
Teladorsagia Circumcincta Galectin-Mucosal Interactome in Sheep

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sheep Tissue
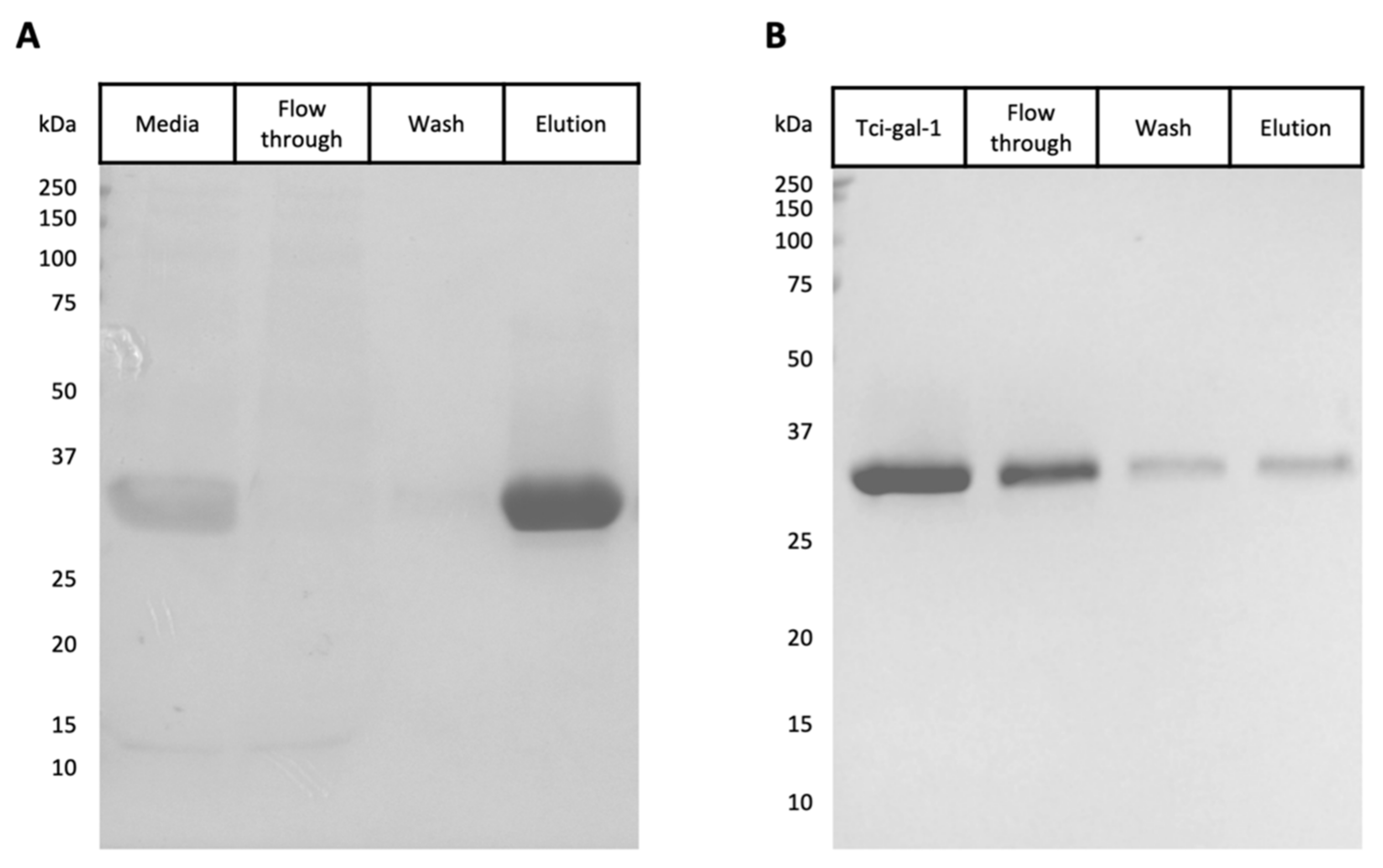
2.2. Tci-gal-1 Expression and Purification
2.3. Determination of Lactose Binding Affinity
2.4. Conjugation of Tci-gal-1 Onto N-Hydroxysuccinamide (NHS)-Activated Sepharose
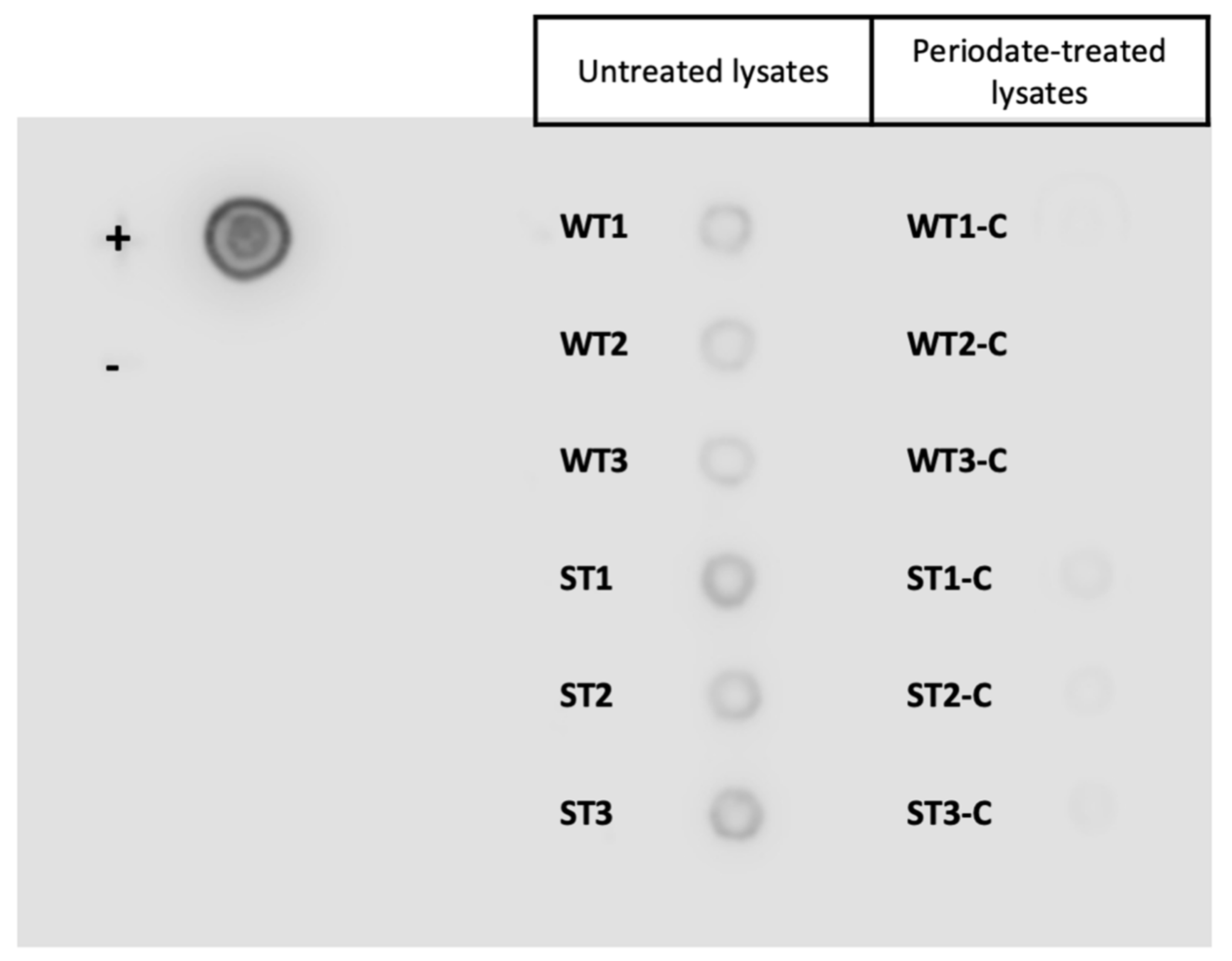
2.5. Sodium Periodate Treatment
2.6. Isolation of Tci-gal-1-Binding Ligands from Sheep Abomasal Tissue
2.7. Mass Spectrometry
2.8. Protein Identification and Quantification
2.9. Protein Annotation and Glycosylation Analysis
3. Results
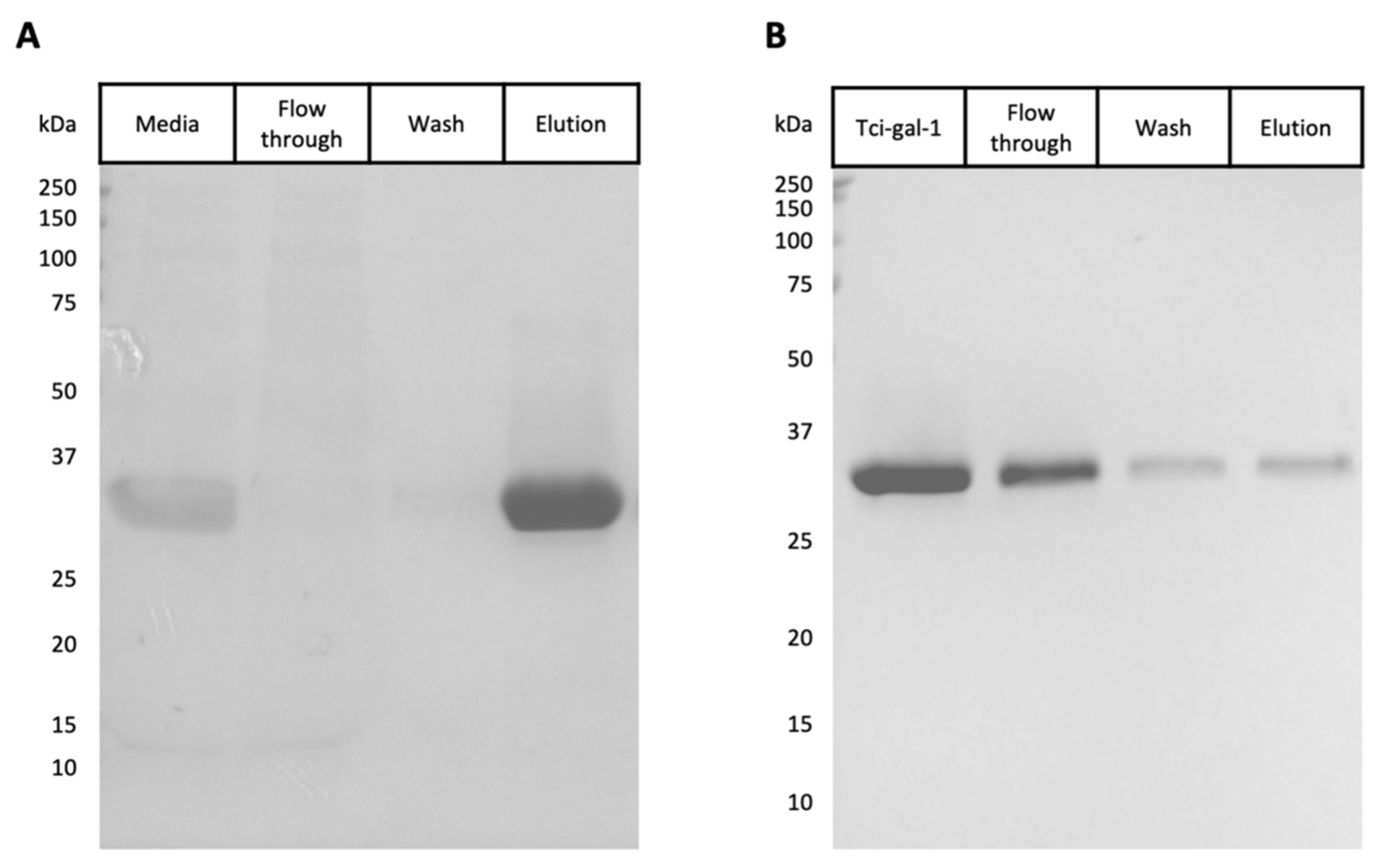
3.1. Expression and Purification of Tci-gal-1
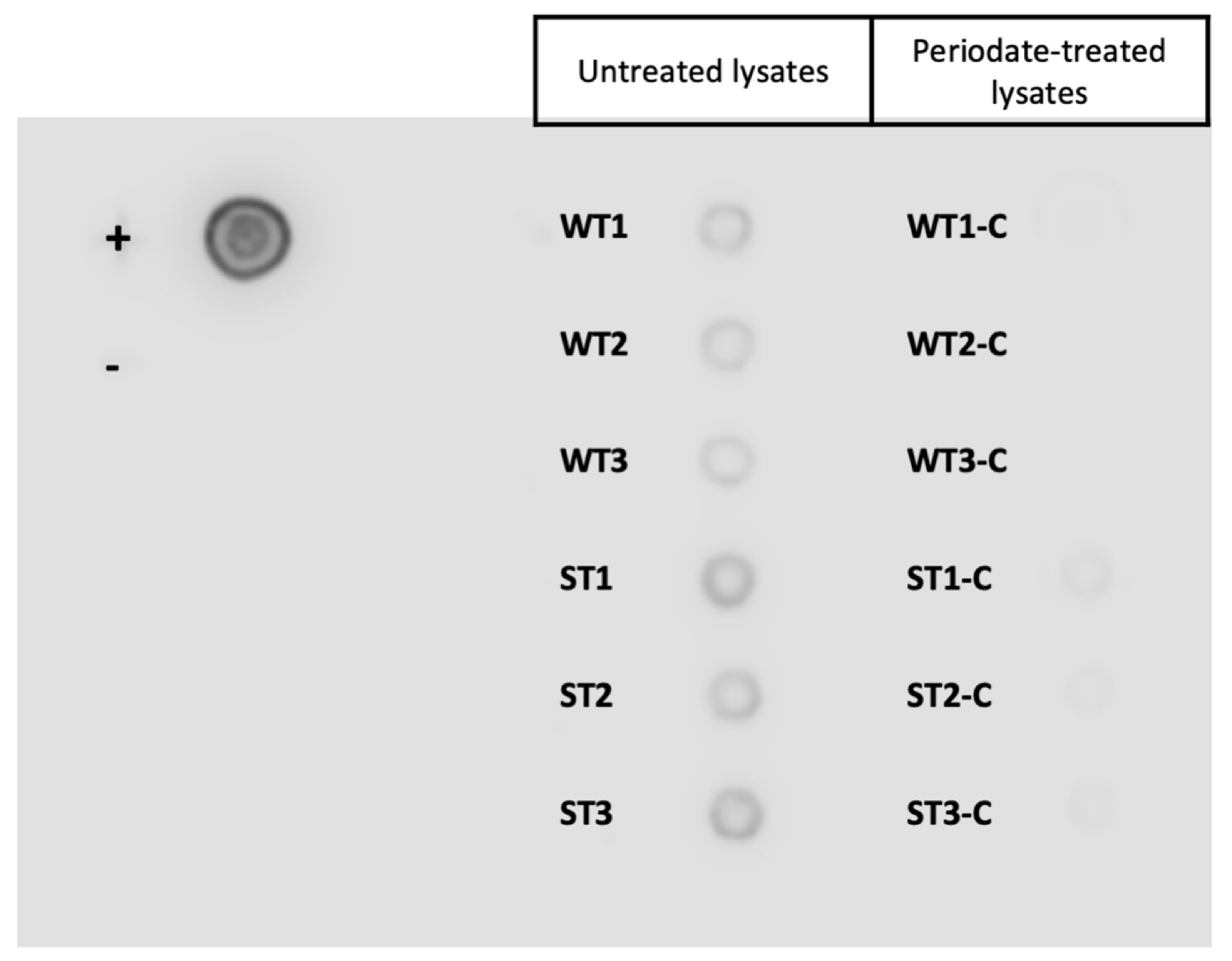
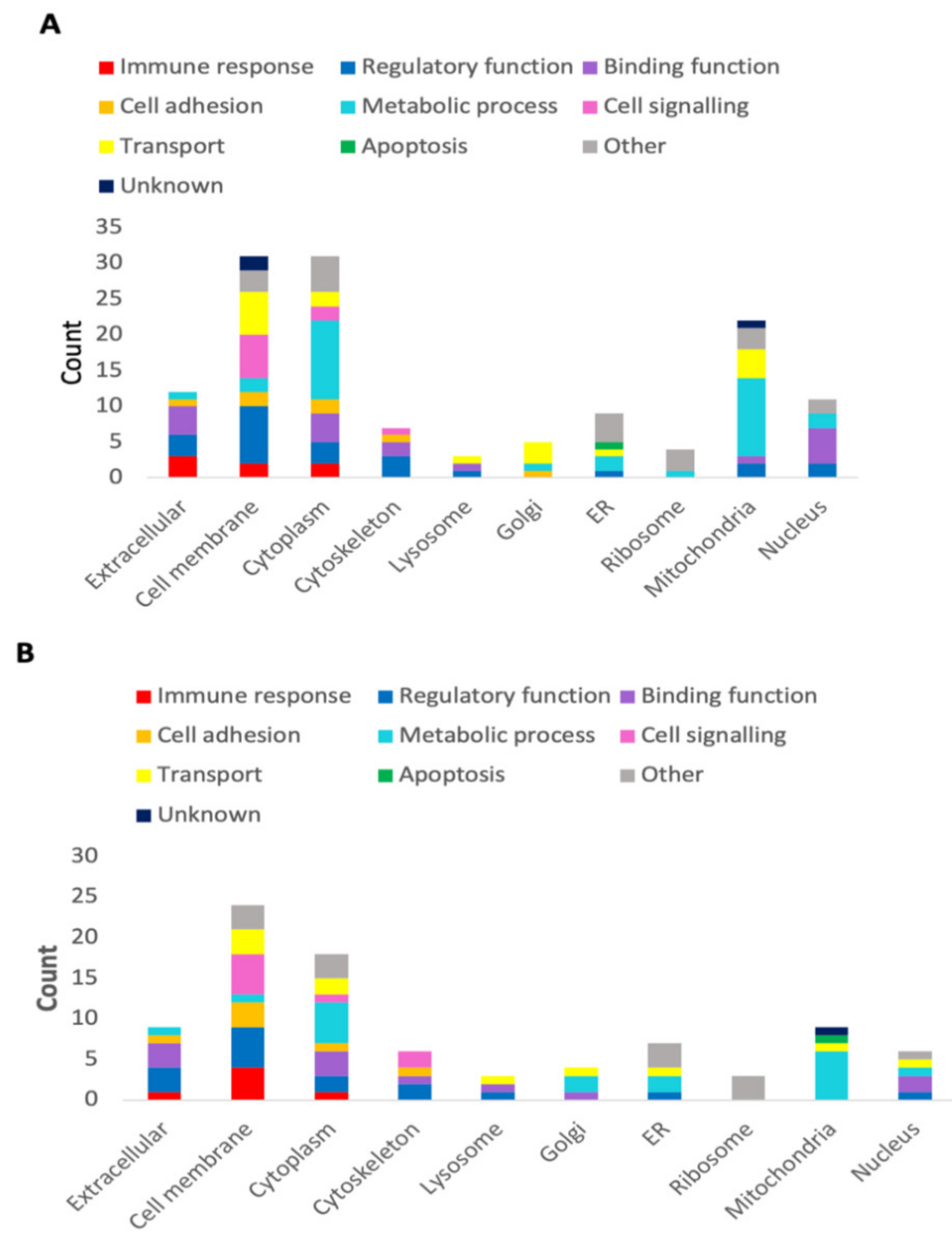
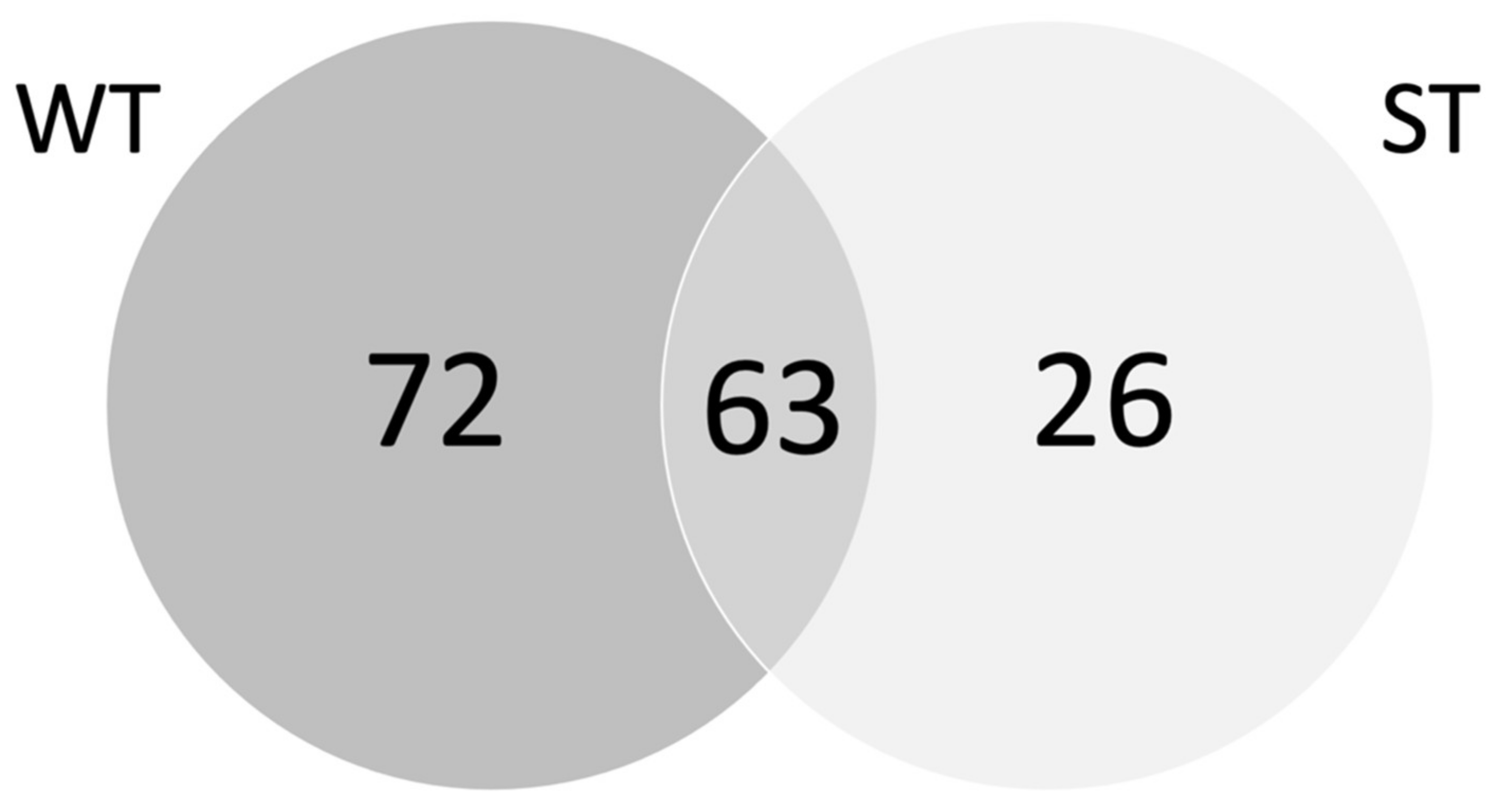
3.2. Tci-gal-1-Binding Glycoproteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roeber, F.; Jex, A.R.; Gasser, R.B. Impact of gastrointestinal parasitic nematodes of sheep, and the role of advanced molecular tools for exploring epidemiology and drug resistance—An Australian perspective. Parasit. Vectors 2013, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Charlier, J.; Thamsborg, S.M.; Bartley, D.J.; Skuce, P.J.; Kenyon, F.; Geurden, T.; Hoste, H.; Williams, A.R.; Sotiraki, S.; Hoglund, J.; et al. Mind the gaps in research on the control of gastrointestinal nematodes of farmed ruminants and pigs. Transbound. Emerg. Dis. 2018, 65 (Suppl. 1), 217–234. [Google Scholar] [CrossRef]
- Lane, J.; Jubb, T.; Shephard, R.; Webb-Ware, J.; Fordyce, G. Final Report Priority List of Endemic Diseases for the Red Meat Industries; Meat & Livestock Australia Limited: Sydney, Australia, 2015. [Google Scholar]
- Charlier, J.; Rinaldi, L.; Musella, V.; Ploeger, H.W.; Chartier, C.; Vineer, H.R.; Hinney, B.; von Samson-Himmelstjerna, G.; Bacescu, B.; Mickiewicz, M.; et al. Initial assessment of the economic burden of major parasitic helminth infections to the ruminant livestock industry in Europe. Prev. Vet. Med. 2020, 182, 105103. [Google Scholar] [CrossRef]
- Roeber, F.; Kahn, L. The specific diagnosis of gastrointestinal nematode infections in livestock: Larval culture technique, its limitations and alternative DNA-based approaches. Vet. Parasitol. 2014, 205, 619–628. [Google Scholar] [CrossRef]
- McNeilly, T.N.; Devaney, E.; Matthews, J.B. Teladorsagia circumcincta in the sheep abomasum: Defining the role of dendritic cells in T cell regulation and protective immunity. Parasite Immunol. 2009, 31, 347–356. [Google Scholar] [CrossRef]
- Sargison, N.D.; Jackson, F.; Bartley, D.J.; Moir, A.C.P. Failure of moxidectin to control benzimidazole-, levamisole- and ivermectin-resistant Teladorsagia circumcincta in a sheep flock. Vet. Rec. 2005, 156, 105–109. [Google Scholar] [CrossRef]
- Sutherland, I.A.; Brown, A.E.; Leathwick, D.M.; Bisset, S.A. Resistance to prophylactic treatment with macrocyclic lactone anthelmintics in Teladorsagia circumcincta. Vet. Parasitol. 2003, 115, 301–309. [Google Scholar] [CrossRef]
- Stear, M.J.; Doligalska, M.; Donskow-Schmelter, K. Alternatives to anthelmintics for the control of nematodes in livestock. Parasitology 2007, 134, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Pemberton, A.D.; Brown, J.K.; Craig, N.M.; Pate, J.; McLean, K.; Inglis, N.F.; Knox, D.; Knight, P.A. Changes in protein expression in the sheep abomasum following trickle infection with Teladorsagia circumcincta. Parasitology 2012, 139, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Stear, M.J.; Bishop, S.C.; Henderson, N.G.; Scott, I. A Key Mechanism of Pathogenesis in Sheep Infected with the Nematode Teladorsagia circumcincta. Anim. Health Res. Rev. 2003, 4, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, J.P.; Grainger, J.R.; Maizels, R.M. Helminth immunoregulation: The role of parasite secreted proteins in modulating host immunity. Mol. Biochem. Parasitol. 2009, 167, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hoerauf, A.; Satoguina, J.; Saeftel, M.; Specht, S. Immunomodulation by filarial nematodes. Parasite Immunol. 2005, 27, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Craig, H.; Wastling, J.M.; Knox, D.P. A preliminary proteomic survey of the in vitro excretory/secretory products of fourth-stage larval and adult Teladorsagia circumcincta. Parasitology 2006, 132, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Knox, D.P. Development of vaccines against gastrointestinal nematodes. Parasitology 2000, 120, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Finlay, C.M.; Walsh, K.P.; Mills, K.H.G. Induction of regulatory cells by helminth parasites: Exploitation for the treatment of inflammatory diseases. Immunol. Rev. 2014, 259, 206–230. [Google Scholar] [CrossRef]
- Greenhalgh, C.J.; Loukas, A.; Newton, S.E. The organization of a galectin gene from Teladorsagia circumcincta. Mol. Biochem. Parasitol. 1999, 101, 199–206. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; Toscano, M.A. Turning ‘sweet’ on immunity: Galectin–glycan interactions in immune tolerance and inflammation. Nat. Rev. Immunol. 2009, 9, 338–352. [Google Scholar] [CrossRef]
- Shi, W.; Xue, C.; Su, X.Z.; Lu, F. The roles of galectins in parasitic infections. Acta Trop. 2018, 177, 97–104. [Google Scholar] [CrossRef]
- Donskow-Lysoniewska, K.; Maruszewska-Cheruiyot, M.; Stear, M. The interaction of host and nematode galectins influences the outcome of gastrointestinal nematode infections. Parasitology 2021, 148, 648–654. [Google Scholar] [CrossRef]
- Hughes, R.C. Secretion of the galectin family of mammalian carbohydrate-binding proteins. Biochim. Biophys. Acta 1999, 1473, 172–185. [Google Scholar] [CrossRef]
- Xu, J.; Yang, F.; Yang, D.Q.; Jiang, P.; Liu, R.D.; Zhang, X.; Cui, J.; Wang, Z.Q. Molecular characterization of Trichinella spiralis galectin and its participation in larval invasion of host’s intestinal epithelial cells. Vet. Res. 2018, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popa, S.J.; Stewart, S.E.; Moreau, K. Unconventional secretion of annexins and galectins. Semin. Cell Dev. Biol. 2018, 83, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Vasta, G.R. Roles of galectins in infection. Nat. Rev. Immunol. 2009, 7, 424–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinovich, G.A.; Gruppi, A. Galectins as immunoregulators during infectious processes: From microbial invasion to the resolution of the disease. Parasite Immunol. 2005, 27, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.H.; Qiao, R.; Xin, L.; Chen, X.; Liu, C.; Zhang, X.; Shan, B.; Ghodsi, A.; Li, M. Deep learning enables de novo peptide sequencing from data-independent-acquisition mass spectrometry. Nat. Methods 2019, 16, 63–66. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Greenhalgh, C.J.; Newton, S.E. Recombinant expression of a galectin from the sheep gastrointestinal parasite Teladorsagia circumcincta: Its use in isolating galectin-glycoconjugates. Protein Express. Purif. 1999, 16, 152–159. [Google Scholar] [CrossRef]
- Arata, Y.; Hirabayashi, J.; Kasai, K. The two lectin domains of the tandem-repeat 32-kDa galectin of the nematode Caenorhabditis elegans have different binding properties. Studies with recombinant protein. J. Biochem. 1997, 121, 1002–1009. [Google Scholar] [CrossRef]
- Shakya, K.P.; Miller, J.E.; Lomax, L.G.; Burnett, D.D. Evaluation of immune response to artificial infections of Haemonchus contortus in Gulf Coast Native compared with Suffolk lambs. Vet Parasitol 2011, 181, 239–247. [Google Scholar] [CrossRef]
- Aboshady, H.M.; Stear, M.J.; Johansson, A.; Jonas, E.; Bambou, J.C. Immunoglobulins as Biomarkers for Gastrointestinal Nematodes Resistance in Small Ruminants: A systematic review. Sci. Rep. 2020, 10, 7765. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Guzman, M.A.; Cuellar-Ordaz, J.A.; Valdivia-Anda, A.G.; Buendia-Jimenez, J.A.; Alba-Hurtado, F. Correlation of parasitological and immunological parameters in sheep with high and low resistance to haemonchosis. Can. J. Anim. Sci. 2006, 86, 363–371. [Google Scholar] [CrossRef]
- Poulsen, S.S.; Kissow, H.; Hare, K.; Hartmann, B.; Thim, L. Luminal and parenteral TFF2 and TFF3 dimer and monomer in two models of experimental colitis in the rat. Regul. Pept. 2005, 126, 163–171. [Google Scholar] [CrossRef]
- Sharpe, C.; Thornton, D.J.; Grencis, R.K. A sticky end for gastrointestinal helminths; the role of the mucus barrier. Parasite Immunol. 2018, 40, 1–10. [Google Scholar] [CrossRef]
- Taupin, D.; Podolsky, D.K. Trefoil factors: Initiators of mucosal healing. Nat. Rev. Mol. Cell Biol. 2003, 4, 721–732. [Google Scholar] [CrossRef]
- Wills-Karp, M.; Rani, R.; Dienger, K.; Lewkowich, I.; Fox, J.G.; Perkins, C.; Lewis, L.; Finkelman, F.D.; Smith, D.E.; Bryce, P.J.; et al. Trefoil factor 2 rapidly induces interleukin 33 to promote type 2 immunity during allergic asthma and hookworm infection. J. Exp. Med. 2012, 209, 607–622. [Google Scholar] [CrossRef] [Green Version]
- Rowe, A.; Gondro, C.; Emery, D.; Sangster, N. Sequential microarray to identify timing of molecular responses to Haemonchus contortus infection in sheep. Vet. Parasitol. 2009, 161, 76–87. [Google Scholar] [CrossRef]
- McRae, K.M.; Stear, M.J.; Good, B.; Keane, O.M. The host immune response to gastrointestinal nematode infection in sheep. Parasite Immunol. 2015, 37, 605–613. [Google Scholar] [CrossRef]
- Stear, M.J.; Bishop, S.C.; Doligalska, M.; Duncan, J.L.; Holmes, P.H.; Irvine, J.; McCririe, L.; McKellar, Q.A.; Sinski, E.; Murray, M. Regulation of egg production, worm burden, worm length and worm fecundity by host responses in sheep infected with Ostertagia circumcincta. Parasite Immunol. 1995, 17, 643–652. [Google Scholar] [CrossRef] [PubMed]
- McRae, K.M.; Good, B.; Hanrahan, J.P.; Glynn, A.; O’Connell, M.J.; Keane, O.M. Response to Teladorsagia circumcincta infection in Scottish Blackface lambs with divergent phenotypes for nematode resistance. Vet. Parasitol. 2014, 206, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, L.; Eckersall, P.D.; Bishop, S.C.; Pettit, J.J.; Huntley, J.F.; Burchmore, R.; Stear, M.J. Genetic variation among lambs in peripheral IgE activity against the larval stages of Teladorsagia circumcincta. Parasitology 2010, 137, 1249–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negrão-Corrêa, D. Importance of immunoglobulin E (IgE) in the protective mechanism against gastrointestinal nematode infection: Looking at the intestinal mucosae. Rev. Inst. Med. Trop. São Paulo 2001, 43, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Brown, E. Integrin-associated protein (CD47) and its ligands. Trends Cell Biol. 2001, 11, 130–135. [Google Scholar] [CrossRef]
- Roebuck, K.A.; Finnegan, A. Regulation of intercellular adhesion molecule-1 (CD54) gene expression. J. Leukoc. Biol. 1999, 66, 876–888. [Google Scholar] [CrossRef] [PubMed]
- Pols, M.S.; Klumperman, J. Trafficking and function of the tetraspanin CD63. Exp. Cell Res. 2009, 315, 1584–1592. [Google Scholar] [CrossRef]
- van Putten, J.P.M.; Strijbis, K. Transmembrane mucins: Signaling receptors at the intersection of inflammation and cancer. J. Innate Immun. 2017, 9, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Hasnain, S.Z.; Evans, C.M.; Roy, M.; Gallagher, A.L.; Kindrachuk, K.N.; Barron, L.; Dickey, B.F.; Wilson, M.S.; Wynn, T.A.; Grencis, R.K.; et al. Muc5ac: A critical component mediating the rejection of enteric nematodes. J. Exp. Med. 2011, 208, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Pemberton, A.D.; Verdon, B.; Inglis, N.F.; Pearson, J.P. Sheep intelectin-2 co-purifies with the mucin Muc5ac from gastric mucus. Vet. Res. 2011, 91, 53–57. [Google Scholar] [CrossRef]
- Hoorens, P.R.; Rinaldi, M.; Li, R.W.; Goddeeris, B.; Claerebout, E.; Vercruysse, J.; Geldhof, P. Genome wide analysis of the bovine mucin genes and their gastrointestinal transcription profile. BMC Genom. 2011, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Palmai-Pallag, T.; Khodabukus, N.; Kinarsky, L.; Leir, S.H.; Sherman, S.; Hollingsworth, M.A.; Harris, A. The role of the SEA (sea urchin sperm protein, enterokinase and agrin) module in cleavage of membrane-tethered mucins. FEBS J. 2005, 272, 2901–2911. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Zach, M.; Petersson, A.C.; Svensson, F.; Johansson, D.G.; Hansson, G.C. Unfolding dynamics of the mucin SEA domain probed by force spectroscopy suggest that it acts as a cell-protective device. FEBS J. 2013, 280, 1491–1501. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.H.; Triyana, S.; Wang, R.; Das, I.; Gerloff, K.; Florin, T.H.; Sutton, P.; McGuckin, M.A. MUC1 and MUC13 differentially regulate epithelial inflammation in response to inflammatory and infectious stimuli. Mucosal Immunol. 2013, 6, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Shih, I.M. The role of CD146 (Mel-CAM) in biology and pathology. J. Pathol. 1999, 189, 4–11. [Google Scholar] [CrossRef]
- Perez-Sanchez, J.; Estensoro, I.; Redondo, M.J.; Calduch-Giner, J.A.; Kaushik, S.; Sitja-Bobadilla, A. Mucins as diagnostic and prognostic biomarkers in a fish-parasite model: Transcriptional and functional analysis. PLoS ONE 2013, 8, e65457. [Google Scholar] [CrossRef] [Green Version]
- Marcos-Lopez, M.; Calduch-Giner, J.A.; Mirimin, L.; MacCarthy, E.; Rodger, H.D.; O’Connor, I.; Sitja-Bobadilla, A.; Perez-Sanchez, J.; Piazzon, M.C. Gene expression analysis of Atlantic salmon gills reveals mucin 5 and interleukin 4/13 as key molecules during amoebic gill disease. Sci. Rep. 2018, 8, 13689. [Google Scholar] [CrossRef] [Green Version]
- Giancotti, F.G.; Ruoslahti, E. Integrin signalling. Science 1999, 285, 1028–1032. [Google Scholar] [CrossRef]
- Ginsberg, M.H.; Partridge, A.; Shattil, S.J. Integrin regulation. Curr. Opin. Cell Biol. 2005, 17, 509–516. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Gene ID | Protein Annotation | Mol. Weight (kDa) | Sequence Length | Unique Peptides | Average Abundance (Log2 LFQ) | Number of Predicted Glycosylation Sites | Signal Peptides | Predicted Cellular Location | Predicted Function | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| N-glycan | O-glycan | ||||||||||
| W5PCT5 | SLC25A31 | Solute carrier family 25 member 31 | 28.72 | 262 | 6 | 24.86 | 0 | 0 | N | Mitochondria | Membrane transport |
| W5PLZ4 | TFF2 | Trefoil factor 2 | 14.31 | 131 | 2 | 24.57 | 0 | 1 | Y | Extracellular | Regulatory function |
| W5P7F8 | CD63 | Tetraspanin | 25.83 | 236 | 4 | 24.37 | 4 | 0 | N | Cell membrane | Regulatory function |
| W5P0U4 | MUC1 | SEA domain-containing protein (Mucin 1 related) | 58.65 | 588 | 4 | 23.98 | 5 | 158 | Y | Cell membrane | Regulatory function |
| W5PBS4 | LRP1 | LDL receptor related protein 1 | 502.55 | 4526 | 14 | 23.56 | 33 | 122 | N | Cell membrane | Regulatory function |
| W5QAH5 | - | Transcription factor, GTP-binding domain | 30.79 | 277 | 1 | 23.51 | 2 | 1 | N | Nucleus | Other |
| W5QA42 | MFAP4 | Fibrinogen C-terminal domain-containing protein | 21.42 | 193 | 2 | 23.38 | 1 | 0 | Y | Extracellular | Metabolic function |
| W5PQ79 | RPS15 | Ribosomal protein S15 | 16.43 | 141 | 2 | 23.34 | 0 | 0 | N | Ribosome | Other |
| W5PZK7 | ACTA2 | Actin alpha 2, smooth muscle | 42.01 | 377 | 1 | 23.26 | 1 | 1 | N | Cytoskeleton | Regulatory function |
| W5P5W1 | ITGA3 | Integrin subunit alpha 3 | 115.18 | 1039 | 10 | 23.09 | 11 | 4 | Y | Cell membrane | Cell signalling |
| W5PEN0 | ITGA1 | Integrin subunit alpha 1 | 129.29 | 1166 | 9 | 23.09 | 19 | 15 | Y | Cell membrane | Regulatory function |
| P00922 | CA2 | Carbonic anhydrase 2 | 29.21 | 260 | 2 | 23.06 | 0 | 1 | N | Cell membrane | Other |
| W5PTZ9 | LOC114116824 | Histone H3 | 15.39 | 136 | 1 | 22.93 | 0 | 7 | N | Nucleus | Binding function |
| W5P8R7 | FCGBP | IgGFc-binding protein | 271.26 | 2544 | 17 | 22.92 | 10 | 44 | Y | Extracellular | Binding function |
| W5PNH6 | FAM234A | Family with sequence similarity 234 member A-related | 57.62 | 540 | 7 | 22.86 | 3 | 6 | N | Cell membrane | Unknown |
| W5QBH1 | CSRP1 | Cysteine and glycine rich protein 1 | 18.72 | 175 | 2 | 22.59 | 3 | 7 | N | Nucleus | Binding function |
| W5QB48 | LOC101113624 | ATP synthase, subunit F | 10.84 | 93 | 2 | 22.59 | 0 | 3 | N | Mitochondria | Metabolic function |
| W5NWN0 | GSTZ1 | Glutathione S-transferase zeta 1 | 26.7 | 240 | 4 | 22.54 | 4 | 1 | N | Mitochondria | Metabolic function |
| W5PI50 | GLRX | Glutaredoxin domain-containing protein | 11.81 | 106 | 2 | 22.53 | 1 | 0 | N | Cytoplasm | Regulatory function |
| W5NY03 | CD163 | CD163, scavenger receptor cysteine rich domain | 122.35 | 1136 | 6 | 22.45 | 7 | 13 | Y | Cell membrane | Cell signalling |
| W5Q5W2 | SCARB2 | CD36, scavenger receptor class B member 2 | 49.78 | 439 | 5 | 22.38 | 9 | 0 | N | Lysosome | Binding function |
| W5QCQ4 | DHX9 | DExH-box helicase 9 | 142.13 | 1289 | 9 | 22.36 | 7 | 29 | N | Nucleus | Regulatory function |
| W5Q436 | SLC25A5 | Mitochondrial carrier protein | 29.33 | 263 | 3 | 22.34 | 1 | 0 | N | Mitochondria | Regulatory function |
| W5PPJ0 | STT3A | STT3 oligosaccharyltransferase complex catalytic subunit A | 90.02 | 789 | 5 | 22.34 | 1 | 1 | N | ER | Metabolic function |
| W5QD93 | LOC101119050 | Dehydrogenase/reductase SDR member 4-like | 29.6 | 279 | 3 | 22.31 | 1 | 3 | N | Mitochondria | Metabolic function |
| W5NTX6 | RIPK1 | Receptor interacting serine/threonine kinase 1 | 87.21 | 770 | 1 | 22.26 | 2 | 38 | N | Cytoplasm | Regulatory function |
| W5PQK3 | PFKL | ATP-dependent 6-phosphofructokinase | 82.65 | 752 | 5 | 22.23 | 2 | 1 | N | Cytoplasm | Metabolic function |
| W5Q8K4 | SLC3A2 | Solute carrier family 3 member 2 | 63.34 | 577 | 5 | 22.22 | 3 | 7 | N | Cytoplasm | Metabolic function |
| W5QFP1 | PABPC4 | Polyadenylate-binding protein | 72.31 | 660 | 2 | 22.19 | 0 | 8 | N | Cytoplasm | Binding function |
| W5PQS4 | PROCR | MHC class 1-like antigen recognition domain | 27.09 | 241 | 3 | 22.18 | 4 | 1 | Y | Cytoskeleton | Cell signalling |
| W5QJ31 | MTHFD1 | Methylenetetrahydrofolate dehydrogenase 1 | 105.89 | 977 | 6 | 22.17 | 0 | 4 | N | Cytoplasm | Metabolic function |
| Q863C4 | ITGB6 | Integrin beta-6 | 85.75 | 787 | 3 | 22.17 | 7 | 17 | Y | Cell membrane | Cell signalling |
| W5QFZ8 | RPL13 | Ribosomal protein L13 | 23.41 | 203 | 2 | 22.14 | 1 | 4 | N | Ribosome | Other |
| W5PEE9 | LAMP1 | Lysosomal associated membrane protein 1 | 42.1 | 392 | 3 | 22.13 | 17 | 5 | N | Lysosome | Regulatory function |
| W5Q9K1 | QSOX1 | Sulfhydryl oxidase | 81.6 | 747 | 5 | 22.11 | 2 | 30 | Y | Golgi | Metabolic function |
| W5NQL2 | OLA1 | Obg-like ATPase 1 | 47.25 | 417 | 6 | 22.09 | 3 | 2 | N | Nucleus | Binding function |
| W5PTV7 | ACE | Angiotensin-converting enzyme | 138.24 | 1206 | 5 | 22.08 | 6 | 12 | N | Extracellular | Binding function |
| W5P375 | TCP1 | T-complex 1 | 60.52 | 558 | 6 | 22.07 | 0 | 3 | N | Cytoplasm | Regulatory function |
| W5PVM5 | LAMA5 | Laminin subunit alpha 5 | 375.79 | 3464 | 5 | 22.07 | 13 | 173 | N | Extracellular | Regulatory function |
| W5QC22 | CD47 | CD47 molecule | 32.16 | 291 | 3 | 22.07 | 5 | 4 | N | Extracellular | Immune response |
| W5QCP9 | COL6A3 | Collagen type VI alpha 3 chain, von Willebrand factor A domain | 340.43 | 3154 | 6 | 22 | 6 | 86 | Y | Extracellular | Binding function |
| W5PHN8 | AKR | Aldo-keto reductase | 36.67 | 323 | 2 | 21.99 | 1 | 0 | N | Cytoplasm | Metabolic function |
| W5P061 | ABCC3 | ATP binding cassette subfamily C member 3 | 169.72 | 1526 | 5 | 21.97 | 6 | 13 | N | Cell membrane | Ion transport |
| W5Q633 | BLVRA | Biliverdin reductase A | 33.61 | 296 | 4 | 21.94 | 0 | 0 | N | Cytoplasm | Metabolic function |
| W5PEH7 | SRP68 | Signal recognition particle subunit SRP68 | 70.9 | 626 | 6 | 21.9 | 0 | 19 | N | Cytoplasm | Binding function |
| W5QEY8 | GFPT1 | Glutamine--fructose-6-phosphate transaminase 1 | 67.81 | 603 | 4 | 21.86 | 1 | 1 | N | Cytoplasm | Metabolic function |
| W5Q2E5 | PTPRC | Protein tyrosine phosphatase receptor type C | 139.33 | 1235 | 4 | 21.83 | 19 | 55 | N | Cell membrane | Cell signalling |
| W5Q496 | RPL36 | 60S ribosomal protein L36 | 12.21 | 105 | 3 | 21.81 | 0 | 4 | N | Ribosome | Metabolic function |
| W5P6U5 | LMAN1 | Lectin, mannose binding 1 | 52.87 | 465 | 5 | 21.79 | 0 | 4 | N | Golgi | Protein transport |
| W5PK26 | LASP1 | LIM and SH3 protein 1 | 29.7 | 260 | 4 | 21.74 | 0 | 9 | N | Cytoplasm | Binding function |
| Accession Number | Gene ID | Protein Annotation | Mol. Weight (kDa) | Sequence Length | Unique Peptides | Average Abundance (Log2 LFQ) | Number of Predicted Glycosylation Sites | Signal Peptides | Predicted Cellular Location | Predicted Function | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| N-glycan | O-glycan | ||||||||||
| W5PLZ4 | TFF2 | Trefoil factor 2 | 14.31 | 131 | 2 | 24.21 | 0 | 1 | Y | Extracellular | Regulatory function |
| W5QAH5 | - | Transcription factor, GTP-binding domain | 30.79 | 277 | 1 | 23.93 | 2 | 1 | N | Nucleus | Other |
| W5PQ79 | RPS15 | Ribosomal protein S15 | 16.43 | 141 | 2 | 23.8 | 0 | 0 | N | Ribosome | Other |
| W5P5W1 | ITGA3 | Integrin subunit alpha 3 | 115.18 | 1039 | 10 | 23.64 | 11 | 4 | Y | Cell membrane | Cell signalling |
| W5QB48 | LOC101113624 | ATP synthase, subunit F | 10.84 | 93 | 2 | 23 | 0 | 3 | N | Mitochondria | Metabolic function |
| W5PBS4 | LRP1 | LDL receptor related protein 1 | 502.55 | 4526 | 14 | 22.89 | 33 | 122 | N | Cell membrane | Regulatory function |
| W5PEN0 | ITGA1 | Integrin subunit alpha 1 | 129.29 | 1166 | 9 | 22.86 | 19 | 15 | Y | Cell membrane | Regulatory function |
| W5P1J8 | CAO | Copper amine oxidase | 85.25 | 766 | 3 | 22.71 | 4 | 11 | Y | Cytoplasm | Metabolic function |
| Q863C4 | ITGB6 | Integrin beta-6 | 85.75 | 787 | 3 | 22.58 | 7 | 17 | Y | Cell membrane | Cell signalling |
| W5Q9K1 | QSOX1 | Sulfhydryl oxidase | 81.6 | 747 | 5 | 22.46 | 2 | 30 | Y | Golgi | Metabolic function |
| W5P5W6 | NDRG1 | N-myc downstream regulated 1 | 44.65 | 414 | 3 | 22.36 | 0 | 19 | N | Cytoplasm | Regulatory function |
| W5NVK4 | SPCS1 | Signal peptidase complex subunit 1 | 18.11 | 163 | 1 | 22.34 | 1 | 2 | N | ER | Other |
| W5NY03 | CD163 | CD163, scavenger receptor cysteine rich domain | 122.35 | 1136 | 6 | 22.3 | 7 | 13 | Y | Cell membrane | Cell signalling |
| W5P3H8 | IGF2R | Insulin like growth factor 2 receptor, mannose-6-phosphate receptor | 271.7 | 2463 | 7 | 22.28 | 16 | 13 | N | Golgi | Binding function |
| W5PVR9 | ERMP1 | Endoplasmic reticulum metallopeptidase 1 | 100.02 | 905 | 4 | 22.28 | 2 | 5 | N | ER | Metabolic function |
| W5QA42 | MFAP4 | Fibrinogen C-terminal domain-containing protein | 21.42 | 193 | 2 | 22.21 | 1 | 0 | Y | Extracellular | Metabolic function |
| W5QCH8 | NPTN | Neuroplastin | 42.43 | 376 | 6 | 22.21 | 4 | 3 | N | Cell membrane | Cell adhesion |
| W5P6A5 | SPINT2 | Serine peptidase inhibitor, Kunitz type 2 | 27.96 | 250 | 2 | 22.17 | 2 | 9 | Y | Cell membrane | Regulatory function |
| W5PEE9 | LAMP1 | Lysosomal associated membrane protein 1 | 42.1 | 392 | 3 | 22.15 | 17 | 5 | N | Lysosome | Regulatory function |
| W5QC22 | CD47 | CD47 molecule | 32.16 | 291 | 3 | 22.12 | 5 | 4 | N | Cell membrane | Cell adhesion |
| W5NTX6 | RIPK1 | Receptor interacting serine/threonine kinase 1 | 87.21 | 770 | 1 | 22.1 | 2 | 38 | N | Cytoplasm | Regulatory function |
| W5PTV7 | ACE | Angiotensin-converting enzyme | 138.24 | 1206 | 5 | 22.07 | 6 | 12 | N | Extracellular | Binding function |
| W5P1H4 | CTL4 | Choline transporter-like protein 4 | 79.6 | 711 | 3 | 21.98 | 8 | 6 | N | Cell membrane | Membrane transport |
| W5P180 | MTX1 | Metaxin | 32.19 | 285 | 4 | 21.95 | 0 | 5 | N | Mitochondria | Protein transport |
| W5Q0Z6 | MCAM | Melanoma cell adhesion molecule (MUC18-related) | 68.13 | 615 | 11 | 21.9 | 6 | 6 | N | Cell membrane | Immune response |
| W5NYJ4 | DKC1 | PUA domain-containing protein | 56.76 | 506 | 3 | 21.86 | 0 | 20 | N | Nucleus | Regulatory function |
| W5QG70 | MUC13 | SEA domain-containing protein | 53.93 | 508 | 2 | 21.79 | 9 | 62 | Y | Cell membrane | Cell signalling |
| W5NYN6 | RTN3 | Reticulon | 111.06 | 1028 | 1 | 21.78 | 1 | 151 | N | ER | Other |
| W5P0R5 | RPL21E | Ribosomal protein L21e | 18.41 | 160 | 1 | 21.71 | 0 | 0 | N | Ribosome | Other |
| W5Q8Y5 | HDLBP | High density lipoprotein binding protein/Vigilin | 141.91 | 1273 | 3 | 21.71 | 0 | 30 | N | Nucleus | Binding function |
| W5PQ27 | GATD3A | Glutamine Amidotransferase Like Class 1 Domain Containing 3A | 28.69 | 274 | 4 | 21.66 | 1 | 8 | Y | Mitochondria | Unknown |
| W5PK26 | LASP1 | LIM and SH3 protein 1 | 29.7 | 260 | 4 | 21.66 | 0 | 9 | N | Cytoplasm | Binding function |
| W5Q501 | GNAS | GNAS complex locus | 111.42 | 1037 | 3 | 21.63 | 1 | 71 | N | Cytoplasm | Cell signalling |
| W5PQS4 | PROCR | MHC class 1-like antigen recognition domain | 27.09 | 241 | 3 | 21.61 | 4 | 1 | Y | Cytoskeleton | Cell signalling |
| W5QJ31 | MTHFD1 | Methylenetetrahydrofolate dehydrogenase 1 | 105.89 | 977 | 6 | 21.61 | 0 | 4 | N | Cytoplasm | Metabolic function |
| W5PVM5 | LAMA5 | Laminin subunit alpha 5 | 375.79 | 3464 | 5 | 21.6 | 13 | 173 | N | Extracellular | Regulatory function |
| W5QFP1 | PABPC4 | Polyadenylate-binding protein | 72.31 | 660 | 2 | 21.6 | 0 | 8 | N | Cytoplasm | Binding function |
| W5PD64 | ARL8B | ADP ribosylation factor-like GTPase 8B | 20.41 | 176 | 3 | 21.57 | 2 | 0 | N | Lysosome | Binding function |
| W5PDH7 | NPC1 | Niemann-Pick C type protein | 141.53 | 1275 | 4 | 21.54 | 14 | 9 | Y | Lysosome | Lipid transport |
| W5P066 | CYB5B | Cytochrome b5 type B | 16.98 | 153 | 2 | 21.49 | 0 | 1 | N | Mitochondria | Metabolic function |
| W5P900 | RPL29 | 60S ribosomal protein L29 | 17.13 | 156 | 2 | 21.48 | 2 | 0 | N | Ribosome | Other |
| W5P246 | TM9SF2 | Nonaspanin | 76.13 | 666 | 3 | 21.46 | 0 | 2 | Y | Cell membrane | Membrane transport |
| W5P1G5 | ERGIC1 | Endoplasmic reticulum-golgi intermediate compartment 1 | 32.62 | 290 | 1 | 21.46 | 1 | 0 | N | Nucleus | ER-Golgi transport |
| W5P906 | DPP4 | Dipeptidyl peptidase 4 | 88.44 | 765 | 3 | 21.45 | 9 | 7 | Y | Cell membrane | Metabolic function |
| W5PWS0 | CD46 | Membrane cofactor protein CD46 | 39.43 | 363 | 3 | 21.35 | 3 | 10 | Y | Cell membrane | Immune response |
| W5PZG7 | PECAM1 | Platelet and endothelial cell adhesion molecule 1 (Ig-like) | 90.96 | 812 | 3 | 21.35 | 11 | 2 | N | Extracellular | Immune response |
| W5QG66 | ITGB5 | Integrin beta-5 | 88.13 | 802 | 3 | 21.31 | 5 | 25 | N | Cell membrane | Cell signalling |
| W5P8N8 | IDH3B | Isocitrate dehydrogenase [NAD] subunit | 42.5 | 385 | 4 | 21.27 | 0 | 6 | N | Mitochondria | Metabolic function |
| W5PXR3 | ENPP1 | Ectonucleotide pyrophosphatase/phosphodiesterase 1 | 94.68 | 823 | 4 | 21.27 | 6 | 8 | N | Extracellular | Regulatory function |
| W5Q3J8 | HLA-DRB3 | MHC class II antigen DRB3 | 27.99 | 244 | 2 | 21.23 | 1 | 1 | Y | Cell membrane | Immune response |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafidi, N.N.; Swan, J.; Faou, P.; Lowe, R.; Rajapaksha, H.; Cairns, C.; Stear, M.; Beddoe, T. Teladorsagia Circumcincta Galectin-Mucosal Interactome in Sheep. Vet. Sci. 2021, 8, 216. https://doi.org/10.3390/vetsci8100216
Hafidi NN, Swan J, Faou P, Lowe R, Rajapaksha H, Cairns C, Stear M, Beddoe T. Teladorsagia Circumcincta Galectin-Mucosal Interactome in Sheep. Veterinary Sciences. 2021; 8(10):216. https://doi.org/10.3390/vetsci8100216
Chicago/Turabian StyleHafidi, Nur Nasuha, Jaclyn Swan, Pierre Faou, Rohan Lowe, Harinda Rajapaksha, Callum Cairns, Michael Stear, and Travis Beddoe. 2021. "Teladorsagia Circumcincta Galectin-Mucosal Interactome in Sheep" Veterinary Sciences 8, no. 10: 216. https://doi.org/10.3390/vetsci8100216
APA StyleHafidi, N. N., Swan, J., Faou, P., Lowe, R., Rajapaksha, H., Cairns, C., Stear, M., & Beddoe, T. (2021). Teladorsagia Circumcincta Galectin-Mucosal Interactome in Sheep. Veterinary Sciences, 8(10), 216. https://doi.org/10.3390/vetsci8100216