Novel and Selective Rhipicephalus microplus Triosephosphate Isomerase Inhibitors with Acaricidal Activity

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Tick Strains
2.2. In Silico Analyses
2.3. Cloning of the RmTIM cDNA in the pET System, Growth of Cells, and Protein Expression
2.4. Purification of RmTIM
2.5. Homo sapiens TIM and Rabbit TIM
2.6. Triosephosphate Isomerase Activity Assays
2.7. Inhibition Screening and Compound Library
2.8. Ligand–Protein Molecular Docking
2.9. MTT Viability Assay
2.10. Effect of Compounds on BME26 Cell Cultures
2.11. Effect of Compounds on BME26 Morphology
2.12. Adults Immersion Test
2.13. Effect of Administration of Inhibitor 14 in Partially Engorged Female Ticks
2.14. Effects of RmTIM Inhibitor 14 on Tick Physiology
2.15. Oral Acute Toxicology in Mice
3. Results
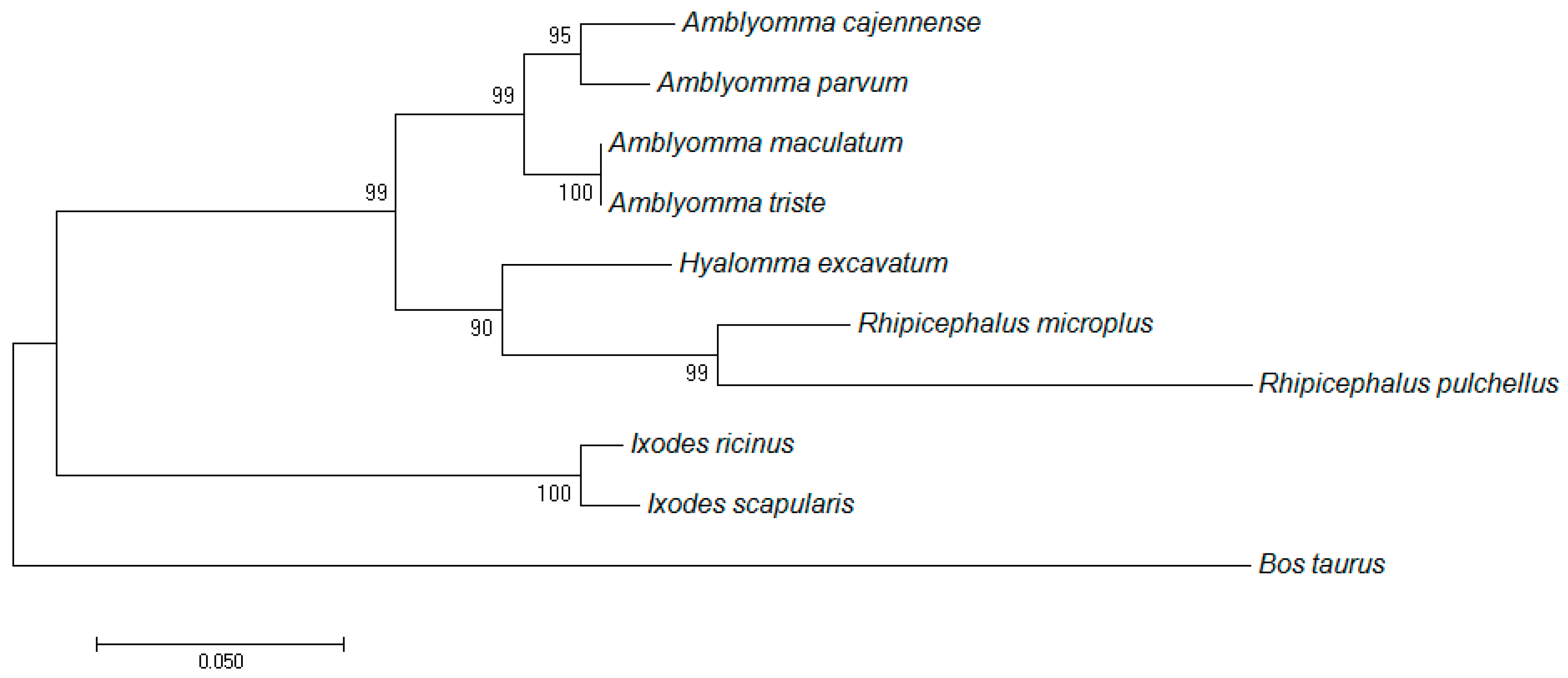
3.1. Sequence Analyses
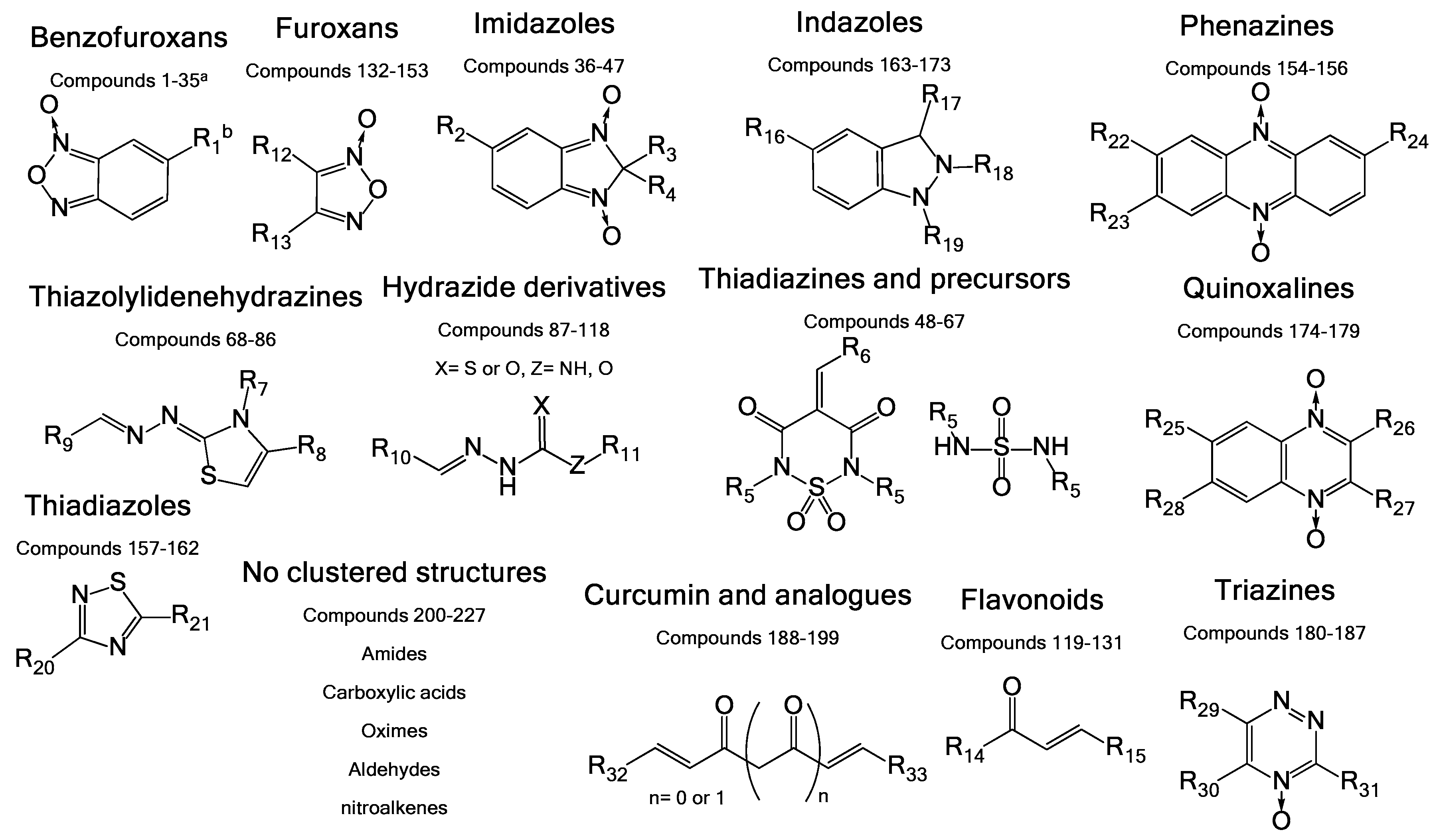
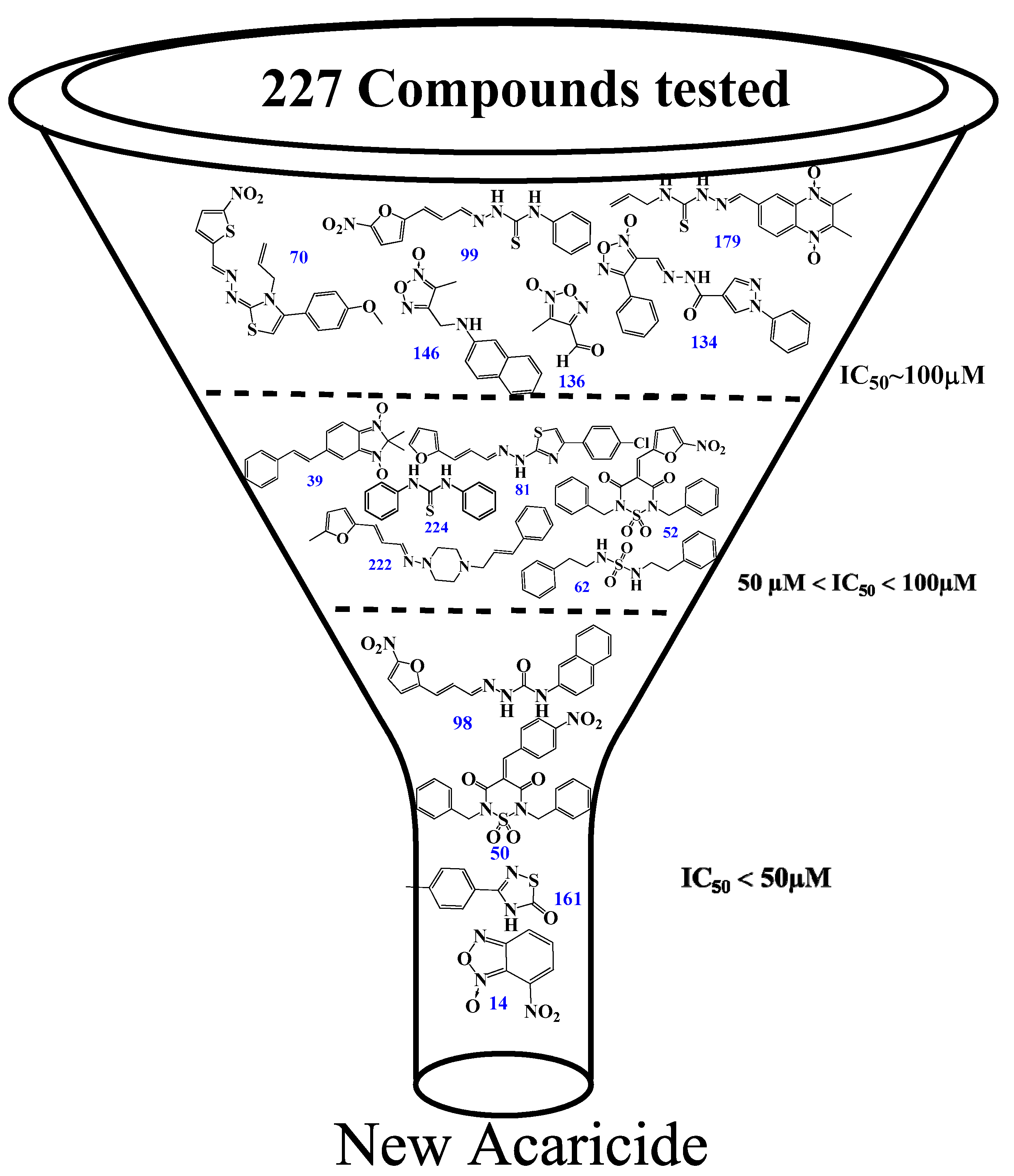
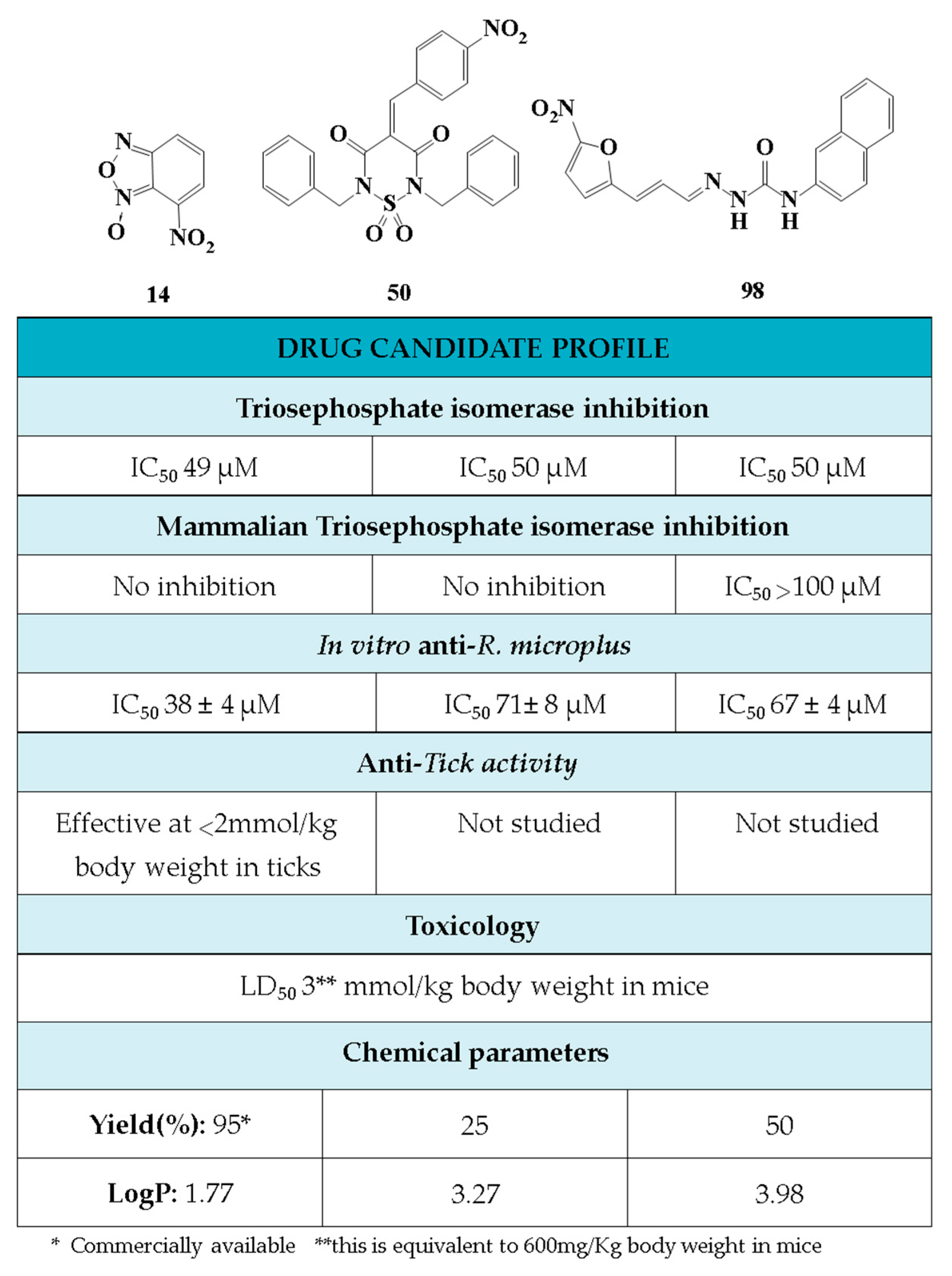
3.2. Chemical Structure Leads Used in a Primary In Vitro Screening
3.3. In Vitro Inhibition at Recombinant Enzymes
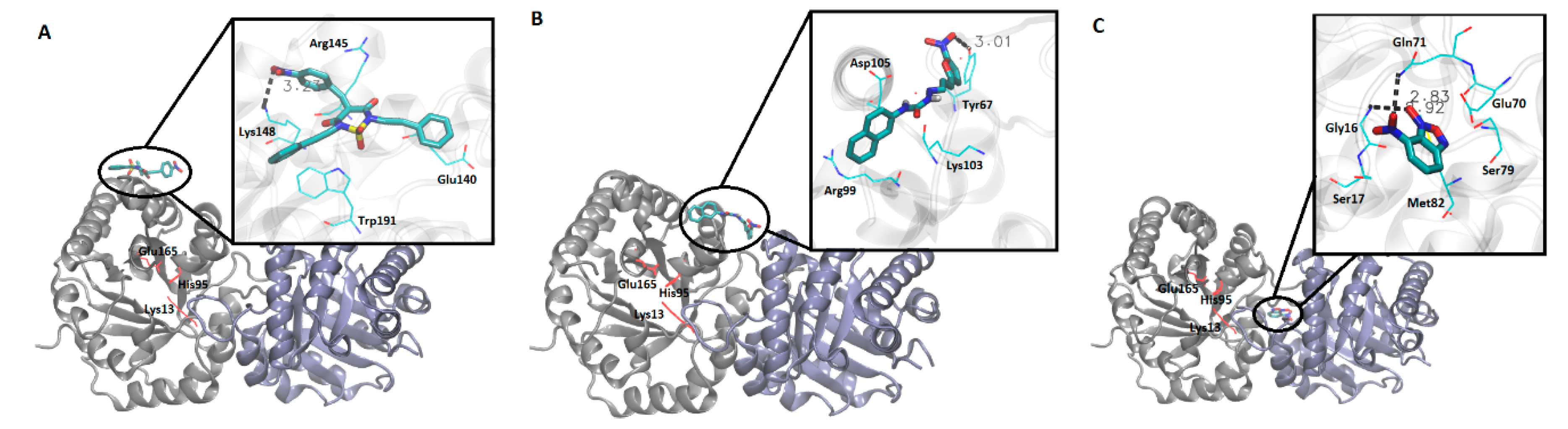
3.4. Molecular Docking Simulation Analysis
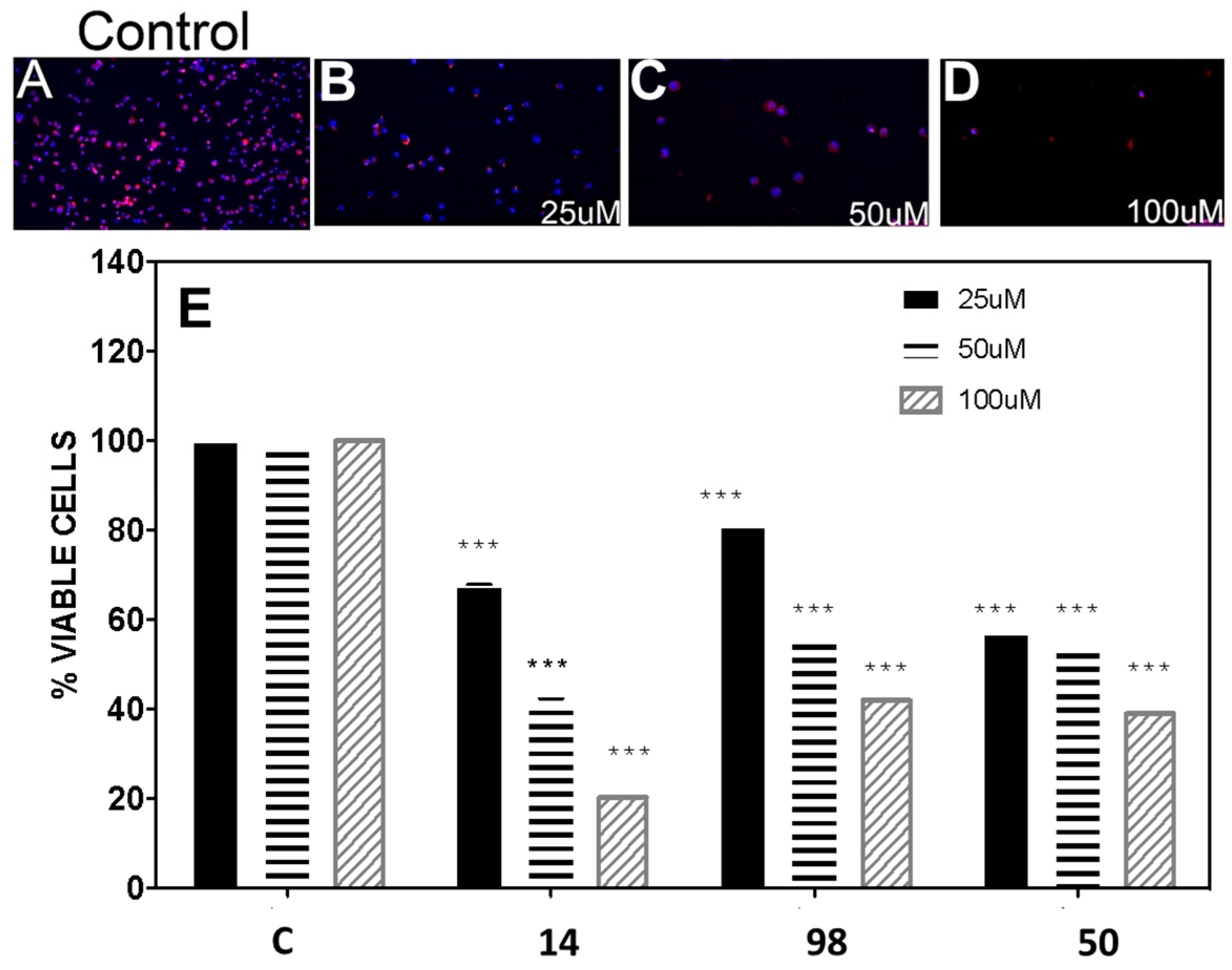
3.5. In Vitro Inhibition of the BME26 Cell Line
3.6. Effect of TIM Inhibitor 14 on the Physiology of the Tick
3.7. Effect of TIM Inhibitor in Oral Acute Toxicology in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Karbanowicz, T.P.; Nouwens, A.; Tabor, A.E.; Rodriguez-Valle, M. Comparison of protein gut samples from Rhipicephalus spp. using a crude and an innovative preparation method for proteome analysis. Vet. Sci. 2018, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, N.N. The productivity effects of cattle tick (Boophilus microplus) infestation on cattle, with particular reference to Bos indicus cattle and their crosses. Vet. Parasitol. 2006, 137, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hajdušek, O.; Síma, R.; Ayllón, N.; Jalovecká, M.; Perner, J.; de la Fuente, J.; Kopáček, P. Interaction of the tick immune system with transmitted pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Graf, J.F.; Gogolewski, R.; Leach-Bing, N.; Sabatini, G.A.; Molento, M.B.; Bordin, E.L.; Arantes, G.J. Tick control: An industry point of view. Parasitology 2004, 129 (Suppl. 1), S427–S442. [Google Scholar] [CrossRef] [PubMed]
- Klafke, G.; Webster, A.; Dall Agnol, B.; Pradel, E.; Silva, J.; de La Canal, L.H.; Becker, M.; Osório, M.F.; Mansson, M.; Barreto, R.; et al. Multiple resistance to acaricides in field populations of Rhipicephalus microplus from Rio Grande do Sul state, Southern Brazil. Ticks Tick. Borne Dis. 2017, 8, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Abbas, R.Z.; Zaman, M.A.; Colwell, D.D.; Gilleard, J.; Iqbal, Z. Acaricide resistance in cattle ticks and approaches to its management: The state of play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Li, A.Y.; Davey, R.B.; Miller, R.J.; George, J.E. Detection and Characterization of Amitraz Resistance in the Southern Cattle Tick, Boophilus microplus (Acari: Ixodidae). J. Med. Entomol. 2004, 41, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.C.; Carvalho, D.D.; Daffre, S.; Vaz, S.; Masuda, A. In vitro establishment of ivermectin-resistant Rhipicephalus microplus cell line and the contribution of ABC transporters on the resistance mechanism. Vet. Parasitol. 2014, 204, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.; Sagar, S.V.; Chigure, G.; Fular, A.; Sharma, A.K.; Nagar, G.; Kumar, S.; Saravanan, B.C.; Ghosh, S. Determination and validation of discriminating concentration of ivermectin against Rhipicephalus microplus. Vet. Parasitol. 2018, 250, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Castro-Janer, E.; Martins, J.R.; Mendes, M.C.; Namindome, A.; Klafke, G.M.; Schumaker, T.T. Diagnoses of fipronil resistance in Brazilian cattle ticks (Rhipicephalus (Boophilus) microplus) using in vitro larval bioassays. Vet. Parasitol. 2010, 173, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Parizi, L.F.; Pohl, P.C.; Masuda, A.; Junior, V.; da Silva, I. New approaches toward anti-Rhipicephalus (Boophilus) microplus tick vaccine. Rev. Bras. Parasitol. Vet. 2009, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mulenga, A.; Kim, T.K.; Ibelli, A.M.G. Deorphanization and target validation of cross-tick species conserved novel Amblyomma americanum tick saliva protein. Int. J. Parasitol. 2013, 43, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Saramago, L.; Franceschi, M.; Logullo, C.; Masuda, A.; Vaz Ida, S.; Farias, S.E.; Moraes, J. Inhibition of enzyme activity of Rhipicephalus (Boophilus) microplus triosephosphate isomerase and BME26 cell growth by monoclonal antibodies. Int. J. Mol. Sci. 2012, 13, 13118–13133. [Google Scholar] [CrossRef] [PubMed]
- Maithal, K.; Ravindra, G.; Balaram, H.; Balaram, P. Inhibition of Plasmodium falciparum triose-phosphate isomerase by chemical modification of an interface cysteine: Electrospray ionization mass spectrometric analysis of differential cysteine reactivities. J. Biol. Chem. 2002, 277, 25106–25114. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, E.; Varela, J.; Birriel, E.; Serna, E.; Torres, S.; Yaluff, G.; DeBilbao, N.V.; Aguirre-López, B.; Cabrera, N.; DíazMazariegos, S.; et al. Potent and Selective Inhibitors of Trypanosoma cruzi Triosephosphate Isomerase with Concomitant Inhibition of Cruzipain: Inhibition of Parasite Growth through Multitarget Activity. ChemMedChem 2016, 11, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Illana, V.; Rodríguez-Romero, A.; Becker, I.; Berzunza, M.; García, J.; Pérez-Montfort, R.; Cabrera, N.; López-Calahorra, F.; de Gómez-Puyou, M.T.; Gómez-Puyou, A. Perturbation of the Dimer Interface of Triosephosphate Isomerase and its Effect on Trypanosoma cruzi. PLoS Negl. Trop. Dis. 2007, 1, e1. [Google Scholar] [CrossRef] [PubMed]
- Moraes, J.; Arreola, R.; Cabrera, N.; Saramago, L.; Freitas, D.; Masuda, A.; da Silva Vaz, I., Jr.; Tuena de Gomez-Puyou, M.; Perez-Montfort, R.; Gomez-Puyou, A.; et al. Structural and biochemical characterization of a recombinant triosephosphate isomerase from Rhipicephalus (Boophilus) microplus. Insect Biochem. Mol. Biol. 2011, 41, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Téllez-Valencia, A.; Olivares-Illana, V.; Hernández-Santoyo, A.; Pérez-Montfort, R.; Costas, M.; Rodríguez-Romero, A.; López-Calahorra, F.; Tuena De Gómez-Puyou, M.; Gómez-Puyou, A. Inactivation of Triosephosphate Isomerase from Trypanosoma cruzi by an Agent that Perturbs its Dimer Interface. J. Mol. Biol. 2004, 341, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- Reck, J.; Klafke, G.M.; Webster, A.; Agnol, B.D.; Scheffer, R.; Souza, U.A.; Corassini, V.B.; Vargas, R.; Silveira, J.; Ricardo, J.; et al. First report of fluazuron resistance in Rhipicephalus microplus: A field tick population resistant to six classes of acaricides. Vet. Parasitol. 2014, 201, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Eiden, A.L.; Kaufman, P.E.; Oi, F.M.; Dark, M.J.; Bloomquist, J.R.; Miller, R.J. Determination of metabolic resistance mechanisms in pyrethroid-resistant and fipronil-tolerant brown dog ticks. Med. Vet. Entomol. 2017, 31, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Canale, A.; Mehlhorn, H.; Benelli, G. Application of ethnobotanical repellents and acaricides in prevention, control and management of livestock ticks: A review. Res. Vet. Sci. 2016, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Harrison, I.R.; Kozlik, A.; Mccarthy, J.F.; Palmer, B.H.; Wakerley, S.B.; Watkins, T.I.; Weighton, D.M. 1,3,5-Triazapenta-1,4-dienes: Chemical Aspects of a New Group of Pesticides. Pestic. Sci. 1973, 4, 901–910. [Google Scholar] [CrossRef]
- Bull, M.; Swindale, S.; Doverend, D.; Hess, E. Suppression of Boophilus microplus populations with fluazuron: An acarine growth regulator. Aust. Vet. J. 1996, 74, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Vivas, R.I.; Pérez-Cogollo, L.C.; Rosado-Aguilar, J.A.; Ojeda-Chi, M.M.; Trinidad-Martinez, I.; Miller, R.J.; Li, A.Y.; Pérez de León, A.; Guerrero, F.; Klafke, G. Rhipicephalus (Boophilus) microplus resistant to acaricides and ivermectin in cattle farms of Mexico. Rev. Bras. Parasitol. Vet. 2014, 23, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.C.; Martins, J.R.D.S.; Cesio, M.V.; Azevedo, J.L.; Heinzen, H.; de Barros, N.M. Acaricidal activity of Palicourea marcgravii, a species from the Amazon forest, on cattle tick Rhipicephalus (Boophilus) microplus. Vet. Parasitol. 2011, 179, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Tiwari, S.S.; Srivastava, S.; Sharma, A.K.; Kumar, S.; Ray, D.D.; Rawat, A.K. Acaricidal properties of Ricinus communis leaf extracts against organophosphate and pyrethroids resistant Rhipicephalus (Boophilus) microplus. Vet. Parasitol. 2013, 192, 259–267. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Souza Senra, T.; Zeringóta, V.; de Oliveira Monteiro, C.M.; Calmon, F.; Maturano, R.; Gomes, G.A.; Faza, A.; de Carvalho, M.G.; Daemon, E. Assessment of the acaricidal activity of carvacrol, (E)-cinnamaldehyde, trans-anethole, and linalool on larvae of Rhipicephalus microplus and Dermacentor nitens (Acari: Ixodidae). Parasitol. Res. 2013, 112, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, Y.; Li, Y.; Wang, Q. Design, Synthesis, Acaricidal/Insecticidal Activity, and Structure-Activity Relationship Studies of Novel Oxazolines Containing Sulfone/Sulfoxide Groups Based on the Sulfonylurea Receptor Protein-Binding Site. J. Agric. Food Chem. 2016, 64, 3034–3040. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, G.; Aguirre-López, B.; Varela, J.; Cabrera, M.; Merlino, A.; López, G.V.; Lavaggi, M.L.; Porcal, W.; Di Maio, R.; González, M.; et al. Massive screening yields novel and selective Trypanosoma cruzi triosephosphate isomerase dimer-interface-irreversible inhibitors with anti-trypanosomal activity. Eur. J. Med. Chem. 2010, 45, 5767–5772. [Google Scholar] [CrossRef] [PubMed]
- Cerecetto, H.; González, M. Anti-T. cruzi agents: Our experience in the evaluation of more than five hundred compounds. Mini Rev. Med. Chem. 2008, 8, 1355–1383. [Google Scholar] [CrossRef] [PubMed]
- Reck, J.; Berger, M.; Terra, R.M.S.; Marks, F.S.; da Silva Vaz, I.; Guimarães, J.A.; Termignoni, C. Systemic alterations of bovine hemostasis due to Rhipicephalus (Boophilus) microplus infestation. Res. Vet. Sci. 2009, 86, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nicholas, K.B.; Nicholas, H.B.J.; Deerfield, D.W.I. Analysis and Visualization of Genetic Variation. EMBNet News 1997, 4, 14. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, B.; Costas, M.; Cabrera, N.; Mendoza-Hernandez, G.; Helseth, D.L., Jr.; Fernandez, P.; de Gomez-Puyou, M.T.; Perez-Montfort, R.; Torres-Larios, A.; Gomez Puyou, A. A ribosomal misincorporation of Lys for Arg in human triosephosphate isomerase expressed in Escherichia coli gives rise to two protein populations. PLoS ONE 2011, 6, e21035. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.M.; Noce, B.D.; Waltero, C.F.; Costa, E.P.; de Abreu, L.A.; Githaka, N.W.; Moraes, J.; Gomes, H.F.; Konnai, S.; Vaz Ida, S., Jr.; et al. Non-classical gluconeogenesis-dependent glucose metabolism in Rhipicephalus microplus embryonic cell line BME26. Int. J. Mol. Sci. 2015, 16, 1821–1839. [Google Scholar] [CrossRef] [PubMed]
- Esteves, E.; Lara, F.A.; Lorenzini, D.M.; Costa, G.H.N.; Fukuzawa, A.H.; Pressinotti, L.N.; Silva, J.R.M.C.; Ferro, J.A.; Kurtti, T.J.; Munderloh, U.G.; et al. Cellular and molecular characterization of an embryonic cell line (BME26) from the tick Rhipicephalus (Boophilus) microplus. Insect Biochem. Mol. Biol. 2008, 38, 568–580. [Google Scholar] [CrossRef] [PubMed]
- Oba, M.S.P.; de Campos Pereira, M.; de Almeida, M.A.C. Ensaios in vitro pelos critérios de oba drummond de chlorpyrifos sobre linhagem supostamente resistente de Boophilus microplus. Rev. Fac. Med. Vet. Zootec. Univ. São Paulo 1976, 13, 409–420. [Google Scholar] [CrossRef]
- Fabres, A.; De Andrade, C.P.; Guizzo, M.; Sorgine, M.H.F.; Paiva-Silva, G.D.O.; Masuda, A.; Vaz Ida, S., Jr.; Logullo, C. Effect of GSK-3 activity, enzymatic inhibition and gene silencing by RNAi on tick oviposition and egg hatching. Parasitology 2010, 137, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Gonsioroski, A.V.; Bezerra, I.A.; Utiumi, K.U.; Driemeier, D.; Farias, S.E.; da Silva Vaz, I.; Masuda, A. Anti-tick monoclonal antibody applied by artificial capillary feeding in Rhipicephalus (Boophilus) microplus females. Exp. Parasitol. 2012, 130, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.C.; Klafke, G.M.; Carvalho, D.D.; Martins, J.R.; Daffre, S.; da Silva Vaz, I., Jr.; Masuda, A. ABC transporter efflux pumps: A defense mechanism against ivermectin in Rhipicephalus (Boophilus) microplus. Int. J. Parasitol. 2011, 41, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- OECD. OECD Guidelines for the Testing of Chemicals, Section 4, Test No. 425: Acute Oral Toxicity—Up-and-Down Procedure. Guidel. Test. Chem. 2001, 26. [Google Scholar] [CrossRef]
- Álvarez, G.; Varela, J.; Cruces, E.; Fernández, M.; Gabay, M.; Leal, S.M.; Escobar, P.; Sanabria, L.; Serna, E.; Torres, S.; et al. Identification of a new amide-containing thiazole as a drug candidate for treatment of chagas’ disease. Antimicrob. Agents Chemother. 2015, 59, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Zhu, N.; Mo, J. In vitro bioassay methods for laboratory screening of novel mosquito repellents. Entomol. Sci. 2014, 17, 365–370. [Google Scholar] [CrossRef]
- Wimmersberger, D.; Tritten, L.; Keiser, J. Development of an in vitro drug sensitivity assay for Trichuris muris first-stage larvae. Parasit. Vectors 2013, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Picazo, E.; Giordanetto, F. Small molecule inhibitors of ebola virus infection. Drug Discov. Today 2014, 20, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Quincoces Suarez, J.A.; Rando, D.G.; Santos, R.P.; Gonalves, C.P.; Ferreira, E.; De Carvalho, J.E.; Kohn, L.; Maria, D.A.; Faião-Flores, F.; Michalik, D.; et al. New antitumoral agents I: In vitro anticancer activity and in vivo acute toxicity of synthetic 1,5-bis(4-hydroxy-3-methoxyphenyl)-1,4-pentadien-3-one and derivatives. Bioorg. Med. Chem. 2010, 18, 6275–6281. [Google Scholar] [CrossRef] [PubMed]
- Dynamics, I.; Kurkcuoglu, Z.; Findik, D.; Akten, E.D.; Doruker, P. How an Inhibitor Bound to Subunit Interface Alters Triosephosphate. Biophys. J. 2015, 109, 1169–1178. [Google Scholar]
- Olivares-Illana, V.; Perez-Montfort, R.; Lopez-Calahorra, F.; Costas, M.; Rodriguez-Romero, A.; Tuena de Gomez-Puyou, M.; Gomez Puyou, A. Structural differences in triosephosphate isomerase from different species and discovery of a multitrypanosomatid inhibitor. Biochemistry 2006, 45, 2556–2560. [Google Scholar] [CrossRef] [PubMed]
- Fonvielle, M.; Mariano, S.; Therisod, M. New inhibitors of rabbit muscle triose-phosphate isomerase. Bioorg. Med. Chem. Lett. 2005, 15, 2906–2909. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Puyou, A.; Saavedra-Lira, E.; Becker, I.; Zubillaga, R.A.; Rojo-Dominguez, A.; Perez-Montfort, R. Using evolutionary changes to achieve species-specific inhibition of enzyme action—Studies with triosephosphate isomerase. Chem. Biol. 1995, 2, 847–855. [Google Scholar] [CrossRef]
- Alvarez, G.; Aguirre-López, B.; Cabrera, N.; Marins, E.B.; Tinoco, L.; Batthyány, C.I.; de Gómez-Puyou, M.T.; Puyou, A.G.; Pérez-Montfort, R.; Cerecetto, H.; et al. 1,2,4-thiadiazol-5(4H)-ones: A new class of selective inhibitors of Trypanosoma cruzi triosephosphate isomerase. Study of the mechanism of inhibition. J. Enzyme Inhib. Med. Chem. 2013, 28, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, G.; Martinez, J.; Aguirre-Lopez, B.; Cabrera, N.; Perez-Diaz, L.; de Gomez-Puyou, M.T.; Gomez-Puyou, A.; Perez-Montfort, R.; Garat, B.; Merlino, A.; et al. New chemotypes as Trypanosoma cruzi triosephosphate isomerase inhibitors. A deeper insight into the mechanism of inhibition. J. Enzym. Inhib Med. Chem. 2014, 29, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, G.; Martínez, J.; Varela, J.; Birriel, E.; Cruces, E.; Gabay, M.; Leal, S.M.; Escobar, P.; Aguirre-López, B.; Cabrera, N.; et al. Development of bis-thiazoles as inhibitors of triosephosphate isomerase from Trypanosoma cruzi. Identification of new non-mutagenic agents that are active in vivo. Eur. J. Med. Chem. 2015, 100, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Renslo, A.R.; McKerrow, J.H. Drug discovery and development for neglected parasitic diseases. Nat. Chem. Biol. 2006, 2, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Guo, Z.; Huang, C.; Wu, Z.; Li, Y.; Chen, X.; Fu, Y.; Ru, J.; Ali Shar, P.; Wang, Y.; et al. Large-scale Direct Targeting for Drug Repositioning and Discovery. Sci. Rep. 2015, 5, 11970. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, A.; Bolognesi, M.L. Neglected tropical diseases: Multi-target-directed ligands in the search for novel lead candidates against Trypanosoma and Leishmania. J. Med. Chem. 2009, 52, 7339–7359. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Lai, T.; Guo, T.; Chen, F.; Zhang, L.; Ding, W.; Zhang, Y. Synthesis and acaricidal activities of scopoletin phenolic ether derivatives: Qsar, molecular docking study and in silico Adme predictions. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Davey, R.B.; Pound, J.M.; Miller, J.A.; Klavons, J.A. Therapeutic and persistent efficacy of a long-acting (LA) formulation of ivermectin against Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) and sera concentration through time in treated cattle. Vet. Parasitol. 2010, 169, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Tian, Y.; Xu, M.; Wang, X.; Li, D.; Miao, F. Design, Bioactivity and structure- activity of 3-Arylpropionate Derivatives as Potential High- Efficient Acaricides against Psoroptes Cuniculi. Sci. Rep. 2018, 8, 1797. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.F.; Liu, Y.Q.; Guo, X.; Miao, X.L.; Chen, C.; Zhang, J.X.; Xu, X.S.; Yang, G.Z.; Yang, C.J.; Li, J.C.; et al. Application of Sustainable Natural Resources in Agriculture: Acaricidal and Enzyme Inhibitory Activities of Naphthoquinones and Their Analogs against Psoroptes cuniculi. Sci. Rep. 2018, 8, 1609. [Google Scholar] [CrossRef] [PubMed]
- Keenan, M.; Abbott, M.J.; Diao, H.; Wang, Z.; Best, W.M.; Keatley, S.K.; Charman, S.A.; Ryan, E.; Ioset, J. Selection and optimization of hits from a high-throughput phenotypic screen against Trypanosoma cruzi. Future Med. Chem. 2013, 5, 1733–1752. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.M.; Ejendal, K.F.K.; Avramova, L.V.; Garland-Kuntz, E.E.; Giraldo-Calderón, G.I.; Brust, T.F.; Watts, V.J.; Hill, C.A. A “genome-to-lead” approach for insecticide discovery: Pharmacological characterization and screening of Aedes aegypti D1-like dopamine receptors. PLoS Negl. Trop. Dis. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Moraes, J.; Galina, A.; Alvarenga, P.H.; Rezende, G.L.; Masuda, A.; da Silva Vaz, I., Jr.; Logullo, C. Glucose metabolism during embryogenesis of the hard tick Boophilus microplus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Rosario-Cruz, R.; Almazan, C.; Miller, R.J.; Dominguez-Garcia, D.I.; Hernandez-Ortiz, R.; de la Fuente, J. Genetic basis and impact of tick acaricide resistance. Front. Biosci. 2009, 14, 2657–2665. [Google Scholar] [CrossRef]
- Kempf, J.G.; Jung, J.Y.; Ragain, C.; Sampson, N.S.; Loria, J.P. Dynamic requirements for a functional protein hinge. J. Mol. Biol. 2007, 368, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Minini, L.; Alvarez, G.; Gonzalez, M.; Cerecetto, H.; Merlino, A. Molecular docking and molecular dynamics simulation studies of Trypanosoma cruzi triosephosphate isomerase inhibitors. Insights into the inhibition mechanism and selectivity. J. Mol. Gr. Model. 2015, 58, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Romero, A.; Hernandez-Santoyo, A.; del Pozo Yauner, L.; Kornhauser, A.; Fernandez-Velasco, D.A. Structure and inactivation of triosephosphate isomerase from Entamoeba histolytica. J. Mol. Biol. 2002, 322, 669–675. [Google Scholar] [CrossRef]
- Feyereisen, R. Evolution of insect P450. Biochem. Soc. Trans. 2006, 34, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Zhang, Y.; Chen, F.; Li, J.; Li, X.; Yue, J.; Liu, H.; Li, H.; Ran, C. Differential Analysis of the Cytochrome p450 Acaricide-Resistance Genes in Panonychus citri (Trombidiformes: Tetranychidae) Strains. Fla. Entomol. 2015, 98, 318–329. [Google Scholar] [CrossRef]
- Ramasamy, R.; Yan, S.F.; Schmidt, A.M. Methylglyoxal comes of AGE. Cell 2006, 124, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Battah, S.; Karachalias, N.; Babaei-Jadidi, R.; Horanyi, M.; Baroti, K.; Hollan, S.; Thornalley, P.J. Increased formation of methylglyoxal and protein glycation, oxidation and nitrosation in triosephosphate isomerase deficiency. Biochim. Biophys. Acta 2003, 1639, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Merlino, A.; Benitez, D.; Chavez, S.; Da Cunha, J.; Hernández, P.; Tinoco, L.W.; Campillo, N.E.; Páez, J.A.; Cerecetto, H.; González, M. Development of second generation amidinohydrazones, thio- and semicarbazones as Trypanosoma cruzi-inhibitors bearing benzofuroxan and benzimidazole 1,3-dioxide core scaffolds. Medchemcomm 2010, 1, 216–228. [Google Scholar] [CrossRef]
- Ross, F.; Hernandez, P.; Porcal, W.; Lopez, G.V.; Cerecetto, H.; Gonzalez, M.; Basika, T.; Carmona, C.; Flo, M.; Maggioli, G.; et al. Identification of thioredoxin glutathione reductase inhibitors that kill cestode and trematode parasites. PLoS ONE 2012, 7, e35033. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.; Lavaggi, M.L.; Hernandez, P.; Merlino, A.; Gerpe, A.; Porcal, W.; Boiani, M.; Ferreira, A.; Monge, A.; de Cerain, A.L.; et al. Cytotoxic, mutagenic and genotoxic effects of new anti-T. cruzi 5-phenylethenylbenzofuroxans. Contribution of phase I metabolites on the mutagenicity induction. Toxicol. Lett. 2009, 190, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Kessel, D.; Belton, J.G.; Belton, D.K.J.G. Effects of 4-Nitrobenzofurazans and Their N-Oxides on Synthesis of Protein and Nucleic Acid by Murine Leukemia Cells Effects of 4-Nitrobenzofurazans and Their N-Oxides on Synthesis of Protein and Nucleic Acid by Murine Leukemia Cells. Cancer Res. 1975, 35, 3735–3740. [Google Scholar] [PubMed]
- Thompson, S.; Kellicutt, L. Mutagenicity of anti-cancer nitrobenzofuroxans. Mutat. Res. 1977, 48, 145–153. [Google Scholar] [CrossRef]
- Deridder, B.P.; Dixon, D.P.; Beussman, D.J.; Edwards, R.; Goldsbrough, P.B. Induction of glutathione S-transferases in Arabidopsis by herbicide safeners. Plant Physiol. 2002, 130, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.-Y.; Wang, L.; Liu, F.; Cao, L.-L.; Yang, J.; Qiao, C.; Ye, Y. Benzofurazan derivatives as antifungal agents against phytopathogenic fungi. Eur. J. Med. Chem. 2014, 80, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Leyva, E.; Leyva, S.; González-Balderas, R.M.; De Loera, D.; Jiménez-Cataño, R. Fast preparation of benzofuroxans by microwave-assisted pyrolysis of o-nitrophenyl azides. Mendeleev Commun. 2013, 23, 217–218. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, A.K.; Nagar, G.; Ghosh, S. Determination and establishment of discriminating concentrations of malathion, coumaphos, fenvalerate and fipronil for monitoring acaricide resistance in ticks infesting animals. Ticks Tick. Borne Dis. 2015, 6, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Martínez, M.A.; Wu, Q.; Ares, I.; Martínez-Larrañaga, M.R.; Anadón, A.; Yuan, Z. Fipronil insecticide toxicology: Oxidative stress and metabolism. Crit. Rev. Toxicol. 2016, 46, 876–899. [Google Scholar] [CrossRef] [PubMed]
- Pardío, V.T.; Ibarra, N.D.J.; Waliszewski, K.N.; López, K.M. Effect of coumaphos on cholinesterase activity, hematology, and biochemical blood parameters of bovines in tropical regions of Mexico. J. Environ. Sci. Heal. Part B Pestic. Food Contam. Agric. Wastes 2007, 42, 359–366. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the studied compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Structure | IC50 (µM) RmTIM | % Inhibition at 100 µM HmTIM | % Inhibition at 100 µM RbTIM |
|---|---|---|---|---|
| 98 |  | 50 ± 6 | 14 | 21 |
| 50 |  | 50 ± 10 | 0 | 0 |
| 161 |  | 25 ± 3 | 28 | 0 |
| 14 |  | 49 ± 1 | 0 | 0 |
| Group | Total Oviposition | Eggs Total Weight (mg) | Larval Total Weight (mg) | Eclosion Rate |
|---|---|---|---|---|
| PBS | 14/15 | 133 ± 33 | 107 ± 47 | 79% |
| DMSO | 13/15 | 110 ± 45 | 73 ± 45 | 67% |
| C14-2000 ppm | 13/15 | 123 ± 35 | 63 ± 33 | 51% * |
| C14-500 ppm | 15/15 | 113 ± 47 | 60 ± 39 | 51% * |
| Treatment | Oviposition/Total | Total Egg Weight (mg) | Total Larval Weight (mg) | Eclosion | Eclosion Rate |
|---|---|---|---|---|---|
| G1/PBS Buffer | YES 10/13 | 30.32 ± 16.39 | 12.6 ± 9.88 | YES | 43% |
| G2/DMSO | YES 3/13 | 48.67 ± 23.54 | 23.33 ± 16.92 | YES | 47% |
| G3/C14 | NO 0/13 | - | - | NO | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saramago, L.; Gomes, H.; Aguilera, E.; Cerecetto, H.; González, M.; Cabrera, M.; Alzugaray, M.F.; Da Silva Vaz Junior, I.; Nunes da Fonseca, R.; Aguirre-López, B.; et al. Novel and Selective Rhipicephalus microplus Triosephosphate Isomerase Inhibitors with Acaricidal Activity. Vet. Sci. 2018, 5, 74. https://doi.org/10.3390/vetsci5030074
Saramago L, Gomes H, Aguilera E, Cerecetto H, González M, Cabrera M, Alzugaray MF, Da Silva Vaz Junior I, Nunes da Fonseca R, Aguirre-López B, et al. Novel and Selective Rhipicephalus microplus Triosephosphate Isomerase Inhibitors with Acaricidal Activity. Veterinary Sciences. 2018; 5(3):74. https://doi.org/10.3390/vetsci5030074
Chicago/Turabian StyleSaramago, Luiz, Helga Gomes, Elena Aguilera, Hugo Cerecetto, Mercedes González, Mauricio Cabrera, Maria Fernanda Alzugaray, Itabajara Da Silva Vaz Junior, Rodrigo Nunes da Fonseca, Beatriz Aguirre-López, and et al. 2018. "Novel and Selective Rhipicephalus microplus Triosephosphate Isomerase Inhibitors with Acaricidal Activity" Veterinary Sciences 5, no. 3: 74. https://doi.org/10.3390/vetsci5030074
APA StyleSaramago, L., Gomes, H., Aguilera, E., Cerecetto, H., González, M., Cabrera, M., Alzugaray, M. F., Da Silva Vaz Junior, I., Nunes da Fonseca, R., Aguirre-López, B., Cabrera, N., Pérez-Montfort, R., Merlino, A., Moraes, J., & Álvarez, G. (2018). Novel and Selective Rhipicephalus microplus Triosephosphate Isomerase Inhibitors with Acaricidal Activity. Veterinary Sciences, 5(3), 74. https://doi.org/10.3390/vetsci5030074