Stability of Porcine Epidemic Diarrhea Virus on Fomite Materials at Different Temperatures

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. PEDV Propagation and Quantification
2.2. Application of Virus on Fomites and Assessment of Stability
2.3. Quantitative Real-Time RT-PCR
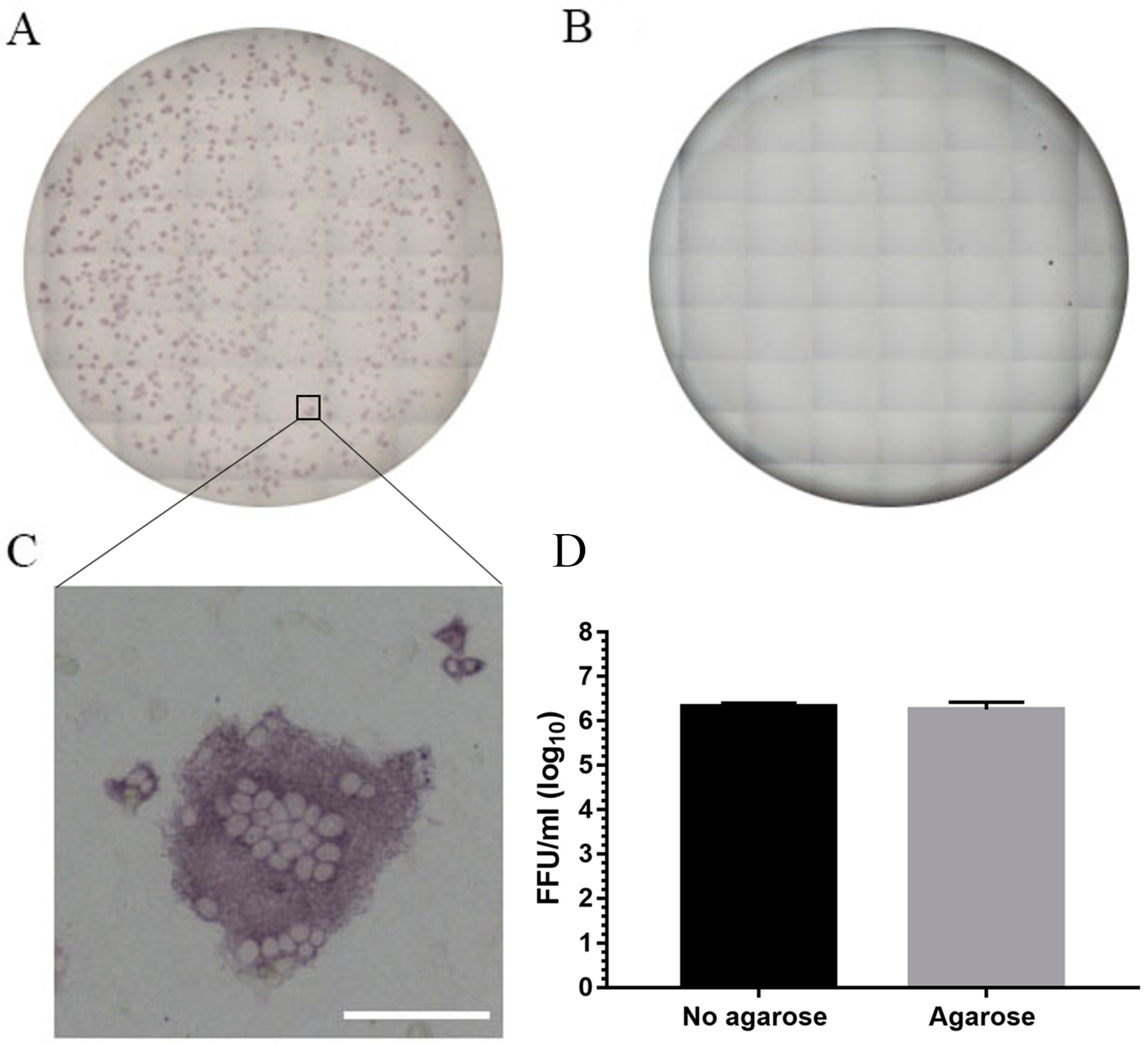
2.4. Immunoplaque Assay
2.5. Statistical Analysis
3. Results
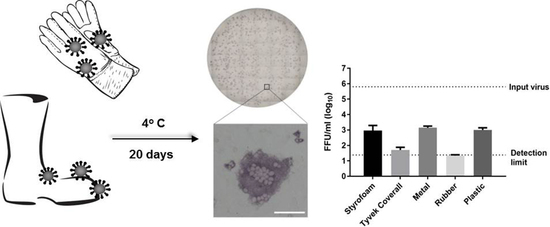
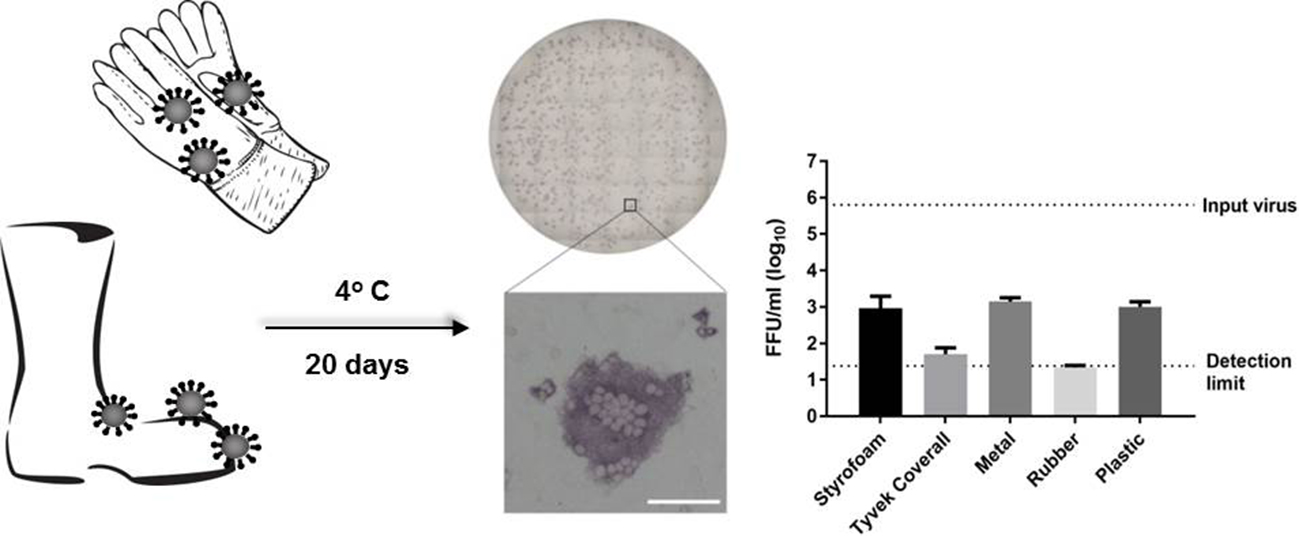
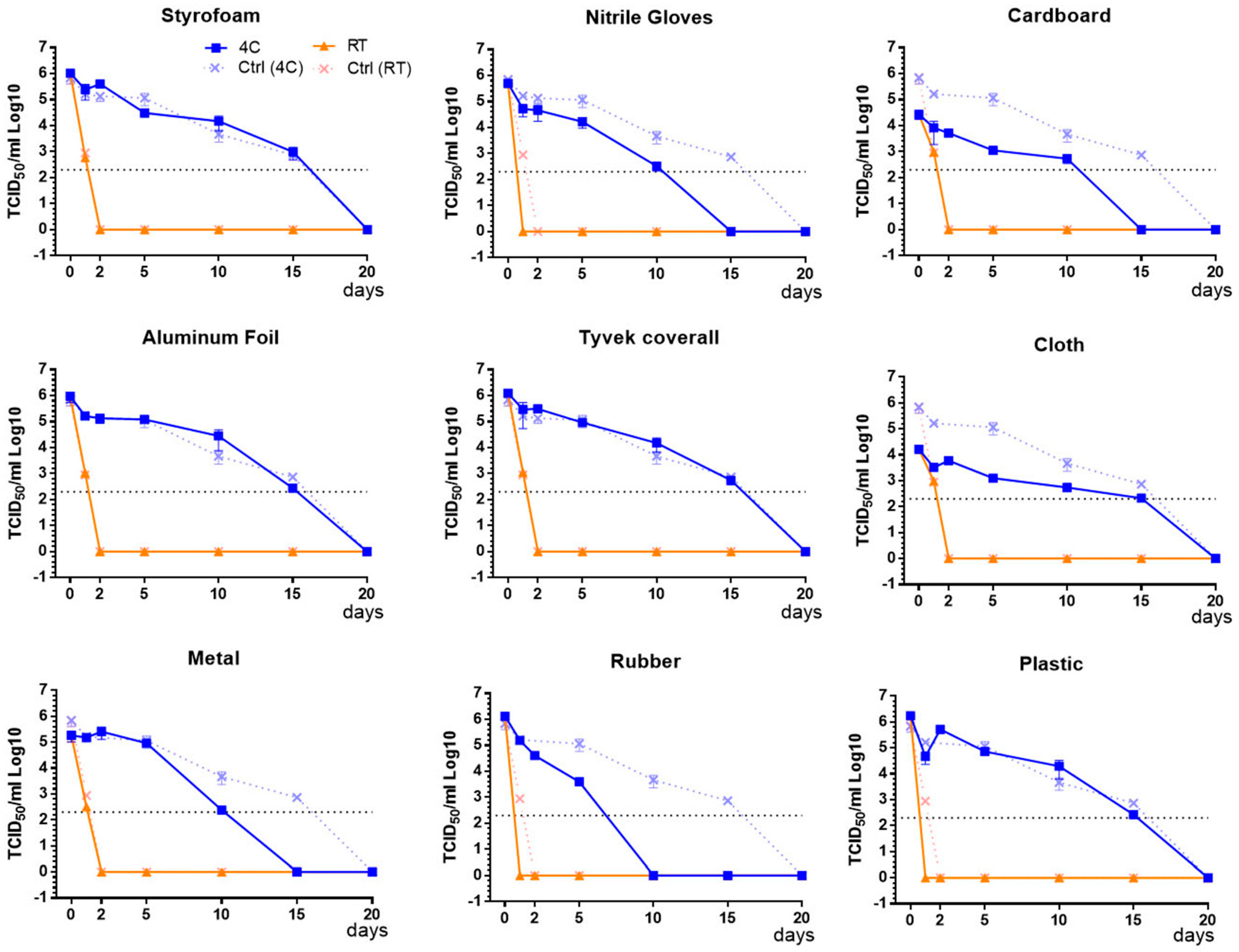
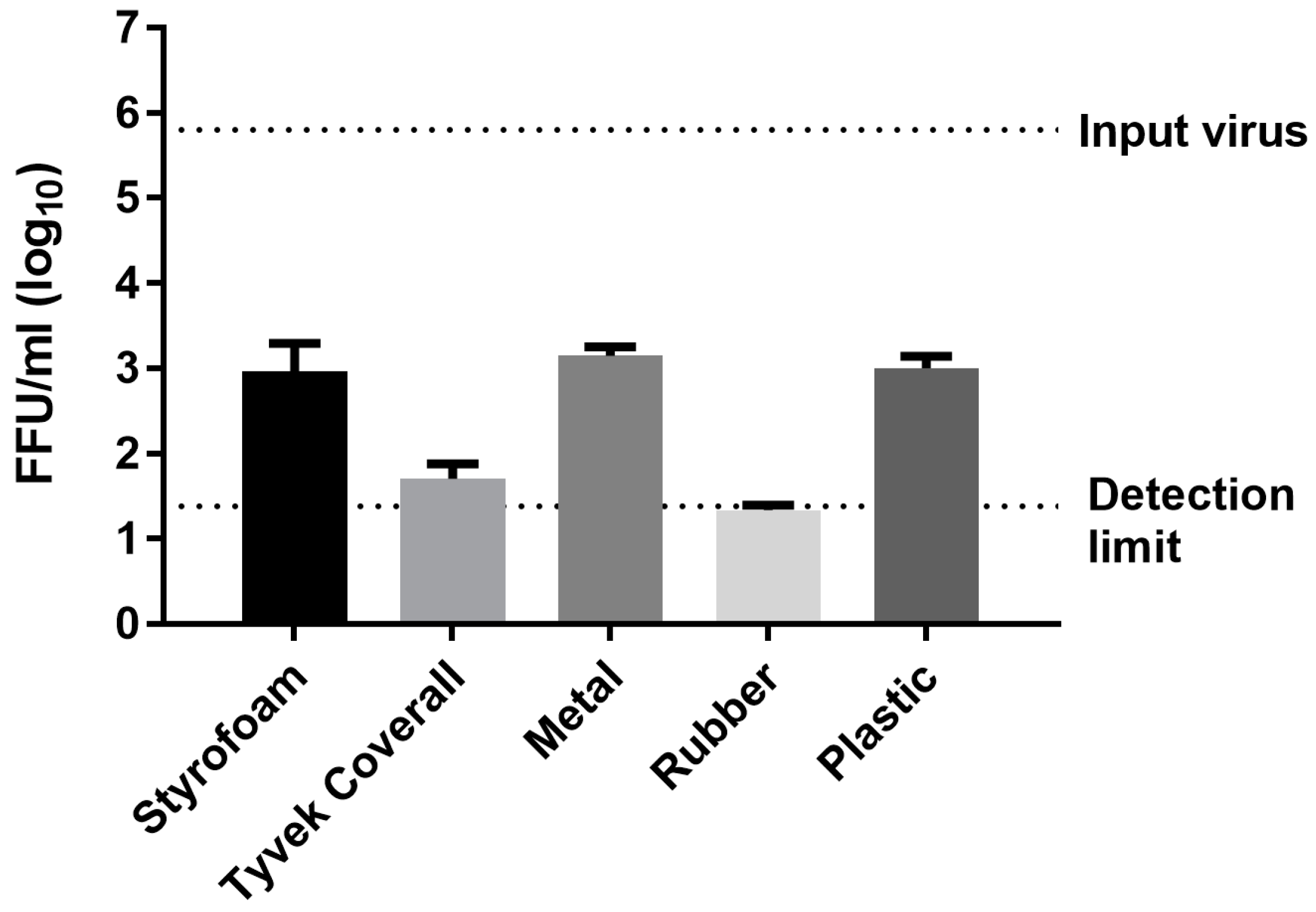
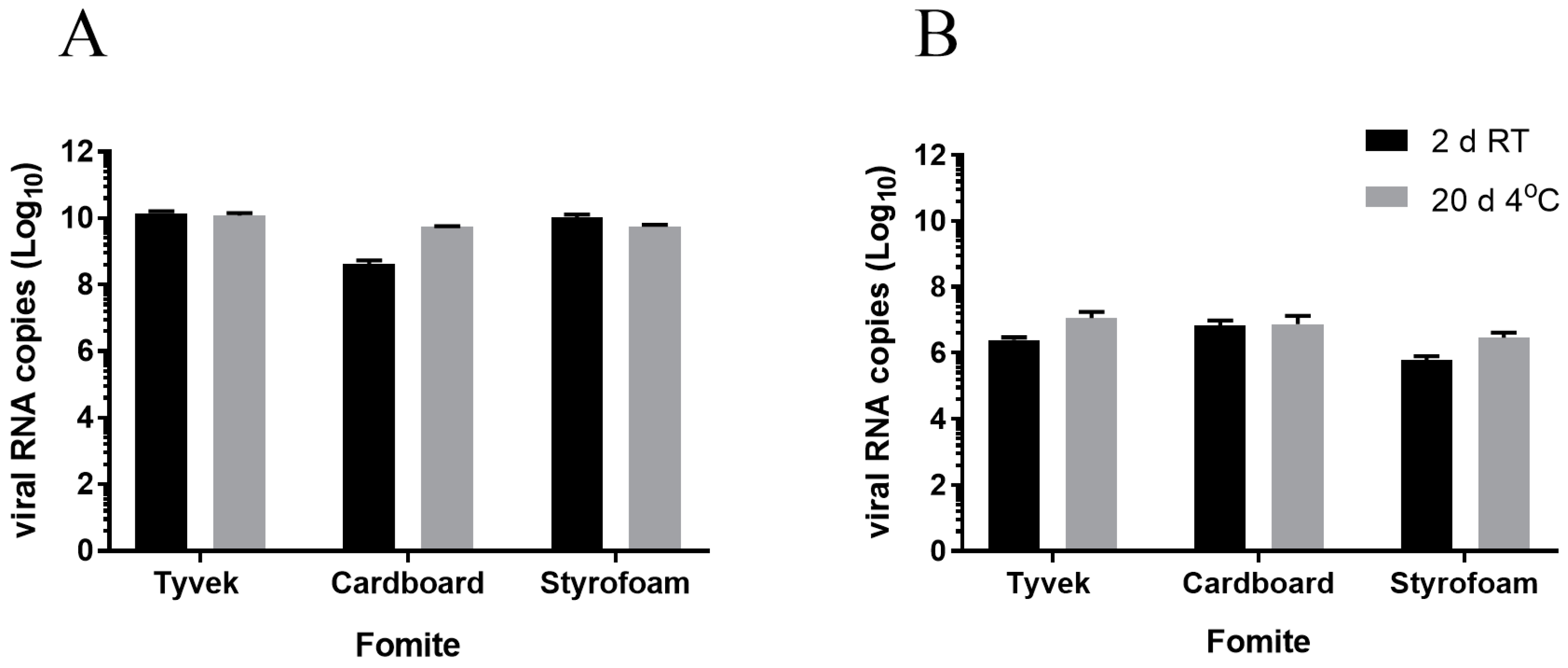
3.1. Survival of Infectious Virus on Fomite Materials
3.2. Immunoplaque Assay
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Pensaert, M.B.; Sestack, K.; Yeo, S.G.; Jung, K. Diseases of swine. In Coronaviruses; Straw, B.E., Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; Volume 48, pp. 501–524. [Google Scholar]
- Kweon, C.H.; Kwon, B.J.; Jung, T.S.; Kee, Y.J.; Hur, D.H.; Hwang, E.K.; Rhee, J.C.; An, S.H. Isolation of porcine epidemic diarrhea virus (pedv) in Korea. Korean J. Vet. Res. 1993, 33, 249–254. [Google Scholar]
- Chen, J.F.; Sun, D.B.; Wang, C.B.; Shi, H.Y.; Cui, X.C.; Liu, S.W.; Qiu, H.J.; Feng, L. Molecular characterization and phylogenetic analysis of membrane protein genes of porcine epidemic diarrhea virus isolates in China. Virus Genes 2008, 36, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Okada, K.; Ohshima, K. An outbreak of swine diarrhea of a new-type associated with coronavirus-like particles in Japan. Nihon Juigaku Zasshi 1983, 45, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Puranaveja, S.; Poolperm, P.; Lertwatcharasarakul, P.; Kesdaengsakonwut, S.; Boonsoongnern, A.; Urairong, K.; Kitikoon, P.; Choojai, P.; Kedkovid, R.; Teankum, K.; et al. Chinese-like strain of porcine epidemic diarrhea virus, Thailand. Emerg. Infect. Dis. 2009, 15, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Dee, S.; Neill, C.; Singrey, A.; Clement, T.; Cochrane, R.; Jones, C.; Patterson, G.; Spronk, G.; Christopher-Hennings, J.; Nelson, E. Modeling the transboundary risk of feed ingredients contaminated with porcine epidemic diarrhea virus. BMC Vet. Res. 2016, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; McCluskey, B.; Brown-Reid, M.; Grear, D.; Pitcher, P.; Ramos, G.; Spencer, D.; Singrey, A. Porcine epidemic diarrhea virus introduction into the united states: Root cause investigation. Prev. Vet. Med. 2016, 123, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.W.; Dickerman, A.W.; Pineyro, P.; Li, L.; Fang, L.; Kiehne, R.; Opriessnig, T.; Meng, X.J. Origin, evolution, and genotyping of emergent porcine epidemic diarrhea virus strains in the United States. MBio 2013, 4, e00737. [Google Scholar] [CrossRef] [PubMed]
- Mole, B. Deadly pig virus slips through us borders. Nature 2013, 499, 388. [Google Scholar] [CrossRef] [PubMed]
- Ojkic, D.; Hazlett, M.; Fairles, J.; Marom, A.; Slavic, D.; Maxie, G.; Alexandersen, S.; Pasick, J.; Alsop, J.; Burlatschenko, S. The first case of porcine epidemic diarrhea in Canada. Can. Vet. J. 2015, 56, 149–152. [Google Scholar] [PubMed]
- Stevenson, G.W.; Hoang, H.; Schwartz, K.J.; Burrough, E.R.; Sun, D.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; et al. Emergence of porcine epidemic diarrhea virus in the united states: Clinical signs, lesions, and viral genomic sequences. J. Vet. Diagn. Investig. 2013, 25, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Marthaler, D.; Wang, Q.; Culhane, M.R.; Rossow, K.D.; Rovira, A.; Collins, J.; Saif, L.J. Distinct characteristics and complex evolution of pedv strains, North America, May 2013–February 2014. Emerg. Infect. Dis. 2014, 20, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Park, B. Porcine epidemic diarrhoea virus: A comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes 2012, 44, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Quist-Rybachuk, G.V.; Nauwynck, H.J.; Kalmar, I.D. Sensitivity of porcine epidemic diarrhea virus (pedv) to ph and heat treatment in the presence or absence of porcine plasma. Vet. Microbiol. 2015, 181, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Dee, S.; Clement, T.; Schelkopf, A.; Nerem, J.; Knudsen, D.; Christopher-Hennings, J.; Nelson, E. An evaluation of contaminated complete feed as a vehicle for porcine epidemic diarrhea virus infection of naive pigs following consumption via natural feeding behavior: Proof of concept. BMC Vet. Res. 2014, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Pujols, J.; Segales, J. Survivability of porcine epidemic diarrhea virus (pedv) in bovine plasma submitted to spray drying processing and held at different time by temperature storage conditions. Vet. Microbiol. 2014, 174, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Pasick, J.; Berhane, Y.; Ojkic, D.; Maxie, G.; Embury-Hyatt, C.; Swekla, K.; Handel, K.; Fairles, J.; Alexandersen, S. Investigation into the role of potentially contaminated feed as a source of the first-detected outbreaks of porcine epidemic diarrhea in Canada. Transbound. Emerg. Dis. 2014, 61, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Tun, H.M.; Cai, Z.; Khafipour, E. Monitoring survivability and infectivity of porcine epidemic diarrhea virus (pedv) in the infected on-farm earthen manure storages (ems). Front. Microbiol. 2016, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Hierholzer, J.; Killington, R. Virus isolation and quantitation. In Virology Methods Manual; Mahy, B., Kangro, H., Eds.; Academic Press: London, UK, 1996; pp. 25–46. [Google Scholar]
- Aboubakr, H.A.; El-Banna, A.A.; Youssef, M.M.; Al-Sohaimy, S.A.; Goyal, S.M. Antiviral effects of lactococcus lactis on feline calicivirus, a human norovirus surrogate. Food Environ. Virol. 2014, 6, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Casanova, L.M.; Jeon, S.; Rutala, W.A.; Weber, D.J.; Sobsey, M.D. Effects of air temperature and relative humidity on coronavirus survival on surfaces. Appl. Environ. Microbiol. 2010, 76, 2712–2717. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.F.; Xiao, C.T.; Chen, Q.; Zhang, J.; Halbur, P.G.; Opriessnig, T. The spray-drying process is sufficient to inactivate infectious porcine epidemic diarrhea virus in plasma. Vet. Microbiol. 2014, 174, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Xiao, C.T.; Gerber, P.F.; Zhang, J.; Halbur, P.G. Porcine epidemic diarrhea virus rna present in commercial spray-dried porcine plasma is not infectious to naive pigs. PLoS ONE 2014, 9, e104766. [Google Scholar] [CrossRef] [PubMed]
- Pirtle, E.C.; Beran, G.W. Stability of porcine reproductive and respiratory syndrome virus in the presence of fomites commonly found on farms. J. Am. Vet. Med. Assoc. 1996, 208, 390–392. [Google Scholar] [PubMed]
- Sattar, S.A.; Lloyd-Evans, N.; Springthorpe, V.S.; Nair, R.C. Institutional outbreaks of rotavirus diarrhoea: Potential role of fomites and environmental surfaces as vehicles for virus transmission. J. Hyg. 2009, 96, 277–289. [Google Scholar] [CrossRef]
- Thomas, J.T.; Chen, Q.; Gauger, P.C.; Gimenez-Lirola, L.G.; Sinha, A.; Harmon, K.M.; Madson, D.M.; Burrough, E.R.; Magstadt, D.R.; Salzbrenner, H.M.; et al. Effect of porcine epidemic diarrhea virus infectious doses on infection outcomes in naive conventional neonatal and weaned pigs. PLoS ONE 2015, 10, e0139266. [Google Scholar] [CrossRef] [PubMed]
- Lowe, J.; Gauger, P.; Harmon, K.; Zhang, J.; Connor, J.; Yeske, P.; Loula, T.; Levis, I.; Dufresne, L.; Main, R. Role of transportation in spread of porcine epidemic diarrhea virus infection, United States. Emerg. Infect. Dis. 2014, 20, 872–874. [Google Scholar] [CrossRef] [PubMed]
- O'Dea, E.B.; Snelson, H.; Bansal, S. Using heterogeneity in the population structure of u.S. Swine farms to compare transmission models for porcine epidemic diarrhoea. Sci. Rep. 2016, 6, 22248. [Google Scholar] [CrossRef] [PubMed]
- Dee, S.; Neill, C.; Clement, T.; Singrey, A.; Christopher-Hennings, J.; Nelson, E. An evaluation of porcine epidemic diarrhea virus survival in individual feed ingredients in the presence or absence of a liquid antimicrobial. Porcine Health Manag. 2015, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bowman, A.S.; Krogwold, R.A.; Price, T.; Davis, M.; Moeller, S.J. Investigating the introduction of porcine epidemic diarrhea virus into an ohio swine operation. BMC Vet. Res. 2015, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, D.; Chattopadhyay, S.; Lyon, W.G.; Wilson, J.T. Effect of surfactants on the survival and sorption of viruses. Environ. Sci. Technol. 2002, 36, 4017–4024. [Google Scholar] [CrossRef] [PubMed]
- Vitosh-Sillman, S.; Loy, J.D.; Brodersen, B.; Kelling, C.; Eskridge, K.; Millmier Schmidt, A. Effectiveness of composting as a biosecure disposal method for porcine epidemic diarrhea virus (pedv)-infected pig carcasses. Porcine Health Manag. 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fomite Material | Immunoplaque Assay (FFU/mL) | qRT-PCR (Ct Value) † | |
|---|---|---|---|
| Cultured | Feces | ||
| Input virus | 1.0 ± 0.7 × 106 | 17.06 ± 0.77 | 29.65 ± 1.79 |
| Room Temperature/2 days * | |||
| Cultured | Feces | ||
| Styrofoam | <24 | 16.57 ± 0.34 | 30.04 ± 0.38 |
| Tyvek coverall | <24 | 16.65 ± 0.12 | 28.61 ± 0.49 |
| Cardboard | <24 | 21.50 ± 0.39 | 27.56 ± 0.26 |
| 4C/20 days * | |||
| Cultured | Feces | ||
| Styrofoam | 9.2 ± 0.6 × 102 | 17.64 ± 0.51 | 28.89 ± 0.54 |
| Tyvek coverall | 5.0 ± 2.5 × 101 | 16.93 ± 0.53 | 27.24 ± 0.75 |
| Cardboard | <24 | 17.64 ± 0.22 | 27.69 ± 0.83 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Krishna, V.D.; Torremorell, M.; Goyal, S.M.; Cheeran, M.C.-J. Stability of Porcine Epidemic Diarrhea Virus on Fomite Materials at Different Temperatures. Vet. Sci. 2018, 5, 21. https://doi.org/10.3390/vetsci5010021
Kim Y, Krishna VD, Torremorell M, Goyal SM, Cheeran MC-J. Stability of Porcine Epidemic Diarrhea Virus on Fomite Materials at Different Temperatures. Veterinary Sciences. 2018; 5(1):21. https://doi.org/10.3390/vetsci5010021
Chicago/Turabian StyleKim, Yonghyan, Venkatramana D. Krishna, Montserrat Torremorell, Sagar M. Goyal, and Maxim C.-J. Cheeran. 2018. "Stability of Porcine Epidemic Diarrhea Virus on Fomite Materials at Different Temperatures" Veterinary Sciences 5, no. 1: 21. https://doi.org/10.3390/vetsci5010021
APA StyleKim, Y., Krishna, V. D., Torremorell, M., Goyal, S. M., & Cheeran, M. C.-J. (2018). Stability of Porcine Epidemic Diarrhea Virus on Fomite Materials at Different Temperatures. Veterinary Sciences, 5(1), 21. https://doi.org/10.3390/vetsci5010021