
The Role of Pathology in the Diagnosis of Swine Respiratory Disease


Abstract
1. Introduction
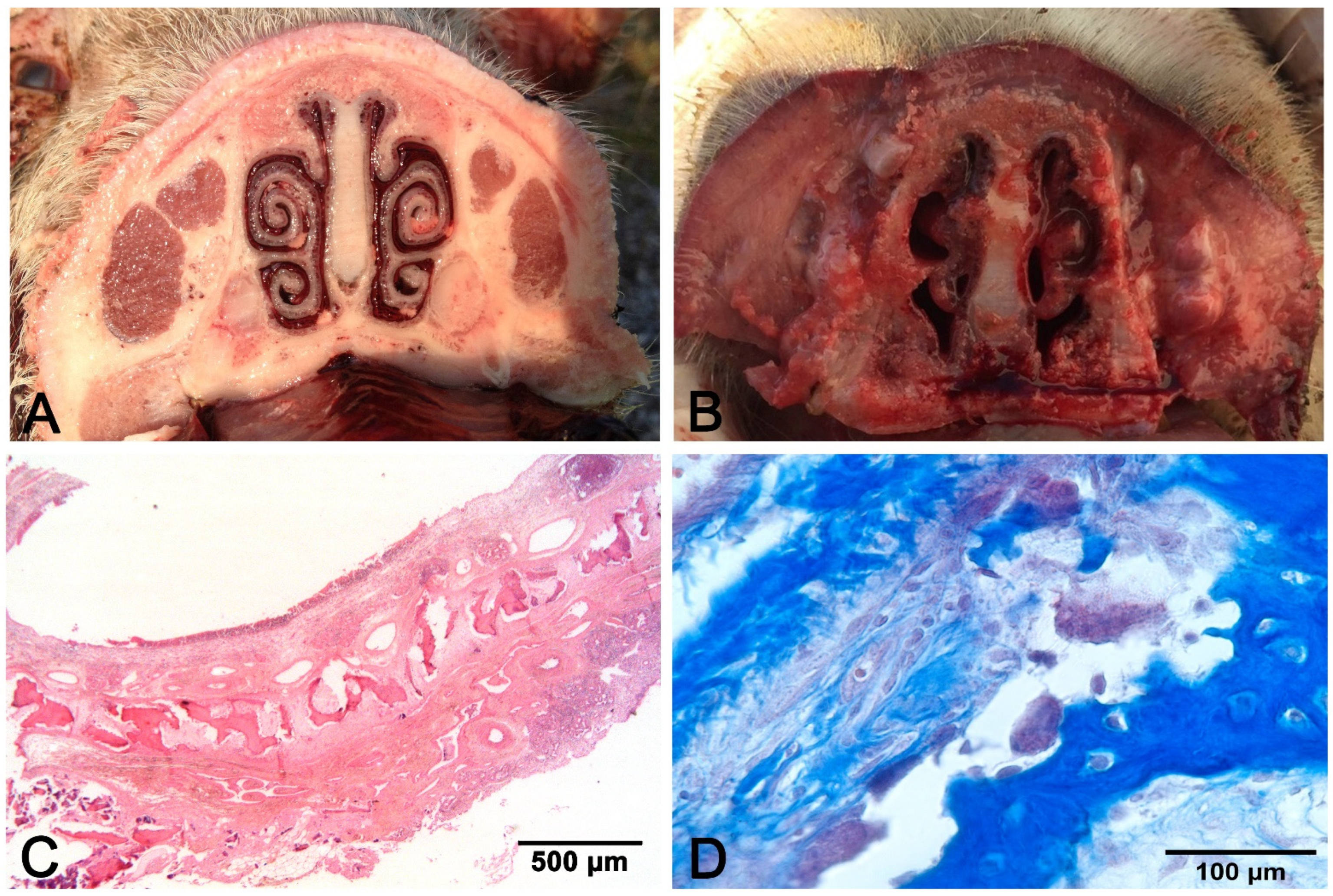
2. Rhinitis
3. Pattern of Pneumonia
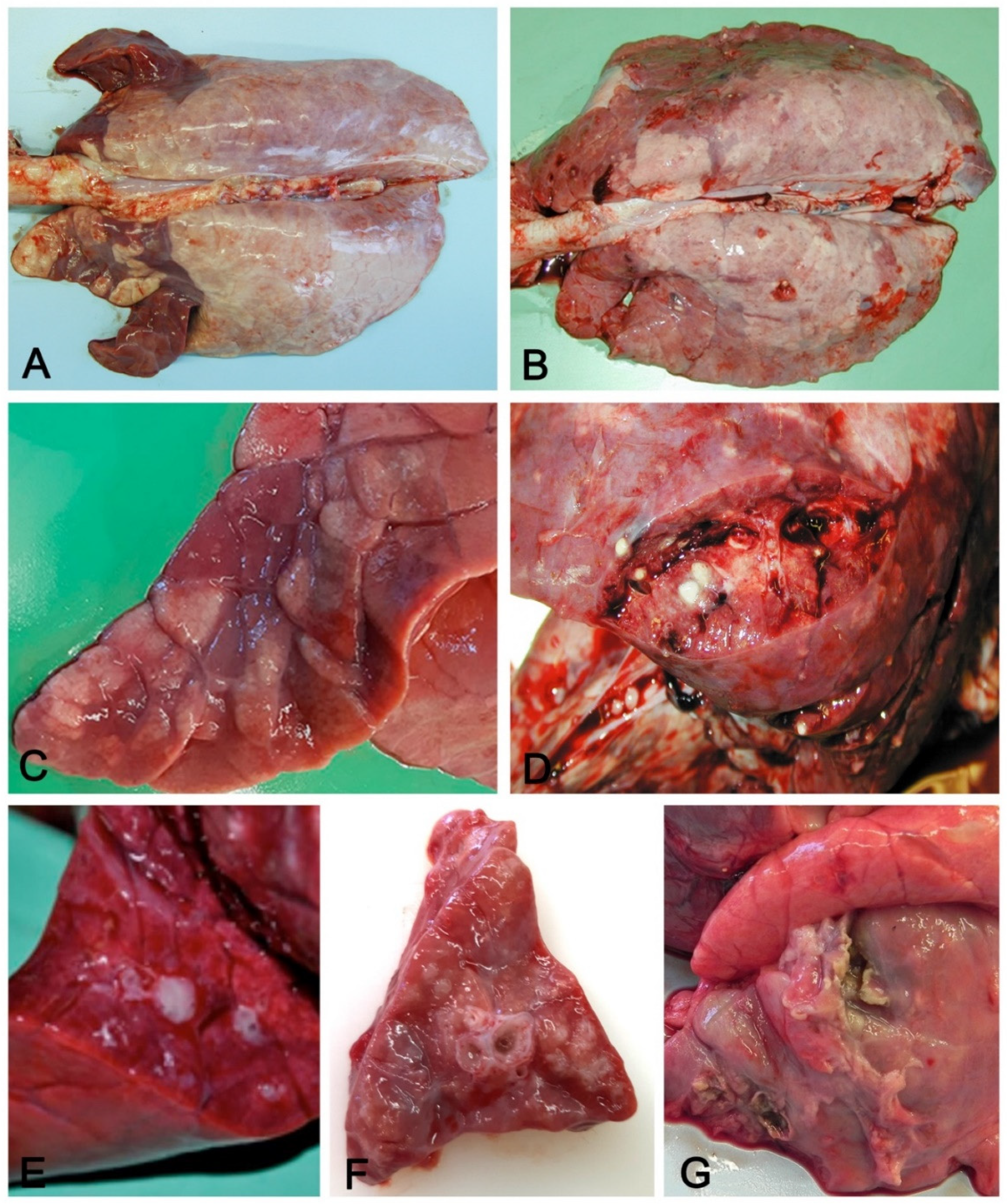
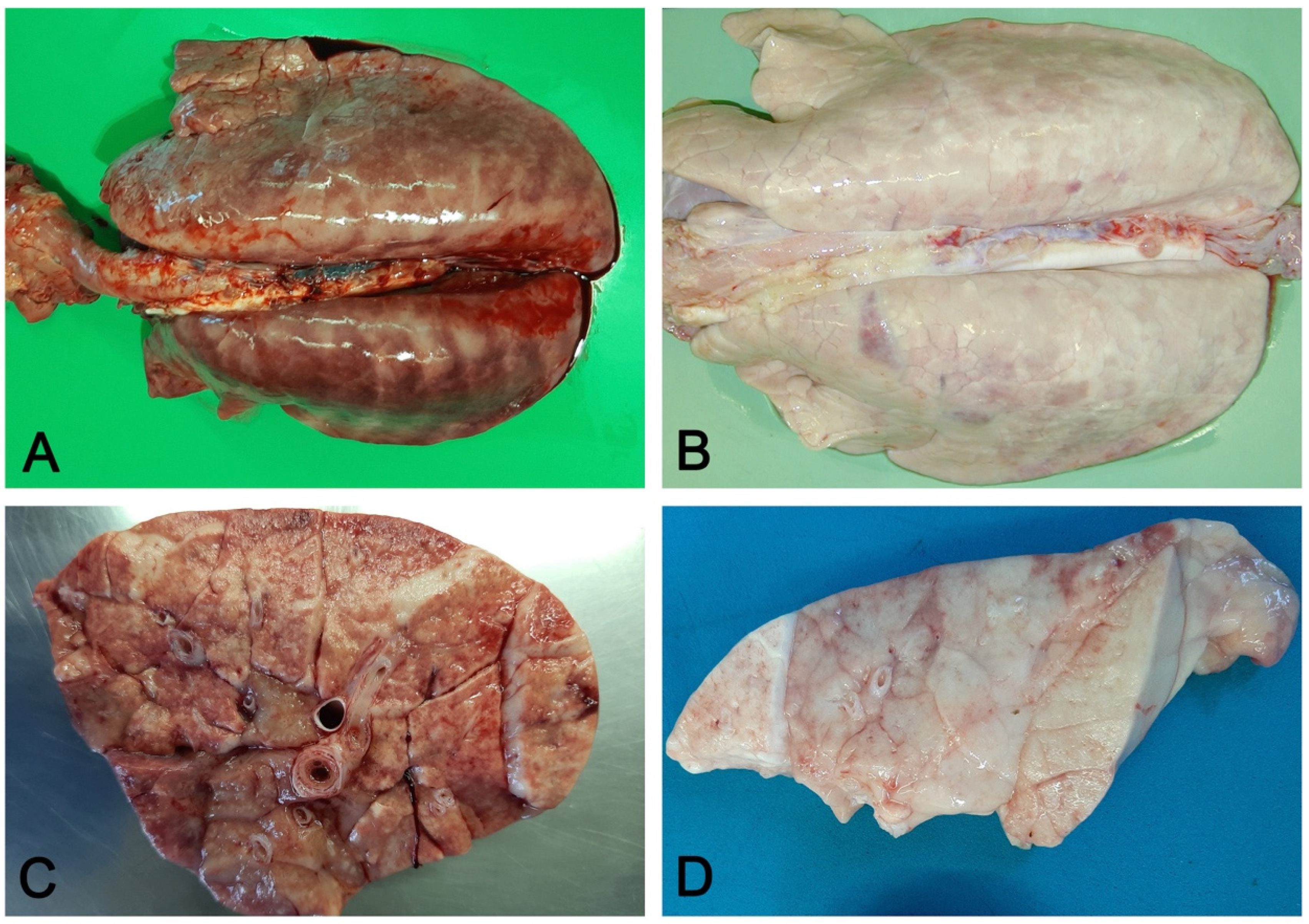
3.1. Macroscopic Pattern of Aerogenous Lung Involvement
- Variation in consistency (from consolidation in acute stages to fibrosis when chronicization occurs);
- Color variation: from dark red in consolidated areas, to a whitish color in chronic lesions. The chromatic variation may also be accentuated by inflammatory edema (acute phase) of the perilobular connective tissue and by its fibrous thickening (chronic phase);
- Presence in the airways, from scarce and dense catarrhal exudate (in EP caused by M. hyopneumoniae) to collection of pus in complicated EP and in bronchopneumonia;
- Whitish mural thickening (cuffing pneumonia) of small airways present in EP.

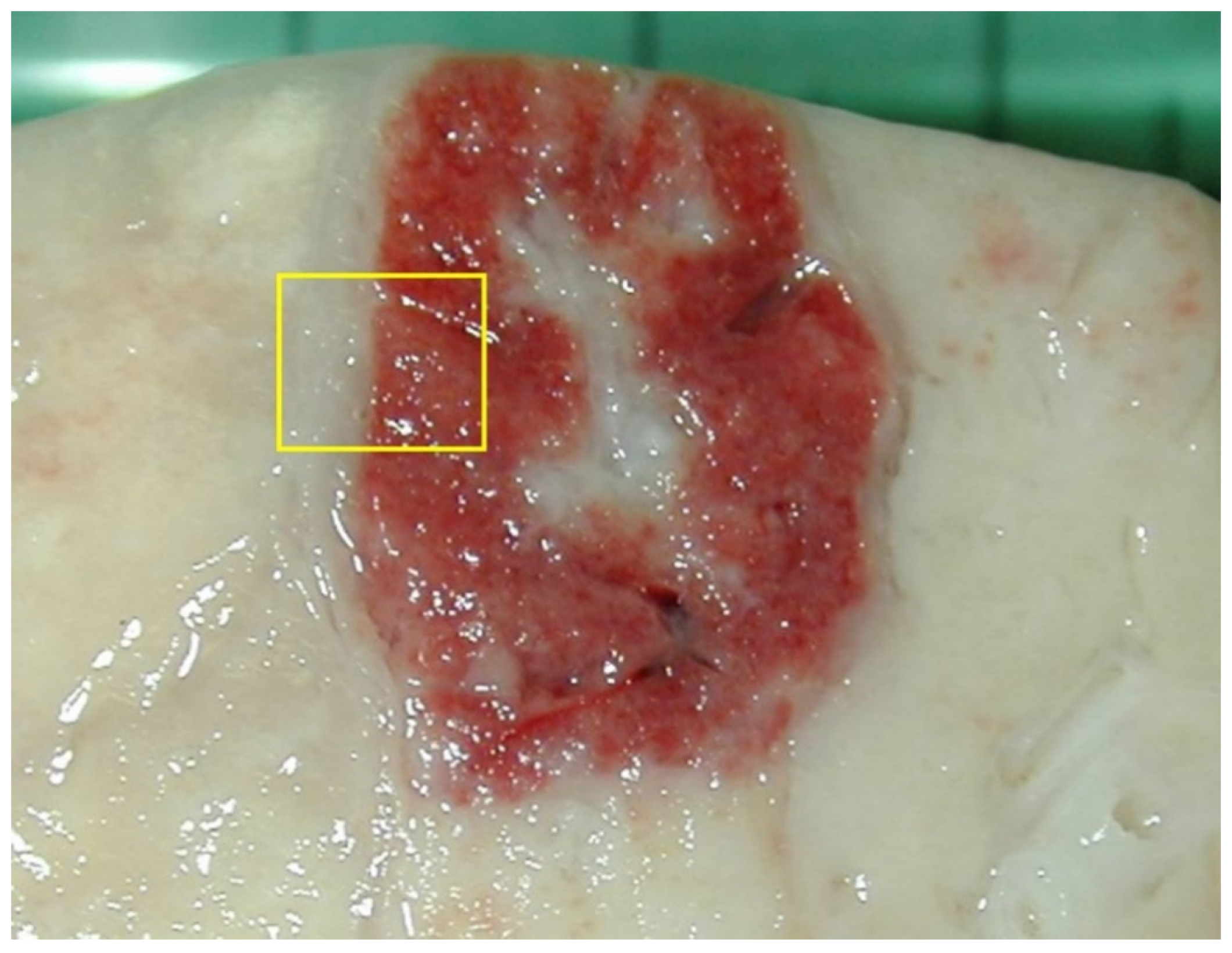
3.2. Macroscopic Pattern of Hematogenous Lung Involvement
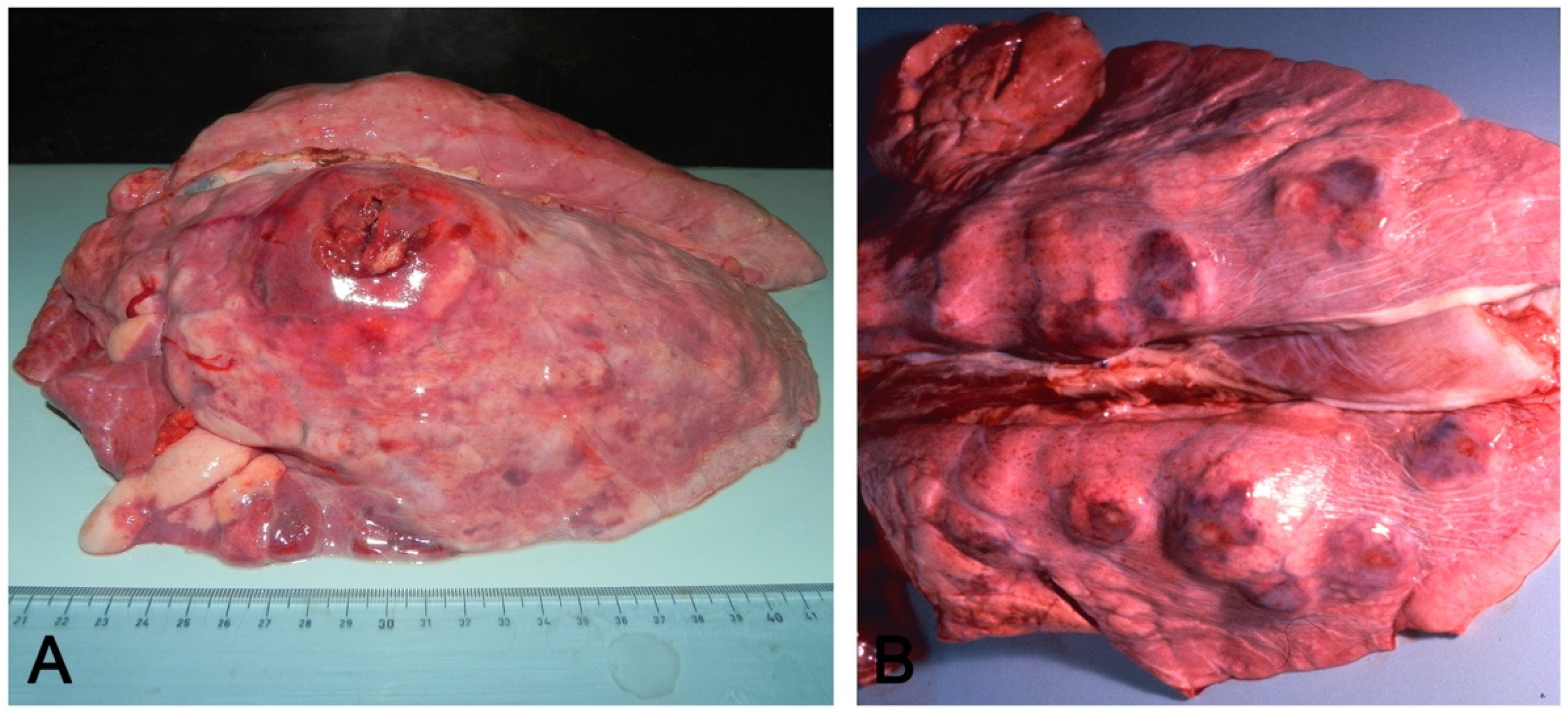
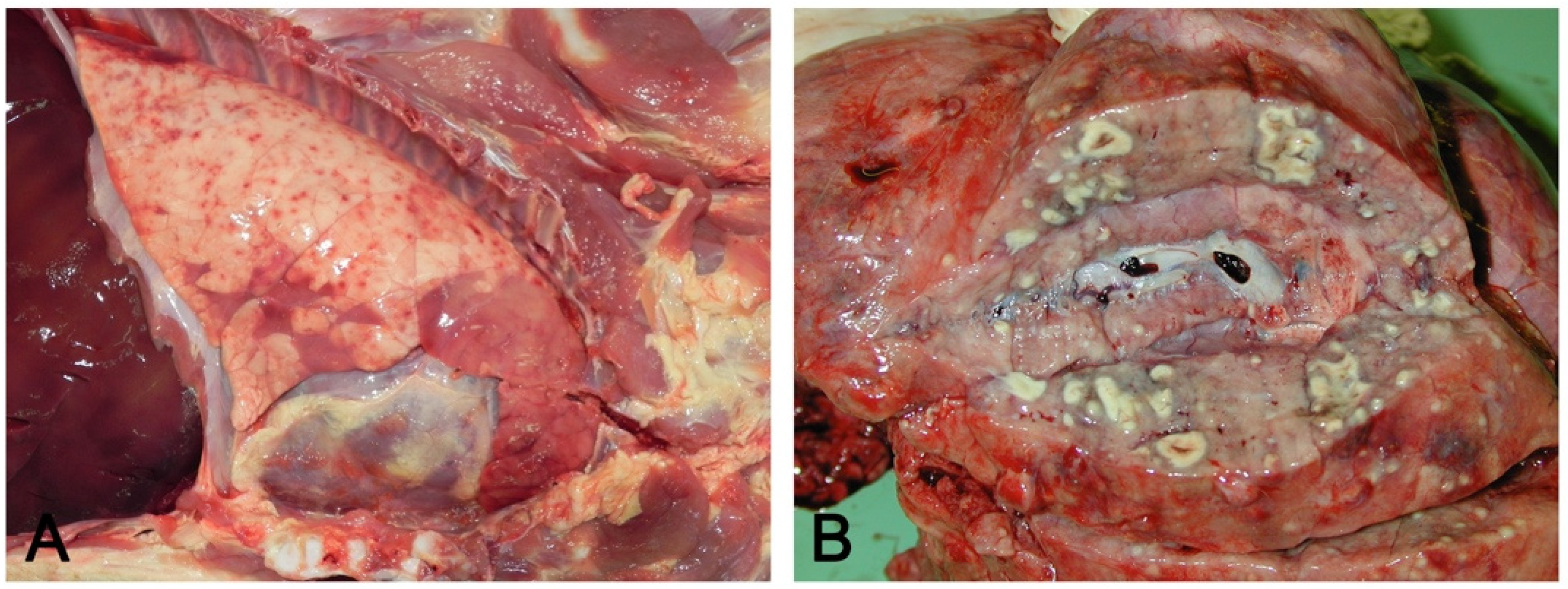
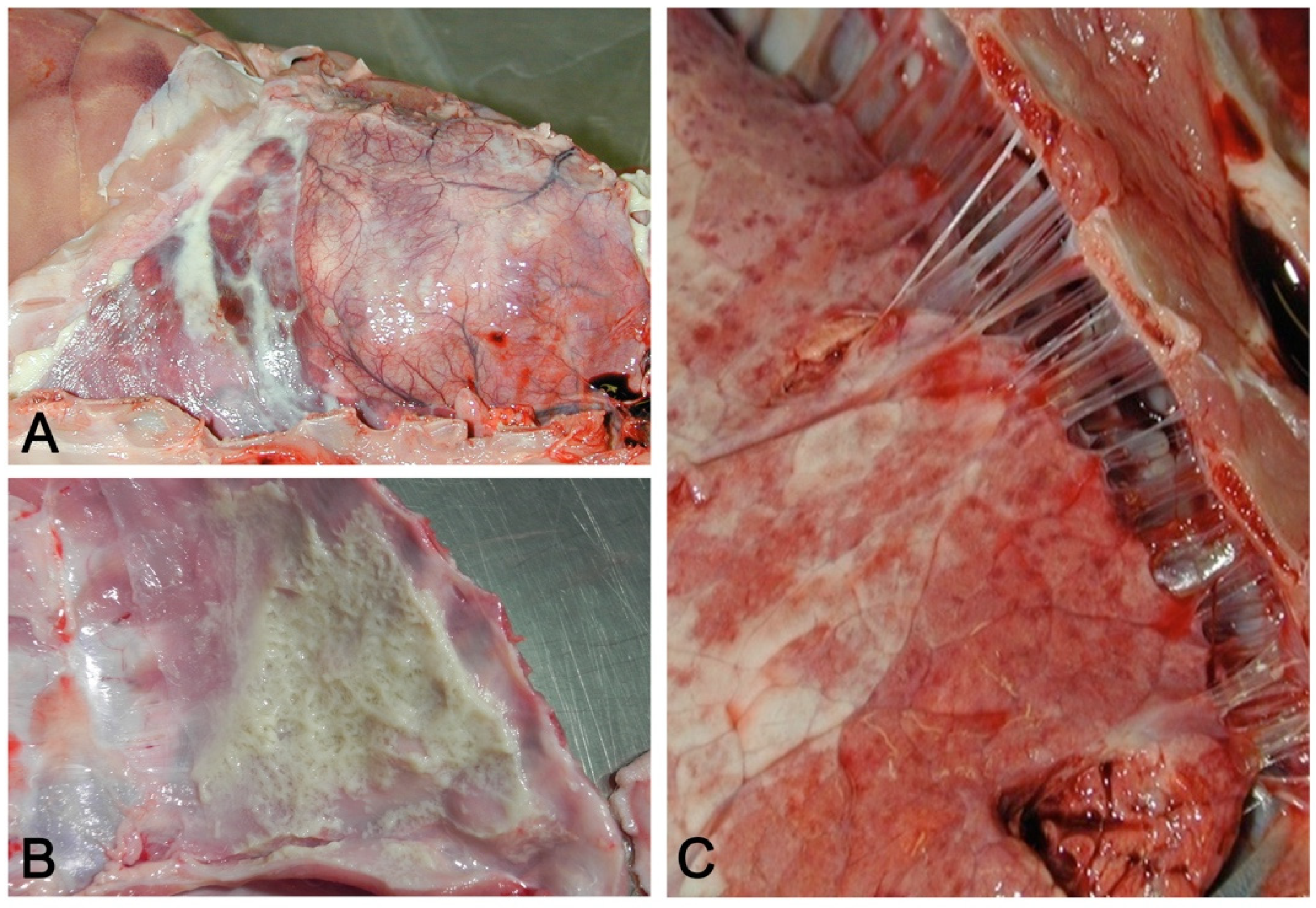
- Embolic pneumonia (arrest in the lung of septic thromboemboli originated from inflammation located elsewhere). Its prominent macroscopic feature is the presence, if recent, of randomly scattered foci of hyperemia/hemorrhage or, if chronic, of abscesses (Figure 4).
- Interstitial pneumonia (arrival of pneumotropic or endotheliotropic agents from primary sites of replication through the blood circulation as in PRRSV and PCV2 infections and septicemia from Gram-negative bacteria). The macroscopic appearance is an interstitial pneumonia characterized by lungs that fail to collapse, with rib impressions, patchy, lobular or diffuse distributions of color variation (ranging from red in the acute stages to a pale whitish color in the chronic forms), changes in consistency (firm texture), interstitial edema (in acute stage), and missing airway involvement (Figure 5).
3.3. Molecular Diagnostic Tools
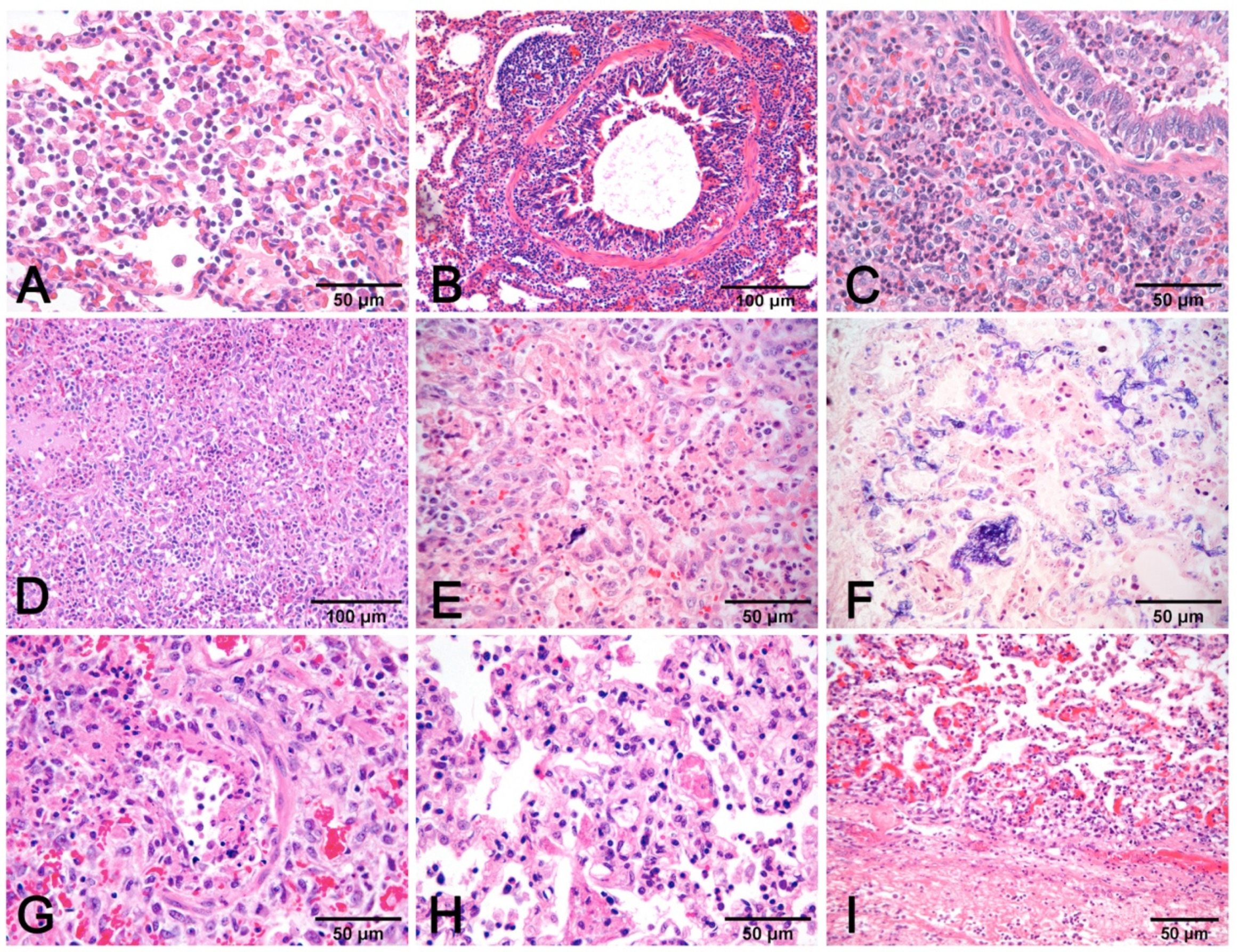
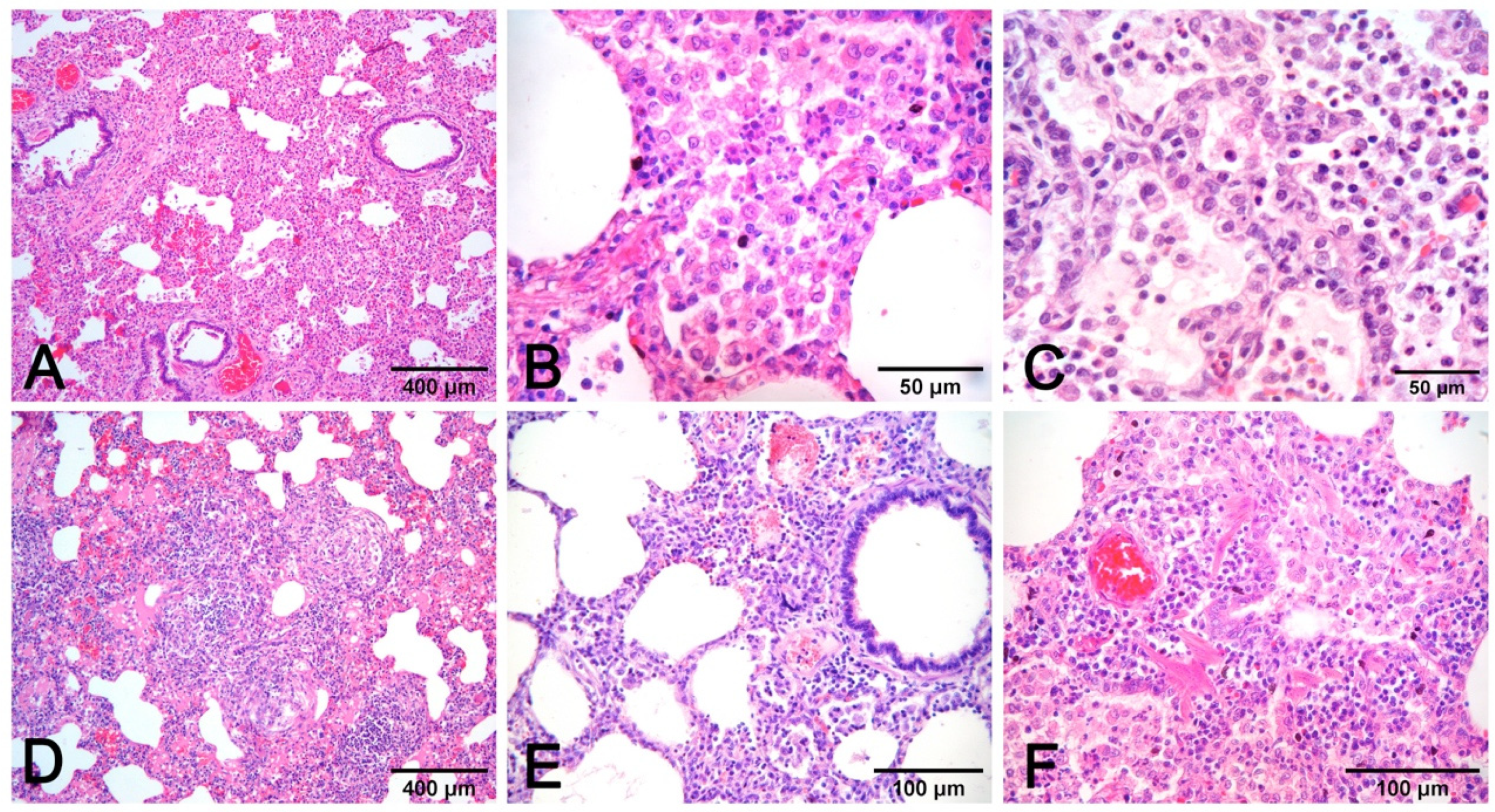
3.4. Histopathology and Lesion–Etiology Colocalization
- Alveolar exudates, mainly represented by edema and macrophages associated with mucus in airways surrounded by lymphocytes to cause peribronchial cuffing, are strongly suggestive of non-complicated EP [19]. The presence of alveolar neutrophils and fibrin is indicative of bacterial complication (Figure 7A–C).
- Necrosis of airway epithelium with cellular debris and leucocytes filling the lumen and broncho-interstitial pneumonia are the hallmarks of SIV infection [19]. Necrosis should also be considered in PRCV infections, where it is limited to small and terminal bronchioles [17], and in PCV2 infections [19] (Figure 8F).
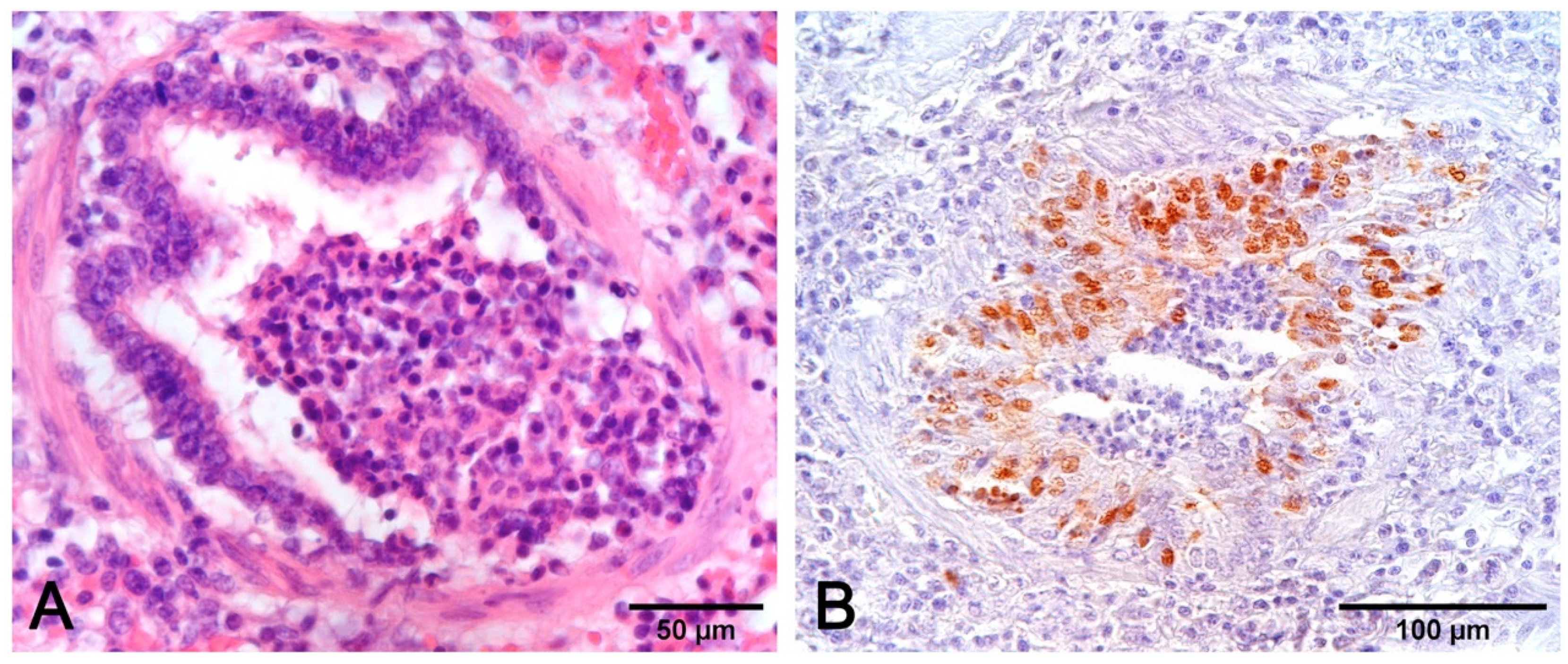
- Fibrinous or necrotizing pneumonia with alveolar necrotic (mainly neutrophils) leucocytes and fibrin collection, associated with vascular thrombosis and fibrinoid mural necrosis, often with concurrent fibrino-hemorrhagic pleuritis, is highly indicative of peracute/acute A. pleuropneumoniae infection (Figure 7D–I). In these cases, the isolation/identification of the etiologic agent is imperative, as the same histologic pattern can be attributed to Actinobacillus suis, septicemic salmonellosis, and pleuritic strains of P. multocida [9,19].
- Interstitial pneumonia, characterized by variable thickening of bronchiolar and alveolar walls, is a hallmark of viral infections:
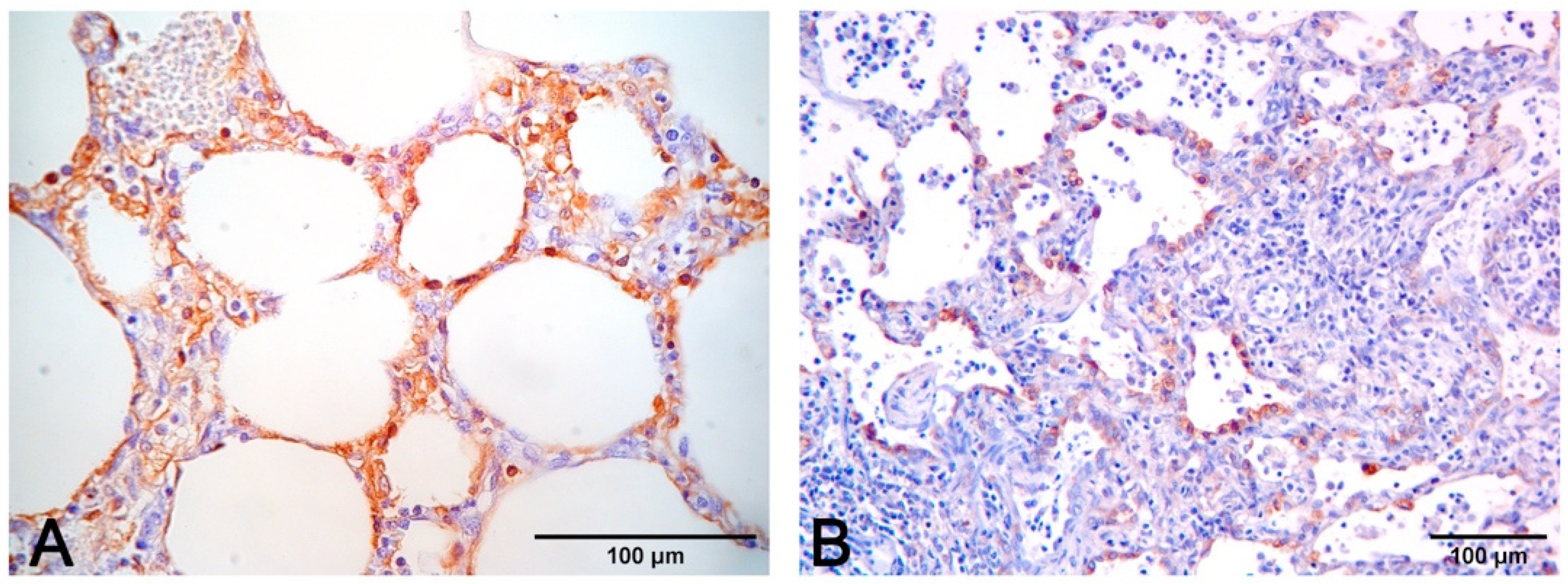
- Pneumonia in PCV2 infection is characterized by peribronchial space and alveolar septa thickened by lymphocytes and macrophages, with a pattern of interstitial to granulomatous pneumonia (Figure 8D,E). Among porcine circovirus diseases (PCVD), lung lesions are reported in both systemic (PCVD-SD; PMWS) and in lung diseases (PCVD-LD); however, when lung lesions are detected, those cases are more often PCVD-SD than PCVD-LD [33]. Other features are bronchiolar necrosis (Figure 8F) progressing to bronchiolitis obliterans and lymphocyte depletion or granulomatous lymphadenitis in regional lymph nodes or other lymphoid tissues [19].
- PRV does not usually induce recognizable microscopic lesions in the lung of older pigs; however, in a few pigs, usually younger, focal necrosis of the parenchyma may be present. Vasculitis, focal cerebral gliosis, and nonsuppurative meningitis in the brain are more consistent lesions [17]. PRV may induce prominent rhinitis.
4. Pleuritis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Opriessnig, T.; Giménez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef]
- Choi, Y.K.; Goyal, S.M.; Joo, H.S. Retrospective analysis of etiologic agents associated with respiratory diseases in pigs. Can. Vet. J. 2003, 44, 735–737. [Google Scholar] [PubMed]
- Fablet, C.; Marois-Créhan, C.; Simon, G.; Grasland, B.; Jestin, A.; Kobisch, M.; Madec, F.; Rose, N. Infectious agents associated with respiratory diseases in 125 far-row-to-finish pig herds: A cross-sectional study. Vet. Microbiol. 2012, 157, 152–163. [Google Scholar] [CrossRef]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine reproductive and respiratory syndrome virus (PRRSV): Pathogenesis and interaction with the immune system. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, S.L.; Halbur, P.G.; Thacker, E.L. Porcine Respiratory Disease Complex. In Polymicrobial Diseases; ASM Press: Washington, DC, USA, 2014; pp. 231–258. [Google Scholar]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-infection of swine with porcine circovirus type 2 and other swine viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Clark, E.; Haines, D.; West, K.; Krakowka, S.; Kennedy, S.; Allan, G.M. Porcine circovirus-2 and concurrent infecions in the field. Vet. Microbiol. 2004, 98, 159–163. [Google Scholar] [CrossRef]
- Vincent, L.L.; Janke, B.H.; Paul, P.S.; Halbur, P.G. A monoclonal-antibody-based immunohistochemical method for the detection of swine influenza virus in formalin-fixed, paraffin-embedded tissues. J. Vet. Diagn. Investig. 1997, 9, 191–195. [Google Scholar] [CrossRef]
- Yaeger, M.J.; Alstine, W.G. Respiratory System. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 393–407. ISBN 9781119350859. [Google Scholar]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; et al. Coinfections and their molecular consequences in the porcine respiratory tract. Vet. Res. 2020, 51, 1–19. [Google Scholar] [CrossRef]
- Jung, K.; Renukaradhya, G.J.; Alekseev, K.P.; Fang, Y.; Tang, Y.; Saif, L.J. Porcine reproductive and respiratory syndrome virus modifies innate immunity and alters disease outcome in pigs subsequently infected with porcine respiratory coronavirus: Implications for respiratory viral co-infections. J. Gen. Virol. 2009, 90, 2713–2723. [Google Scholar] [CrossRef]
- Renukaradhya, G.J.; Alekseev, K.; Jung, K.; Fang, Y.; Saif, L.J. Porcine reproductive and respiratory syndrome virus-induced immunosuppression exacerbates the inflammatory response to porcine respiratory coronavirus in pigs. Viral Immunol. 2010, 23, 457–466. [Google Scholar] [CrossRef]
- Qin, S.; Ruan, W.; Yue, H.; Tang, C.; Zhou, K.; Zhang, B. Viral communities associated with porcine respiratory disease complex in intensive commercial farms in Sichuan province, China. Sci. Rep. 2018, 8, 13341. [Google Scholar] [CrossRef]
- Hansen, M.S.; Pors, S.E.; Jensen, H.E.; Bille-Hansen, V.; Bisgaard, M.; Flachs, E.M.; Nielsen, O.L. An investigation of the pathology and pathogens associated with porcine respiratory disease complex in Denmark. J. Comp. Pathol. 2010, 143, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, M.R.; Segura, M.; Segalés, J.; Gottschalk, M. Review of the speculative role of co-infections in Streptococcus suis-associated diseases in pigs. Vet. Res. 2021, 52, 1–22. [Google Scholar] [CrossRef]
- Pomorska-Mól, M.; Dors, A.; Kwit, K.; Kowalczyk, A.; Stasiak, E.; Pejsak, Z. Kinetics of single and dual infection of pigs with swine influenza virus and Actinobacillus pleuropneumoniae. Vet. Microbiol. 2017, 201, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Janke, B.H. Diagnosis of viral respiratory disease in swine. J. Swine Health Prod. 1995, 3, 116–120. [Google Scholar]
- Sunaga, F.; Tsuchiaka, S.; Kishimoto, M.; Aoki, H.; Kakinoki, M.; Kure, K.; Okumura, H.; Okumura, M.; Okumura, A.; Nagai, M.; et al. Development of a one-run real-time PCR detection system for pathogens associated with porcine respiratory diseases. J. Vet. Med. Sci. 2020, 82, 217–223. [Google Scholar] [CrossRef]
- Caswell, J.L.; Williams, K.J. Respiratory System. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Sixth Edition; Maxie, M.G., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 465–591.e4. [Google Scholar]
- Magyar, T.; Donkó, T.; Repa, I.; Kovács, M. Regeneration of toxigenic Pasteurella multocida induced severe turbinate atrophy in pigs detected by computed tomography. BMC Vet. Res. 2013, 9, 1–7. [Google Scholar] [CrossRef]
- Szeredi, L.; Dán, Á.; Makrai, L.; Takács, N.; Biksi, I. Acute tracheal oedema and haemorrhage with fibrinonecrotic tracheitis in pigs--a porcine counterpart of bovine honker syndrome? J. Comp. Pathol. 2015, 152, 206–210. [Google Scholar] [CrossRef]
- Iowa State University–Influenza. Available online: https://vetmed.iastate.edu/vdpam/FSVD/swine/index-diseases/influenza (accessed on 26 August 2021).
- Ruggeri, J.; Salogni, C.; Giovannini, S.; Vitale, N.; Boniotti, M.B.; Corradi, A.; Pozzi, P.; Pasquali, P.; Alborali, G.L. Association between infectious agents and lesions in post-weaned piglets and fattening heavy pigs with porcine respiratory disease complex (PRDC). Front. Vet. Sci. 2020, 7, 636. [Google Scholar] [CrossRef]
- Lin, K.; Wang, C.; Murtaugh, M.P.; Ramamoorthy, S. Multiplex method for simultaneous serological detection of porcine reproductive and respiratory syndrome virus and porcine circovirus type 2. J. Clin. Microbiol. 2011, 49, 3184–3190. [Google Scholar] [CrossRef]
- Christopher-Hennings, J.; Erickson, G.A.; Hesse, R.A.; Nelson, E.A.; Rossow, S.; Scaria, J.; Slavic, D. Diagnostic Tests, Test Performance, and Considerations for Interpretation. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 75–97. ISBN 9781119350859. [Google Scholar]
- Trang, N.T.; Hirai, T.; Yamamoto, T.; Matsuda, M.; Okumura, N.; Giang, N.T.H.; Lan, N.T.; Yamaguchi, R. Detection of porcine reproductive and respiratory syndrome virus in oral fluid from naturally infected pigs in a breeding herd. J. Vet. Sci. 2014, 15, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Trang, N.T.; Ngan, P.H.; Hop, N.; Van Hirai, T. Nasal swab–a new tool for the detection of porcine respiratory disease complex in natural infected pigs. Acta Sci. Microbiol. 2018, 1, 2–5. [Google Scholar]
- Hernandez-Garcia, J.; Robben, N.; Magnée, D.; Eley, T.; Dennis, I.; Kayes, S.M.; Thomson, J.R.; Tucker, A.W. The use of oral fluids to monitor key pathogens in porcine respiratory disease complex. Porc. Health Manag. 2017, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lung, O.; Ohene-Adjei, S.; Buchanan, C.; Joseph, T.; King, R.; Erickson, A.; Detmer, S.; Ambagala, A. Multiplex PCR and microarray for detection of swine respiratory pathogens. Transbound. Emerg. Dis. 2017, 64, 834–848. [Google Scholar] [CrossRef]
- Maes, R.K.; Langohr, I.M.; Wise, A.G.; Smedley, R.C.; Thaiwong, T.; Kiupel, M. Beyond H&E: Integration of nucleic acid–based analyses into diagnostic pathology. Vet. Pathol. 2014, 51, 238–256. [Google Scholar] [CrossRef]
- Giles, T.A.; Belkhiri, A.; Barrow, P.A.; Foster, N. Molecular approaches to the diagnosis and monitoring of production diseases in pigs. Res. Vet. Sci. 2017, 114, 266–272. [Google Scholar] [CrossRef]
- Gupta, E.; Bhalla, P.; Khurana, N.; Singh, T. Histopathology for the diagnosis of infectious diseases. Indian J. Med. Microbiol. 2009, 27, 100–106. [Google Scholar] [CrossRef]
- Ticó, G.; Segalés, J.; Martínez, J. The blurred border between porcine circovirus type 2-systemic disease and porcine respirtory disease complex. Vet. Microbiol. 2013, 163, 242–247. [Google Scholar] [CrossRef]
- Morandi, F.; Ostanello, F.; Fusaro, L.; Bacci, B.; Nigrelli, A.; Alborali, L.; Dottori, M.; Vezzoli, F.; Barigazzi, G.; Fiorentini, L.; et al. Immunohistochemical detection of aetiological agents of proliferative and necrotizing pneumonia in italian pigs. J. Comp. Pathol. 2010, 142, 74–78. [Google Scholar] [CrossRef][Green Version]
- Grau-Roma, L.; Segalés, J. Detection of porcine reproductive and respiratory syndrome virus, porcine circovirus type 2, swine influenza virus and Aujeszky’s disease virus in cases of porcine proliferative and necrotizing pneumonia (PNP) in Spain. Vet. Microbiol. 2007, 119, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Morin, M.; Girard, C.; Elazhary, Y.; Fajardo, R.; Drolet, R.L. Severe proliferative and necrotizing pneumonia in pigs: A newly recognized disease. Can. Vet. J. 1990, 31, 837. [Google Scholar] [PubMed]
- Dea, S.; Bilodeau, R.; Sauvageau, R.; Montpetit, C.; Martineau, G.P. Antigenic variant of swine influenza virus causing Proliferative and Necrotizing Pneumonia in pigs. J. Vet. Diagn. Investig. 1992, 4, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Guedes, R.M.C.; Gebhart, C.J.; Winkelman, N.L.; Mackie-Nuss, R.A.C.; Marsteller, T.A.; Deen, J. Comparison of different methods for diagnosis of porcine proliferative enteropathy. Can. J. Vet. Res. 2002, 66, 99–107. [Google Scholar] [PubMed]
- Driemeier, D.; Faccini, G.S.; de Oliveira, R.T.; Colodel, E.M.; Traverso, S.D.; Cattani, C. Silver staining combined with alcian blue and hematoxylin-eosin for the detection of Lawsonia intracelullaris in swine proliferative enteropathy. Acta Histochem. 2002, 104, 285–287. [Google Scholar] [CrossRef]
- Woods, G.L.; Walker, D.H. Detection of infection or infectious agents by use of cytologic and histologic stains. Clin. Microbiol. Rev. 1996, 9, 382–404. [Google Scholar] [CrossRef]
- Haines, D.M.; West, K.H. Immunohistochemistry: Forging the links between immunology and pathology. Vet. Immunol. Immunopathol. 2005, 108, 151–156. [Google Scholar] [CrossRef]
- Ellis, J.; Hassard, L.; Clark, E.; Harding, J.; Allan, G.; Willson, P.; Strokappe, J.; Martin, K.; McNeilly, F.; Meehan, B.; et al. Isolation of circovirus from lesions of pigs with postweaning multisystemic wasting syndrome. Can. Vet. J. 1998, 39, 44–51. [Google Scholar]
- West, K.H.; Bystrom, J.M.; Wojnarowicz, C.; Shantz, N.; Jacobson, M.; Allan, G.M.; Haines, D.M.; Clark, E.G.; Krakowka, S.; McNeilly, F.; et al. Myocarditis and abortion associated with intrauterine infection of sows with porcine circovirus 2. J. Vet. Diagn. Investig. 1999, 11, 530–532. [Google Scholar] [CrossRef]
- Halbur, P.G.; Andrews, J.J.; Huffman, E.L.; Paul, P.S.; Meng, X.J.; Niyo, Y. De-velopment of a streptavidin-biotin immunoperoxidase procedure for the detection of porcine reproductive and respiratory syndrome virus antigen in porcine lung. J. Vet. Diagn. Investig. 1994, 6, 254–257. [Google Scholar] [CrossRef]
- Kim, D.; Ha, Y.; Lee, Y.-H.; Chae, S.; Lee, K.; Han, K.; Kim, J.; Lee, J.-H.; Kim, S.-H.; Hwang, K.-K.; et al. Comparative study of in situ hybridization and immunohistochemistry for the detection of porcine circovirus 2 in formalin-fixed, paraffin-embedded tissues. J. Vet. Med. Sci. 2009, 71, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A. Immunohistochemistry: A rapid and specific diagnostic tool for influenza cirus infection in pigs. J. Dairy Vet. Anim. Res. 2017, 5, 2016–2018. [Google Scholar] [CrossRef]
- Hansen, M.S.; Pors, S.E.; Bille-Hansen, V.; Kjerulff, S.K.J.; Nielsen, O.L. Occurrence and tissue distribution of porcine circovirus type 2 identified by immunohistochemistry in danish finishing pigs at slaughter. J. Comp. Pathol. 2010, 142, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Ober, R.A.; Thissen, J.B.; Jaing, C.J.; Cino-Ozuna, A.G.; Rowland, R.R.R.; Nie-derwerder, M.C. Increased microbiome diversity at the time of infection is associated with improved growth rates of pigs after co-infection with porcine reproductive and respiratory syndrome virus (PRRSV) and porcine circovirus type 2 (PCV2). Vet. Microbiol. 2017, 208, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Thacker, E.L.; Yu, S.; Fenaux, M.; Meng, X.J.; Halbur, P.G. Experimental reproduction of postweaning multisystemic wasting syndrome in pigs by dual infection with Mycoplasma hyopneumoniae and porcine circovirus type 2. Vet. Pathol. 2004, 41, 624–640. [Google Scholar] [CrossRef]
- Park, C.; Seo, H.W.; Park, S.J.; Han, K.; Chae, C. Comparison of porcine circovirus type 2 (PCV2)-associated lesions prouced by co-infection between two genotypes of PCV2 and two genotypes of porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2014, 95, 2486–2494. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Opriessnig, T.; Pal, N.; Huang, F.F.; Meng, X.J. Effect of an interferon-stimulated response element (ISRE) mutant of porcine circovirus type 2 (PCV2) on PCV2-induced pathological lesions in a porcine reproductive and respiratory syn-drome virus (PRRSV) co-infection model. Vet. Microbiol. 2011, 147, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J. Porcine circovirus type 2 (PCV2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Opriessnig, T.; Meng, X.-J.; Halbur, P.G. Porcine Circovirus Type 2–Associated Disease: Update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J. Vet. Diagn. Investig. 2007, 19, 591–615. [Google Scholar] [CrossRef]
- Segalés, J. Pathological findings associated with naturally acquired porcine circovirus type 2 associated disease. Vet. Microbiol. 2004, 98, 137–149. [Google Scholar] [CrossRef]
- Segalés, J.; Allan, G.M.; Domingo, M. Circoviruses. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 473–487. [Google Scholar]
- Han, K.; Seo, H.W.; Oh, Y.; Kang, I.; Park, C.; Kang, S.H.; Kim, S.-H.; Lee, B.-H.; Kwon, B.; Chae, C. Evaluation of monoclonal antibody–based immunohistochemistry for the detection of European and North American Porcine reproductive and respiratory syndrome virus and a comparison with in situ hybridization and reverse transcription polymerase chain reaction. J. Vet. Diagn. Investig. 2012, 24, 719–724. [Google Scholar] [CrossRef]
- Zimmerman, J.J.; Dee, S.A.; Holtkamp, D.J.; Murtaugh, M.P.; Stadejek, T.; Stevenson, G.W.; Torremorell, M.; Yang, H.; Zhang, J. Porcine Reproductive and Respiratory Syndrome Viruses (Porcine Arteriviruses). In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 685–708. [Google Scholar]
- Zhang, Q.; Bai, J.; Hou, H.; Song, Z.; Zhao, Y.; Jiang, P. A novel recombinant porcine reproductive and respiratory syndrome virus with significant variation in cell adaption and pathogenicity. Vet. Microbiol. 2017, 208, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Kedkovid, R.; Woonwong, Y.; Arunorat, J.; Sirisereewan, C.; Sangpratum, N.; Lumyai, M.; Kesdangsakonwut, S.; Teankum, K.; Jittimanee, S.; Thanawongnuwech, R. Porcine circovirus type 3 (PCV3) infection in grower pigs from a Thai farm suffering from porcine respiratory disease complex (PRDC). Vet. Microbiol. 2018, 215, 71–76. [Google Scholar] [CrossRef]
- Kim, S.-H.; Park, J.-Y.; Jung, J.; Kim, H.; Park, Y.-R.; Lee, K.; Lyoo, Y.S.; Yeo, S.; Park, C. Detection and genetic characterization of porcine circovirus 3 from aborted fetuses and pigs with respiratory disease in Korea. J. Vet. Sci. 2018, 19, 721. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.E.R.; Vannucci, F.A.; Gabardo, M.d.P.; dos Santos, L.F.; Mores, N.; Guedes, R.M.C. Mycoplasma hyorhinis infection in early cases of mycoplasmal pneumonia in swine and evaluation of diagnostic assays. Pesqui. Vet. Bras. 2017, 37, 1057–1063. [Google Scholar] [CrossRef]
- Hoeltig, D.; Rohde, J.; Frase, R.; Nietfeld, F.; Waldmann, K.H.; Valentin-Weigand, P.; Meens, J. Multi-organ spreading of Actinobacillus pleuropneumoniae serovar 7 in weaned pigs during the first week after experimental infection. Vet. Res. 2018, 49, 1–11. [Google Scholar] [CrossRef]
- Hennig-Pauka, I.; Koch, R.; Hoeltig, D.; Gerlach, G.F.; Waldmann, K.H.; Blecha, F.; Brauer, C.; Gasse, H. PR-39, a porcine host defence peptide, is prominent in mucosa and lymphatic tissue of the respiratory tract in healthy pigs and pigs infected with actino-bacillus pleuropneumoniae. BMC Res. Notes 2012, 5, 1–7. [Google Scholar] [CrossRef]
- Park, S.J.; Seo, H.W.; Park, C.; Chae, C. Interaction between single-dose Myco-plasma hyopneumoniae and porcine reproductive and respiratory syndrome virus vaccines on dually infected pigs. Res. Vet. Sci. 2014, 96, 516–522. [Google Scholar] [CrossRef]
- Bai, X.; Wang, Y.; Xu, X.; Sun, Z.; Xiao, Y.; Ji, G.; Li, Y.; Tan, F.; Li, X.; Tian, K. Commercial vaccines provide limited protection to NADC30-like PRRSV infection. Vaccine 2016, 34, 5540–5545. [Google Scholar] [CrossRef]
- Evenson, D.; Gerber, P.F.; Xiao, C.T.; Halbur, P.G.; Wang, C.; Tian, D.; Ni, Y.Y.; Meng, X.J.; Opriessnig, T. A porcine reproductive and respiratory syndrome virus candidate vaccine based on the synthetic attenuated virus engineering approach is at-tenuated and effective in protecting against homologous virus challenge. Vaccine 2016, 34, 5546–5553. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Xu, H.; Wang, Y.; Liu, Z.; He, F. Efficacy evaluation of two commercial vaccines against a recombinant PRRSV2 strain ZJnb16-2 from lineage 8 and 3 in china. Pathogens 2020, 9, 59. [Google Scholar] [CrossRef]
- Oliver-Ferrando, S.; Segalés, J.; López-Soria, S.; Callén, A.; Merdy, O.; Joisel, F.; Sibila, M. Exploratory field study on the effect of Porcine circovirus 2 (PCV2) sow vaccination on serological, virological and reproductive parameters in a PCV2 subclinically infected sow herd. BMC Vet. Res. 2018, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Castro, A.M.M.G.; Karuppanan, A.K.; Gauger, P.C.; Halbur, P.G.; Matzinger, S.R.; Meng, X.J. A Porcine circovirus type 2b (PCV2b)-based experimental vaccine is effective in the PCV2b-Mycoplasma hyopneumoniae coinfection pig model. Vaccine 2019, 37, 6688–6695. [Google Scholar] [CrossRef] [PubMed]
- Yaeger, M.J. The diagnostic sensitivity of immunohistochemistry for the detection of porcine reproductive and respiratory syndrome virus in the lung of vaccinated and unvaccinated swine. J. Vet. Diagn. Investig. 2002, 14, 15–19. [Google Scholar] [CrossRef]
- Guarneri, F.; Tresoldi, E.T.; Sarli, G.; Boniotti, M.B.; Lelli, D.; Barbieri, I.; Bacci, B.; D’Annunzio, G.; Amadori, M. Protective immunity in swine induced by Porcine Circovirus 2b inactivated vaccines with different antigen payload. Vet. Microbiol. 2021, 252, 108887. [Google Scholar] [CrossRef] [PubMed]
- Merialdi, G.; Dottori, M.; Bonilauri, P.; Luppi, A.; Gozio, S.; Pozzi, P.; Spaggiari, B.; Martelli, P. Survey of pleuritis and pulmonary lesions in pigs at abattoir with a focus on the extent of the condition and herd risk factors. Vet. J. 2012, 193, 234–239. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Pathogens | Secondary Pathogens |
|---|---|
| PRRSV * SIV * PRCV PCV2 * Mycoplasma hyopneumoniae Actinobacillus pleuropneumoniae | Mycoplasma spp. Streptococcus spp. Staphylococcus spp. Escherichia coli Klebsiella spp. Trueperella pyogenes Bordetella bronchiseptica Glaesserella parasuis Pasteurella multocida |
| Suspected Etiology/Disease a | Tissue/Sample | Fresh/Fixed (10% Buffered Formalin) | |
|---|---|---|---|
| Systemic tropism e.g., PCV2 PRRSV PRV M. hyorinis | Lung | From affected areas with different gross appearance (cranial, middle lobes and the cranial portion of the caudal lobe with visible airways) | |
| Lymph nodes | Mandibular, sternal, tracheobronchial, mesenteric and superficial inguinal | ||
| Tonsil | One or both sides | ||
| Spleen | Representative portion or in case of absence of lesions elsewhere | ||
| Heart (PCV2; M. hyorinis) | Left and right ventricles and septum | ||
| Brain (PRV) | Representative portion or in case of absence of lesions elsewhere | ||
| Respiratory tropism e.g., NPAR and PAR M. hyopneumoniae A. pleuropneumoniae | Upper respiratory tract | Nasal turbinate | 1 cm thickness |
| Low respiratory tract | Lung | From affected areas with different gross appearance (cranial, middle lobes and the cranial portion of the caudal lobe with visible airways) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarli, G.; D’Annunzio, G.; Gobbo, F.; Benazzi, C.; Ostanello, F. The Role of Pathology in the Diagnosis of Swine Respiratory Disease. Vet. Sci. 2021, 8, 256. https://doi.org/10.3390/vetsci8110256
Sarli G, D’Annunzio G, Gobbo F, Benazzi C, Ostanello F. The Role of Pathology in the Diagnosis of Swine Respiratory Disease. Veterinary Sciences. 2021; 8(11):256. https://doi.org/10.3390/vetsci8110256
Chicago/Turabian StyleSarli, Giuseppe, Giulia D’Annunzio, Francesca Gobbo, Cinzia Benazzi, and Fabio Ostanello. 2021. "The Role of Pathology in the Diagnosis of Swine Respiratory Disease" Veterinary Sciences 8, no. 11: 256. https://doi.org/10.3390/vetsci8110256
APA StyleSarli, G., D’Annunzio, G., Gobbo, F., Benazzi, C., & Ostanello, F. (2021). The Role of Pathology in the Diagnosis of Swine Respiratory Disease. Veterinary Sciences, 8(11), 256. https://doi.org/10.3390/vetsci8110256