Effect of Cannabidiol (CBD) on Canine Inflammatory Response: An Ex Vivo Study on LPS Stimulated Whole Blood

 ,
,  , , ,
, , ,  ,
,  ,
,  , ,
, ,  ,
,  and
and 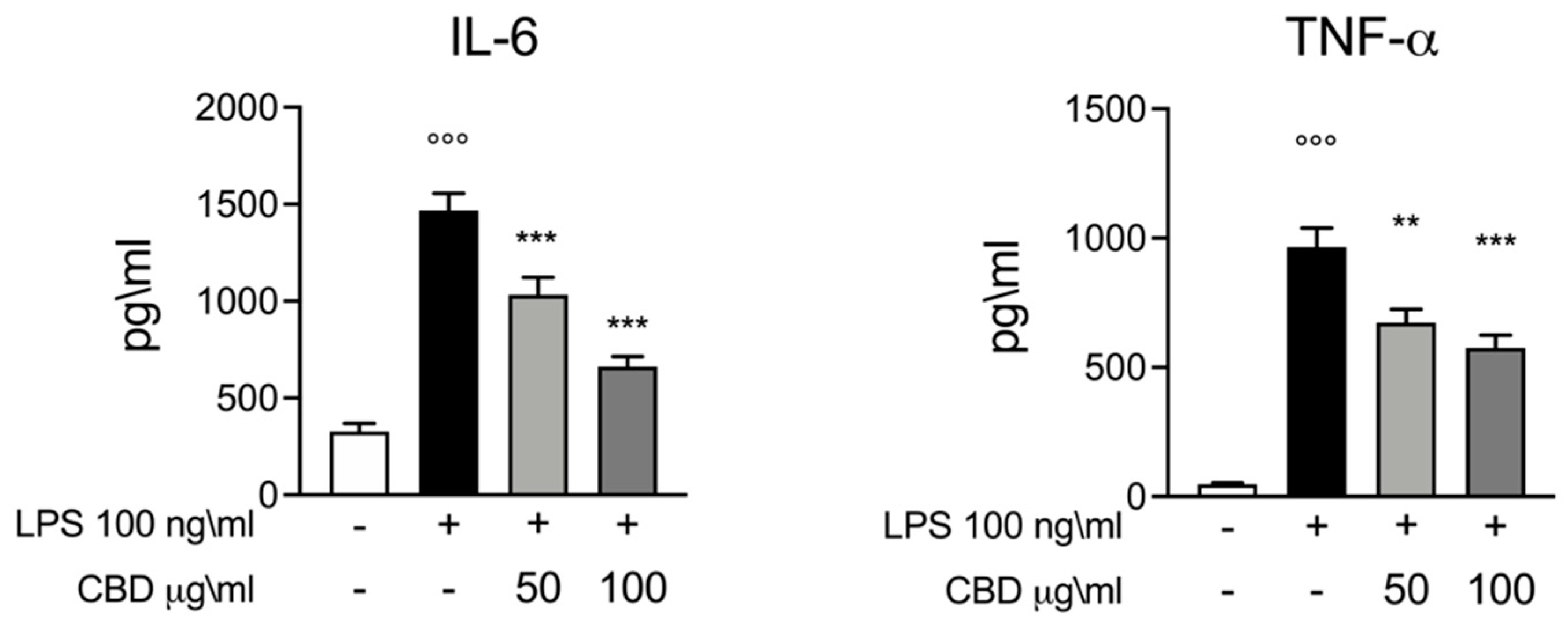{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Canine Donors and Blood Collection
2.2. Whole Blood Culture and Stimulation
- Control: whole blood + vehicle (DMSO 0.1%)
- LPS: whole blood + LPS 100 ng/mL + vehicle (DMSO 0.1%)
- CBD 50 μg: whole blood + LPS 100 ng/mL + CBD (kanarescue 5% in DMSO 0.1%) 50 μg/mL
- CBD 100 μg: whole blood + LPS 100 ng/mL CBD (kanarescue 5% in DMSO 0.1%) 100 μg/mL
2.3. ELISA Assays
2.4. rt-PCR
2.5. Western Blot
2.6. Statistical Analysis
3. Results
3.1. Effect of CBD on IL-6 and TNF-α Response in LPS Stimulated Whole Blood
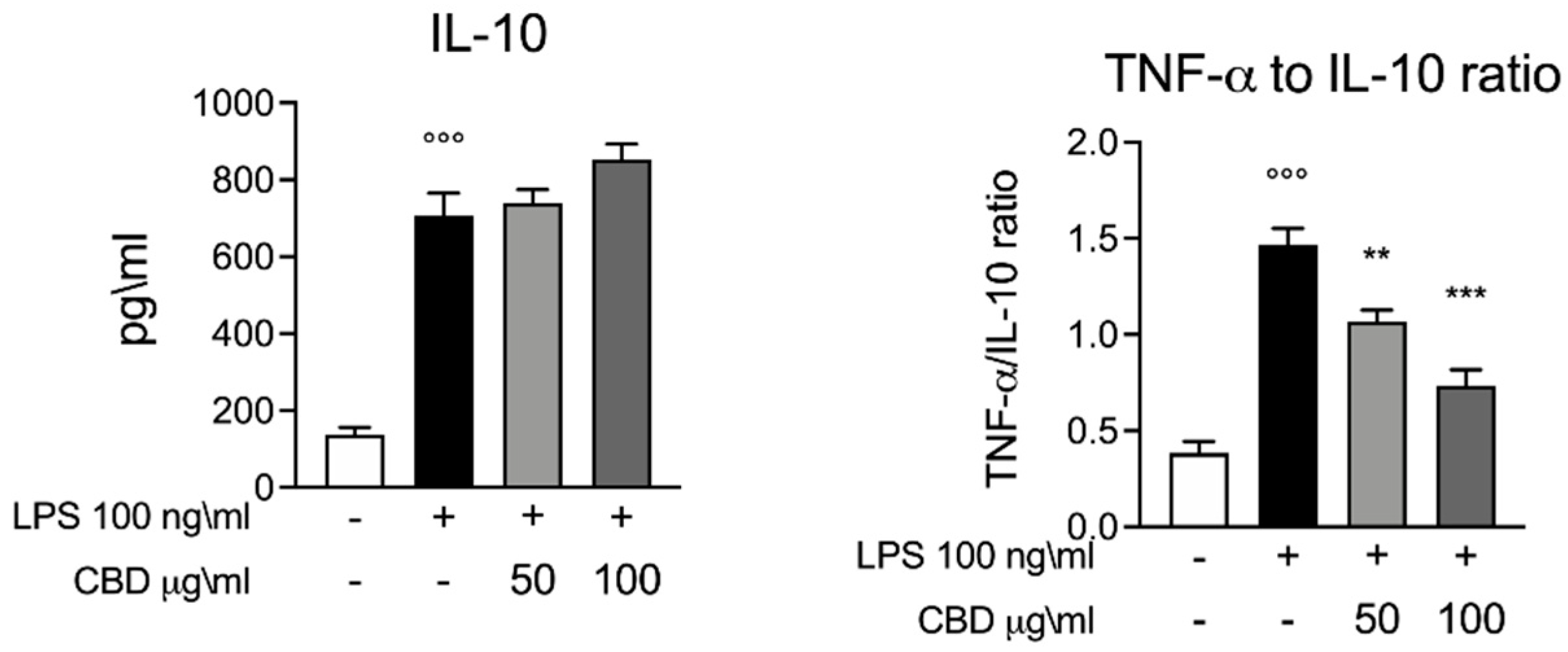
3.2. Effect of CBD on IL-10 in LPS Stimulated Whole Blood
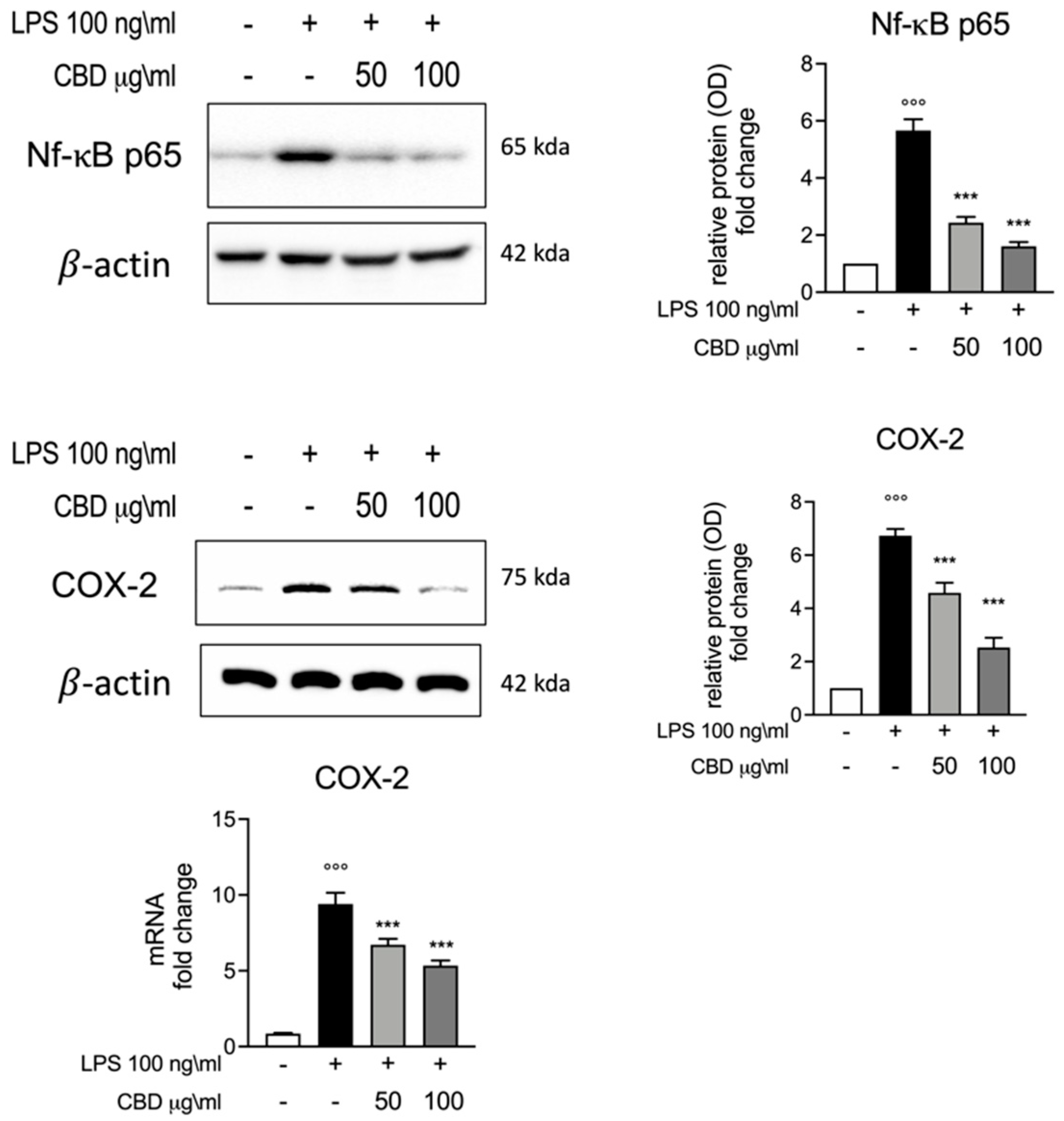
3.3. Effect of CBD on Nf-κB and COX-2 Induction in LPS Stimulated Whole Blood
4. Discussion
5. Limitations of the Study
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ritter, S.; Zadik-Weiss, L.; Almogi-Hazan, O.; Or, R. Cannabis, One Health, and Veterinary Medicine: Cannabinoids’ Role in Public Health, Food Safety, and Translational Medicine. Rambam Maimonides Med. J. 2020, 11. [Google Scholar] [CrossRef]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Properties of Cannabidiol. Antioxidants 2019, 9, 21. [Google Scholar] [CrossRef]
- Della Rocca, G.; Di Salvo, A. Hemp in Veterinary Medicine: From Feed to Drug. Front. Vet. Sci. 2020, 7, 387. [Google Scholar] [CrossRef]
- Vaughn, D.M.; Paulionis, L.J.; Kulpa, J.E. Randomized, placebo-controlled, 28-day safety and pharmacokinetics evaluation of repeated oral cannabidiol administration in healthy dogs. Am. J. Vet. Res. 2021, 82, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Mejia, S.; Duerr, F.M.; Griffenhagen, G.; McGrath, S. Evaluation of the Effect of Cannabidiol on Naturally Occurring Osteoarthritis-Associated Pain: A Pilot Study in Dogs. J. Am. Anim. Hosp. Assoc. 2021, 57, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.H.J.; Rupasinghe, H.P.V. Cannabidiol-based natural health products for companion animals: Recent advances in the management of anxiety, pain, and inflammation. Res. Vet. Sci. 2021, 140, 38–46. [Google Scholar] [CrossRef]
- Pagano, S.; Coniglio, M.; Valenti, C.; Federici, M.I.; Lombardo, G.; Cianetti, S.; Marinucci, L. Biological effects of Cannabidiol on normal human healthy cell populations: Systematic review of the literature. Biomed. Pharmacother. 2020, 132, 110728. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, L.; Butcher, B.; Henson, J.D.; Rutolo, D.; Hall, S. A pilot safety, tolerability and pharmacokinetic study of an oro-buccal administered cannabidiol-dominant anti-inflammatory formulation in healthy individuals: A randomized placebo-controlled single-blinded study. Inflammopharmacology 2021. [Google Scholar] [CrossRef]
- Spierenburg, E.A.J.; Portengen, L.; Smit, L.A.M.; Krop, E.J.M.; Hylkema, M.N.; Rijkers, G.T.; Heederik, D.; Wouters, I.M. Stability of individual LPS-induced ex vivo cytokine release in a whole blood assay over a five-year interval. J. Immunol. Methods 2018, 460, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Burnsides, C.; Corry, J.; Alexander, J.; Balint, C.; Cosmar, D.; Phillips, G.; Marketon, J.I. Ex vivo stimulation of whole blood as a means to determine glucocorticoid sensitivity. J. Inflamm. Res. 2012, 5, 89–97. [Google Scholar] [CrossRef][Green Version]
- Kathrani, A.; Hall, E. A preliminary study assessing cytokine production following ex vivo stimulation of whole blood with diet in dogs with chronic enteropathy. BMC Vet. Res. 2019, 15, 185. [Google Scholar] [CrossRef] [PubMed]
- Peyravian, N.; Deo, S.; Daunert, S.; Jimenez, J.J. Cannabidiol as a Novel Therapeutic for Immune Modulation. Immunotargets Ther. 2020, 9, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.L.; Springs, A.E.; Kaminski, N.E. The profile of immune modulation by cannabidiol (CBD) involves deregulation of nuclear factor of activated T cells (NFAT). Biochem. Pharmacol. 2008, 76, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Angelone, D.F.; Wessels, M.R.; Coughlin, M.; Suter, E.E.; Valentini, P.; Kalish, L.A.; Levy, O. Innate immunity of the human newborn is polarized toward a high ratio of IL-6/TNF-alpha production in vitro and in vivo. Pediatr. Res. 2006, 60, 205–209. [Google Scholar] [CrossRef]
- Fowler, B.L.; Axiak, S.M.; DeClue, A.E. Blunted pathogen-associated molecular pattern motif induced TNF, IL-6 and IL-10 production from whole blood in dogs with lymphoma. Vet. Immunol. Immunopathol. 2011, 144, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; D’Amico, R.; Cordaro, M.; Crupi, R.; Mandalari, G.; Impellizzeri, D.; et al. The Role of Cashew (Anacardium occidentale L.) Nuts on an Experimental Model of Painful Degenerative Joint Disease. Antioxidants 2020, 9, 511. [Google Scholar] [CrossRef]
- Sarr, O.; Dyck, D.J.; Mutch, D.M. [Letter to the Editor] Protein phosphorylation status is preserved following dual RNA and protein extraction using the Qiagen RNeasy Mini Kit. BioTechniques 2016, 61, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Siracusa, R.; Impellizzeri, D.; D’Amico, R.; Peritore, A.F.; Crupi, R.; Gugliandolo, E.; Fusco, R.; Di Paola, R.; Schievano, C.; et al. Safety and efficacy of a new micronized formulation of the ALIAmide palmitoylglucosamine in preclinical models of inflammation and osteoarthritis pain. Arthritis. Res. Ther. 2019, 21, 254. [Google Scholar] [CrossRef]
- Esposito, E.; Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Siracusa, R.; Gugliandolo, E.; Crupi, R.; Cuzzocrea, S. A new co-micronized composite containing palmitoylethanolamide and polydatin shows superior oral efficacy compared to their association in a rat paw model of carrageenan-induced inflammation. Eur. J. Pharmacol. 2016, 782, 107–118. [Google Scholar] [CrossRef]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Campolo, M.; Latteri, S.; Carughi, A.; Mandalari, G.; Cuzzocrea, S. The Antioxidant Activity of Pistachios Reduces Cardiac Tissue Injury of Acute Ischemia/Reperfusion (I/R) in Diabetic Streptozotocin (STZ)-Induced Hyperglycaemic Rats. Front. Pharmacol. 2018, 9, 51. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Siracusa, R.; D’Amico, R.; Peritore, A.F.; Cordaro, M.; Gugliandolo, E.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Melatonin Plus Folic Acid Treatment Ameliorates Reserpine-Induced Fibromyalgia: An Evaluation of Pain, Oxidative Stress, and Inflammation. Antioxidants 2019, 8, 628. [Google Scholar] [CrossRef]
- Jaffey, J.A.; Amorim, J.; DeClue, A.E. Effect of calcitriol on in vitro whole blood cytokine production in critically ill dogs. Vet. J. 2018, 236, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Gamble, L.J.; Boesch, J.M.; Frye, C.W.; Schwark, W.S.; Mann, S.; Wolfe, L.; Brown, H.; Berthelsen, E.S.; Wakshlag, J.J. Pharmacokinetics, Safety, and Clinical Efficacy of Cannabidiol Treatment in Osteoarthritic Dogs. Front. Vet. Sci. 2018, 5, 165. [Google Scholar] [CrossRef]
- McGrath, S.; Bartner, L.R.; Rao, S.; Packer, R.A.; Gustafson, D.L. Randomized blinded controlled clinical trial to assess the effect of oral cannabidiol administration in addition to conventional antiepileptic treatment on seizure frequency in dogs with intractable idiopathic epilepsy. J. Am. Vet. Med. Assoc. 2019, 254, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Tanner, C.; Mechoulam, R.; Pertwee, R.G.; Ross, R.A. Inhibition of human neutrophil chemotaxis by endogenous cannabinoids and phytocannabinoids: Evidence for a site distinct from CB1 and CB2. Mol. Pharmacol. 2008, 73, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Lodzki, M.; Godin, B.; Rakou, L.; Mechoulam, R.; Gallily, R.; Touitou, E. Cannabidiol-transdermal delivery and anti-inflammatory effect in a murine model. J. Control. Release. 2003, 93, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Jan, T.R.; Su, S.T.; Wu, H.Y.; Liao, M.H. Suppressive effects of cannabidiol on antigen-specific antibody production and functional activity of splenocytes in ovalbumin-sensitized BALB/c mice. Int. Immunopharmacol. 2007, 7, 773–780. [Google Scholar] [CrossRef]
- Nichols, J.M.; Kaplan, B.L.F. Immune Responses Regulated by Cannabidiol. Cannabis Cannabinoid Res. 2020, 5, 12–31. [Google Scholar] [CrossRef]
- Bebee, B.; Taylor, D.M.; Bourke, E.; Pollack, K.; Foster, L.; Ching, M.; Wong, A. The CANBACK trial: A randomised, controlled clinical trial of oral cannabidiol for people presenting to the emergency department with acute low back pain. Med. J. Aust. 2021, 214, 370–375. [Google Scholar] [CrossRef]
- De Briyne, N.; Holmes, D.; Sandler, I.; Stiles, E.; Szymanski, D.; Moody, S.; Neumann, S.; Anadon, A. Cannabis, Cannabidiol Oils and Tetrahydrocannabinol-What Do Veterinarians Need to Know? Animals 2021, 11, 892. [Google Scholar] [CrossRef]
- Mlost, J.; Bryk, M.; Starowicz, K. Cannabidiol for Pain Treatment: Focus on Pharmacology and Mechanism of Action. Int. J. Mol. Sci. 2020, 21, 8870. [Google Scholar] [CrossRef]
- Tran, T.A.T.; Grievink, H.W.; Lipinska, K.; Kluft, C.; Burggraaf, J.; Moerland, M.; Tasev, D.; Malone, K.E. Whole blood assay as a model for in vitro evaluation of inflammasome activation and subsequent caspase-mediated interleukin-1 beta release. PLoS ONE 2019, 14, e0214999. [Google Scholar] [CrossRef]
- Schmitz, S.; Henrich, M.; Neiger, R.; Werling, D.; Allenspach, K. Comparison of TNFalpha responses induced by Toll-like receptor ligands and probiotic Enterococcus faecium in whole blood and peripheral blood mononuclear cells of healthy dogs. Vet. Immunol. Immunopathol. 2013, 153, 170–174. [Google Scholar] [CrossRef]
- Messerer, D.A.C.; Vidoni, L.; Erber, M.; Stratmann, A.E.P.; Bauer, J.M.; Braun, C.K.; Hug, S.; Adler, A.; Nilsson Ekdahl, K.; Nilsson, B.; et al. Animal-Free Human Whole Blood Sepsis Model to Study Changes in Innate Immunity. Front. Immunol. 2020, 11, 571992. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- McGeough, M.D.; Pena, C.A.; Mueller, J.L.; Pociask, D.A.; Broderick, L.; Hoffman, H.M.; Brydges, S.D. Cutting edge: IL-6 is a marker of inflammation with no direct role in inflammasome-mediated mouse models. J. Immunol. 2012, 189, 2707–2711. [Google Scholar] [CrossRef] [PubMed]
- Schuttler, J.; Neumann, S. Interleukin-6 as a prognostic marker in dogs in an intensive care unit. Vet. Clin. Pathol. 2015, 44, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Szekely, Y.; Ingbir, M.; Bentur, O.S.; Hochner, O.; Porat, R. Natural cannabinoids suppress the cytokine storm in sepsis-like in vitro model. Eur. Cytokine Netw. 2020, 31, 50–58. [Google Scholar] [CrossRef]
- Vuolo, F.; Petronilho, F.; Sonai, B.; Ritter, C.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Dal-Pizzol, F. Evaluation of Serum Cytokines Levels and the Role of Cannabidiol Treatment in Animal Model of Asthma. Mediators. Inflamm. 2015, 2015, 538670. [Google Scholar] [CrossRef]
- Verrico, C.D.; Wesson, S.; Konduri, V.; Hofferek, C.J.; Vazquez-Perez, J.; Blair, E.; Dunner, K., Jr.; Salimpour, P.; Decker, W.K.; Halpert, M.M. A randomized, double-blind, placebo-controlled study of daily cannabidiol for the treatment of canine osteoarthritis pain. Pain 2020, 161, 2191–2202. [Google Scholar] [CrossRef] [PubMed]
- Joffre, J.; Yeh, C.C.; Wong, E.; Thete, M.; Xu, F.; Zlatanova, I.; Lloyd, E.; Kobzik, L.; Legrand, M.; Hellman, J. Activation of CB1R Promotes Lipopolysaccharide-Induced IL-10 Secretion by Monocytic Myeloid-Derived Suppressive Cells and Reduces Acute Inflammation and Organ Injury. J. Immunol. 2020, 204, 3339–3350. [Google Scholar] [CrossRef] [PubMed]
- Shmarina, G.V.; Pukhalsky, A.L.; Kokarovtseva, S.N.; Pukhalskaya, D.A.; Shabalova, L.A.; Kapranov, N.I.; Kashirskaja, N.J. Tumor necrosis factor-alpha/interleukin-10 balance in normal and cystic fibrosis children. Mediators. Inflamm. 2001, 10, 191–197. [Google Scholar] [CrossRef]
- Winger, E.E.; Reed, J.L.; Ashoush, S.; El-Toukhy, T.; Ahuja, S.; Taranissi, M. Degree of TNF-alpha/IL-10 cytokine elevation correlates with IVF success rates in women undergoing treatment with Adalimumab (Humira) and IVIG. Am. J. Reprod. Immunol. 2011, 65, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.Y.; Argyle, S.A.; Kamida, A.; Morrison, K.O.; Argyle, D.J. The long-acting COX-2 inhibitor mavacoxib (Trocoxil) has anti-proliferative and pro-apoptotic effects on canine cancer cell lines and cancer stem cells in vitro. BMC Vet. Res. 2014, 10, 184. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugliandolo, E.; Licata, P.; Peritore, A.F.; Siracusa, R.; D’Amico, R.; Cordaro, M.; Fusco, R.; Impellizzeri, D.; Di Paola, R.; Cuzzocrea, S.; et al. Effect of Cannabidiol (CBD) on Canine Inflammatory Response: An Ex Vivo Study on LPS Stimulated Whole Blood. Vet. Sci. 2021, 8, 185. https://doi.org/10.3390/vetsci8090185
Gugliandolo E, Licata P, Peritore AF, Siracusa R, D’Amico R, Cordaro M, Fusco R, Impellizzeri D, Di Paola R, Cuzzocrea S, et al. Effect of Cannabidiol (CBD) on Canine Inflammatory Response: An Ex Vivo Study on LPS Stimulated Whole Blood. Veterinary Sciences. 2021; 8(9):185. https://doi.org/10.3390/vetsci8090185
Chicago/Turabian StyleGugliandolo, Enrico, Patrizia Licata, Alessio Filippo Peritore, Rosalba Siracusa, Ramona D’Amico, Marika Cordaro, Roberta Fusco, Daniela Impellizzeri, Rosanna Di Paola, Salvatore Cuzzocrea, and et al. 2021. "Effect of Cannabidiol (CBD) on Canine Inflammatory Response: An Ex Vivo Study on LPS Stimulated Whole Blood" Veterinary Sciences 8, no. 9: 185. https://doi.org/10.3390/vetsci8090185
APA StyleGugliandolo, E., Licata, P., Peritore, A. F., Siracusa, R., D’Amico, R., Cordaro, M., Fusco, R., Impellizzeri, D., Di Paola, R., Cuzzocrea, S., Crupi, R., & Interlandi, C. D. (2021). Effect of Cannabidiol (CBD) on Canine Inflammatory Response: An Ex Vivo Study on LPS Stimulated Whole Blood. Veterinary Sciences, 8(9), 185. https://doi.org/10.3390/vetsci8090185