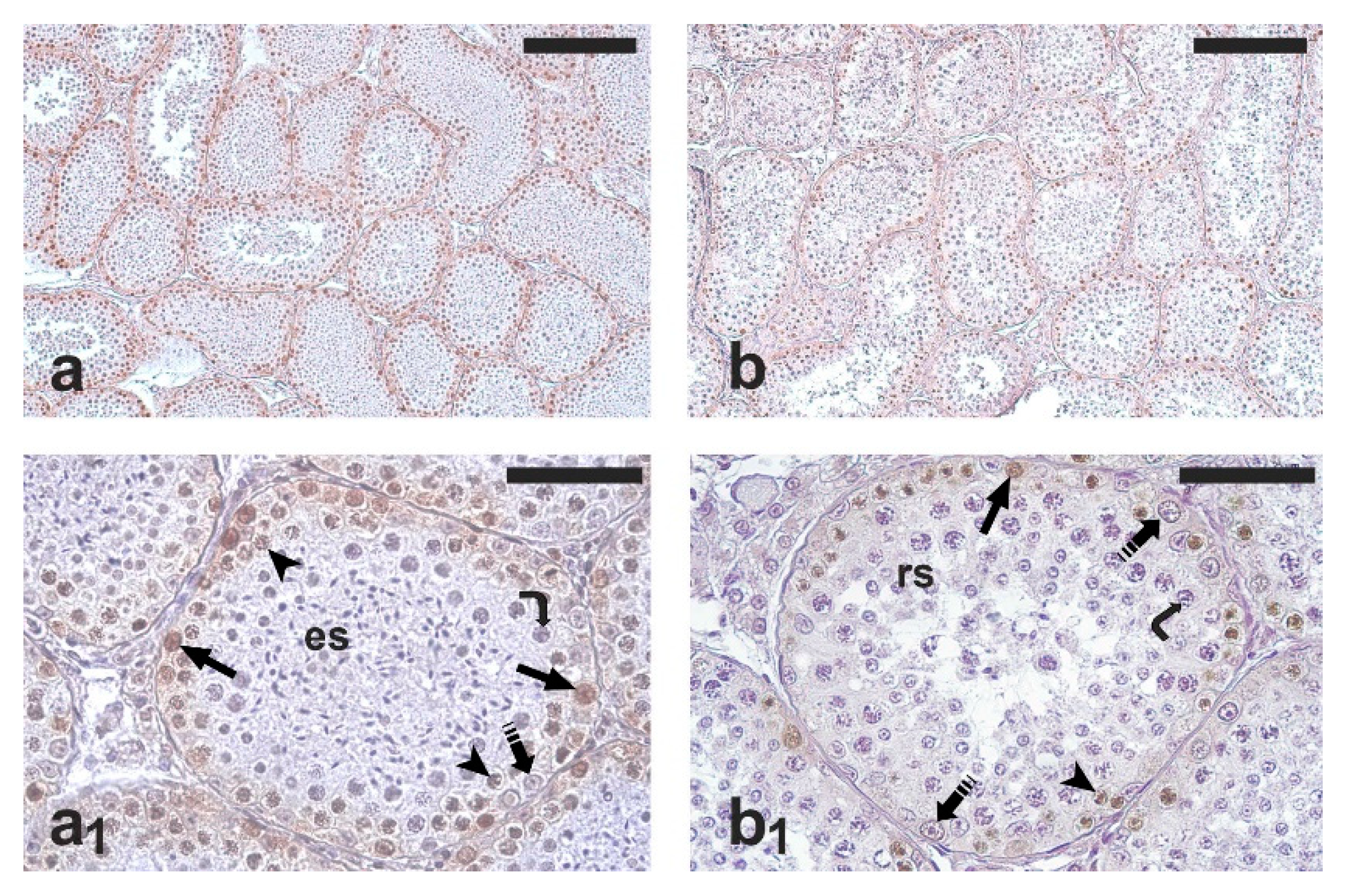
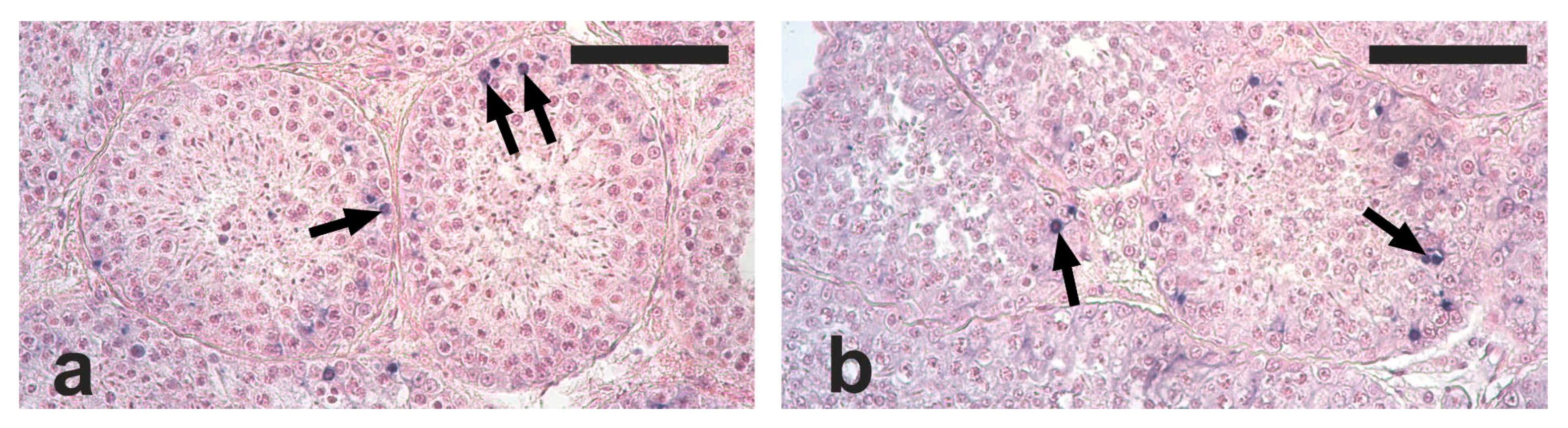
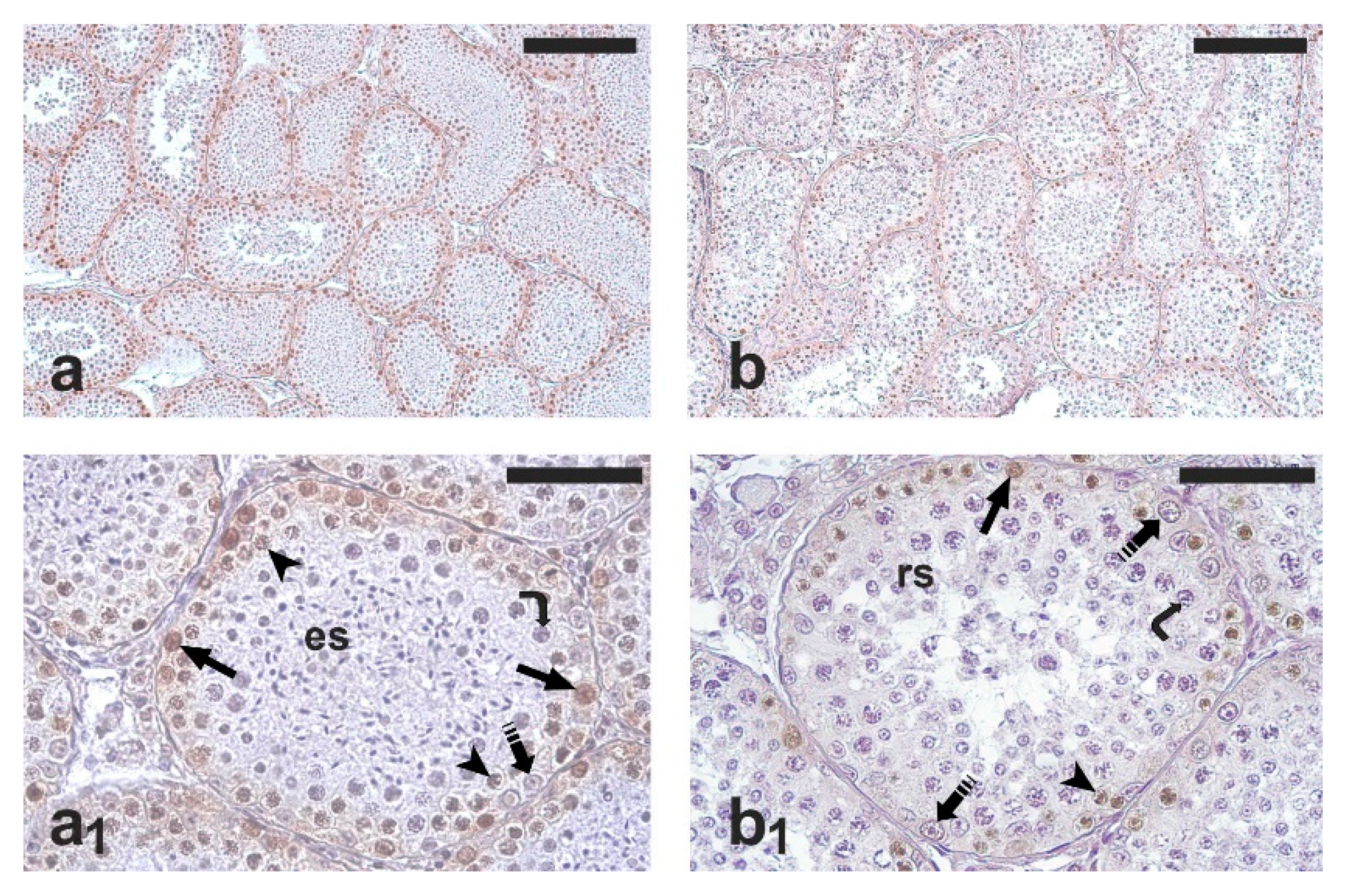
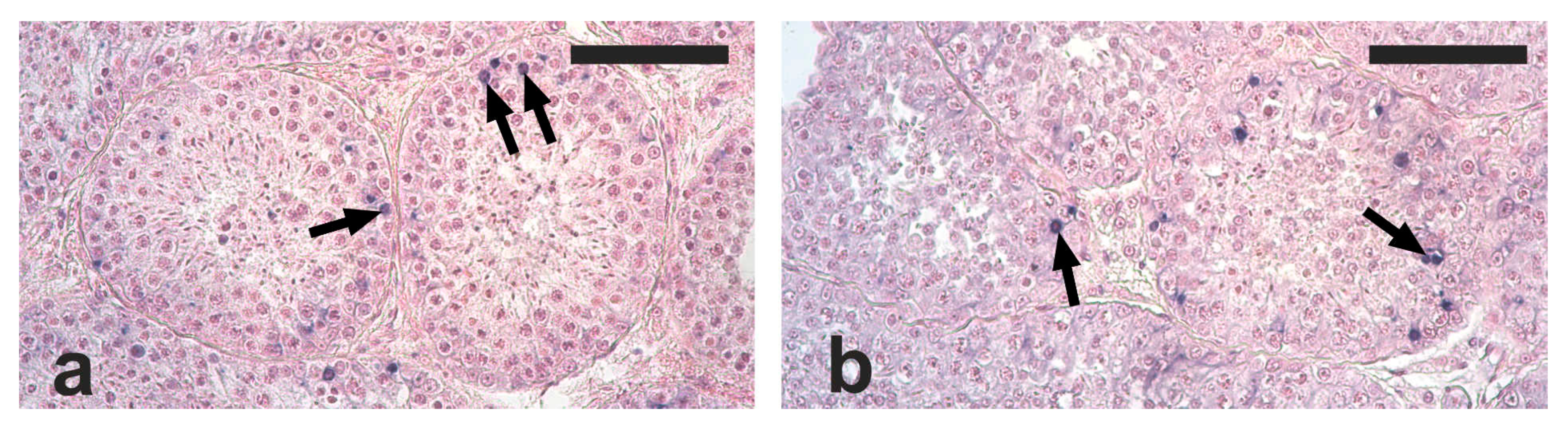
2.2. Testis Histology, Immunohistochemical Detection of Proliferating Germ Cells and Apoptosis
2.3. Quantification of Germ Cell Proliferation and Apoptosis
2.4. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roosen-Runge, E.C. The process of spermatogenesis in mammals. Biol. Rev. 1962, 37, 343–376. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lu, H.; Li, H.; Yan, C.; Wang, X.; Zang, M.; Rooij, D.G.; Madabhushi, A.; Xu, E.Y. Computerized spermatogenesis staging (CSS) of mouse testis sections via quantitative histomorphological analysis. Med. Image Anal. 2021, 70, 101835. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, D.G. Proliferation and differentiation of spermatogonial stem cells. Reproduction 2001, 121, 347–354. [Google Scholar] [CrossRef] [PubMed]
- França, L.R.; Godinho, C.L. Testis morphometry, seminiferous epithelium cycle length, and daily sperm production in domestic cats (Felis catus). Biol. Reprod. 2003, 68, 1554–1561. [Google Scholar] [CrossRef] [Green Version]
- Bristol-Gould, S.; Woodruff, T.K. Folliculogenesis in the domestic cat (Felis catus). Theriogenology 2006, 66, 5–13. [Google Scholar] [CrossRef]
- Schäfer-Somi, S. Effect of melatonin on the reproductive cycle in female cats: A review of clinical experiences and previous studies. J. Feline Med. Surg. 2017, 19, 5–12. [Google Scholar] [CrossRef]
- Spindler, R.E.; Wildt, D.E. Circannual variations in intraovarian oocyte but not epididymal sperm quality in the domestic cat. Biol. Reprod. 1999, 61, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Wildt, D.E.; Brown, J.L.; Swanson, W.F. Cats. In Encyclopedia of Reproduction; Knobil, E., Neill, J., Eds.; Academic Press: New York, NY, USA, 1999; Volume 1, pp. 497–510. [Google Scholar]
- Blanco-Rodríguez, J. DNA replication and germ cell apoptosis during spermatogenesis in the cat. J. Androl. 2002, 23, 484–490. [Google Scholar] [CrossRef]
- Kirkpatrick, J.F. Seasonal testosterone levels, testosterone clearance, and testicular weights in male domestic cats. Canad. J. Zool. 1985, 63, 1285–1287. [Google Scholar] [CrossRef]
- Nuñez-Favre, R.; Bonaura, M.C.; Tittarelli, C.M.; Mansilla-Hermann, D.; de la Sota, R.L.; Stornelli, M.A. Effect of natural photoperiod on epididymal sperm quality and ttosterone serum concentration in domestic cat (Felis silvestris catus). Reprod. Domest. Anim. 2012, 47 (Suppl. 6), 232–234. [Google Scholar] [CrossRef]
- Blottner, S.; Jewgenow, K. Moderate seasonality in testis function of domestic cat. Reprod. Domest. Anim. 2007, 42, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Blottner, S.; Hingst, O.; Meyer, H.H.D. Inverse relationship between testicular proliferation and apoptosis in mammalian seasonal breeders. Theriogenology 1995, 44, 320–328. [Google Scholar] [CrossRef]
- Young, K.A.; Nelson, R.J. Mediation of seasonal testicular regression by apoptosis. Reproduction 2001, 122, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Štrbenc, M.; Fazarinc, G.; Bavdek, S.V.; Pogacnik, A. Apoptosis and proliferation during seasonal testis regression in the brown hare (Lepus europaeus L.). Anat. Histol. Embryol. 2003, 32, 48–53. [Google Scholar] [CrossRef]
- Prisco, M.; Liguoro, A.; Comitato, R.; Cardone, A.; D’Onghia, B.; Ricchiari, L.; Angelini, F.; Andreucci, P. Apoptosis during spermatogenesis in the spotted ray Torpedo marmorata. Mol. Reprod. Dev. 2003, 64, 341–348. [Google Scholar] [CrossRef]
- Corriero, A.; Desantis, S.; Bridges, C.R.; Kime, D.E.; Megalofonou, P.; Santamaria, N.; Cirillo, F.; Ventriglia, G.; Di Summa, A.; Deflorio, M.; et al. Germ cell proliferation and apoptosis during different phases of swordfish (Xiphias gladius L.) spermatogenetic cycle. J. Fish Biol. 2007, 70, 83–99. [Google Scholar] [CrossRef]
- Zupa, R.; Fauvel, C.; Mylonas, C.C.; Santamaria, N.; Valentini, L.; Pousis, C.; Papadaki, M.; Suquet, M.; de la Gándara, F.; Bello, G.; et al. Comparative analysis of male germ cell proliferation and apoptosis in wild and captive Atlantic bluefin tuna (Thunnus thynnus L.). J. Appl. Ichthyol. 2013, 29, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Wang, L.; Chen, H.; Huang, Y.; Yang, P.; Ahmed, N.; Wang, T.; Liu, Y.; Chen, Q. Molecular and cellular mechanisms of apoptosis during dissociated spermatogenesis. Front. Physiol. 2017, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Huang, Y.; Liu, T.; Haseeb, A.; Ahmed, N.; Zhang, L.; Bian, X.; Chen, Q. Characteristics of seasonal spermatogenesis in the soft-shelled Turtle. Anim. Reprod. Sci. 2020, 214, 106307. [Google Scholar] [CrossRef]
- Scaia, M.F.; Czuchlej, S.C.; Cervino, N.; Ceballos, N.R. Apoptosis, proliferation and presence of estradiol receptors in the testes and Bidder’s organ of the toad Rhinella arenarum (Amphibia, Anura). J. Morphol. 2016, 277, 412–423. [Google Scholar] [CrossRef]
- Wang, D.-H.; Hu, J.-R.; Wang, L.-Y.; Hu, Y.-J.; Tan, F.-Q.; Zhou, H.; Shao, J.-Z.; Yang, W.-X. The apoptotic function analysis of p53, Apaf1, Caspase3 and Caspase7 during the spermatogenesis of the chinese fire-bellied newt Cynops orientalis. PLoS ONE 2012, 7, e39920. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, L.K.; Ross, W.L.; Young, K.A. Increases in apoptosis and declines in Bcl-XL protein characterise testicular regression in American crows (Corvus brachyrhynchos). Reprod. Fertil. Dev. 2007, 19, 461–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Rooij, D.G.; Grootegoed, J.A. Spermatogonial stem cells. Curr. Opin. Cell Biol. 1998, 10, 694–701. [Google Scholar] [CrossRef]
- de Rooij, D.G.; Russell, L.D. All you wanted to know about spermatogonia but were afraid to ask. J. Androl. 2000, 21, 776–798. [Google Scholar] [CrossRef]
- Wingfield, J.C.; Kenagy, G.J. Natural regulation of reproductive cycles. In Vertebrate Endocrinology: Fundamental and Biomedical Implications; Pang, P., Schreibman, M., Eds.; Academic Press: New York, NY, USA, 1991; Volume 4B, pp. 181–241. [Google Scholar]
- Bronson, F.H.; Heideman, P.D. Seasonal regulation of reproduction in mammals. In The Physiology of Reproduction; Knobil, E., Neill, J.D., Eds.; Raven Press: New York, NY, USA, 1994; pp. 541–583. [Google Scholar]
- Stornelli, M.A.; Reyna, J.C.; Stornelli, M.C.; Nuñez Favre, R.; Savignone, C.A.; Tittarelli, C.M.; de la Sota, R.L. Seasonal changes in testicular cell morphology in domestic male cats (Felis catus). Reprod. Domest. Anim. 2009, 44 (Suppl. 2), 287–290. [Google Scholar] [CrossRef]
- Yin, Y.; Stahl, B.; De Wolf, W.C.; Morgentaler, A. p53-mediated germ cell quality control in spermatogenesis. Dev. Biol. 1998, 204, 165–171. [Google Scholar] [CrossRef]
- Desjardins, C.; Lopez, M.J. Environmental cues evoke differential responses in pituitary–testicular function in deer mice. Endocrinology 1983, 112, 1398–1406. [Google Scholar] [CrossRef]
- Siemieniuch, M.J. Apoptotic changes in the epithelium germinativum of the cat (Felis catus S. domestica, L. 1758) at different ages and breeding seasons. Reprod. Domest. Anim. 2008, 43, 473–476. [Google Scholar] [CrossRef]
- Prochowska, S.; Partyka, A.; Niżański, W. Expression of apoptosis-related genes in cat testicular tissue in relation to sperm morphology and seasonality—A preliminary study. Animals 2021, 11, 489. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valentini, L.; Zupa, R.; Pousis, C.; Cuko, R.; Corriero, A. Proliferation and Apoptosis of Cat (Felis catus) Male Germ Cells during Breeding and Non-Breeding Seasons. Vet. Sci. 2022, 9, 447. https://doi.org/10.3390/vetsci9080447
Valentini L, Zupa R, Pousis C, Cuko R, Corriero A. Proliferation and Apoptosis of Cat (Felis catus) Male Germ Cells during Breeding and Non-Breeding Seasons. Veterinary Sciences. 2022; 9(8):447. https://doi.org/10.3390/vetsci9080447
Chicago/Turabian StyleValentini, Luisa, Rosa Zupa, Chrysovalentinos Pousis, Rezart Cuko, and Aldo Corriero. 2022. "Proliferation and Apoptosis of Cat (Felis catus) Male Germ Cells during Breeding and Non-Breeding Seasons" Veterinary Sciences 9, no. 8: 447. https://doi.org/10.3390/vetsci9080447
APA StyleValentini, L., Zupa, R., Pousis, C., Cuko, R., & Corriero, A. (2022). Proliferation and Apoptosis of Cat (Felis catus) Male Germ Cells during Breeding and Non-Breeding Seasons. Veterinary Sciences, 9(8), 447. https://doi.org/10.3390/vetsci9080447