
Parallelisms and Contrasts in the Diverse Ecologies of the Anaplasma phagocytophilum and Borrelia burgdorferi Complexes of Bacteria in the Far Western United States

Abstract
:1. Background
2. Introduction to the Anaplasma phagocytophilum and Borrelia burgdorferi sensu lato Complexes
3. Tick-Borne Sensu Stricto Strain/Genospecies Ecology
3.1. Reservoir Hosts
3.2. Clinical Disease
3.3. Vectors
4. Ecology of Other Sensu Lato Strains/Genospecies
4.1. Anaplasma Phagocytophilum Sensu Lato
4.2. Borrelia Burgdorferi Sensu Lato
5. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brown, R.N.; Lane, R.S.; Dennis, D. Geographic Distributions of Tick-Borne Diseases and Their Vectors; ASM Press: Washington, DC, USA, 2005; pp. 363–391. [Google Scholar]
- Foley, J.E.; Foley, P.; Brown, R.N.; Lane, R.S.; Dumler, J.S.; Madigan, J.E. Ecology of granulocytic ehrlichiosis and Lyme disease in the western United States. J. Vector Ecol. 2004, 29, 41–50. [Google Scholar] [PubMed]
- Munderloh, U.; Kurtti, T. Cellular and molecular interrelationships between ticks and prokaryotic tick-borne pathogens. Ann. Rev. Entomol. 1995, 40, 221–243. [Google Scholar] [CrossRef] [PubMed]
- Rollend, L.; Fish, D.; Childs, J.E. Transovarial transmission of Borrelia spirochetes by Ixodes scapularis: A summary of the literature and recent observations. Ticks Tick-Borne Dis. 2013, 4, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Rejmanek, D.; Fleer, K.; Nieto, N. Nidicolous ticks of small mammals in Anaplasma phagocytophilum-enzootic sites in northern California. Ticks Tick-Borne Dis. 2011, 2, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Tate, C.M.; Howerth, E.W.; Mead, D.G.; Dugan, V.G.; Luttrell, M.P.; Sahora, A.I.; Munderloh, U.G.; Davidson, W.R.; Yabsley, M.J. Anaplasma odocoilei sp. nov. (family Anaplasmataceae) from white-tailed deer Odocoileus virginianus. Ticks Tick-Borne Dis. 2013, 4, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Rejmanek, D.; Bradburd, G.; Foley, J.E. Molecular characterization reveals distinct genospecies of Anaplasma phagocytophilum from diverse north American hosts. J. Med. Microbiol. 2012, 61, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Massung, R.F.; Priestley, R.A.; Miller, N.J.; Mather, T.N.; Levin, M.L. Inability of a variant strain of Anaplasma phagocytophilum to infect mice. J. Infect. Dis. 2003, 188, 1757–1763. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.N.; Lane, R.S. Lyme disease in California: A novel enzootic transmission cycle of Borrelia burgdorferi. Science 1992, 256, 1439–1442. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.L.; Castro, M.B.; Kramer, V.L.; Sumner, J.W.; Childs, J.E. Dusky-footed wood rats (Neotoma fuscipes) as reservoirs of granulocytic Ehrlichiae (Rickettsiales: Ehrlichieae) in northern California. J. Clin. Microbiol. 1999, 37, 3323–3327. [Google Scholar] [PubMed]
- Foley, J.E.; Nieto, N.C.; Massung, R.; Barbet, A.; Madigan, J.; Brown, R.N. Distinct ecologically relevant strains of Anaplasma phagocytophilum. Emerg. Infect. Dis. 2009, 15, 842–843. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.J.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and “HGE agent” as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [PubMed]
- Rejmanek, D.; Freycon, P.; Bradburd, G.; Dinstell, J.; Foley, J. Unique strains of Anaplasma phagocytophilum segregate among diverse questing and non-questing Ixodes tick species in the western United States. Ticks Tick-Borne Dis. 2013, 4, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Nieto, N.C.; Madigan, J.E.; Sykes, J. Possible differential tropism in Anaplasma phagocytophilum strains in the western U.S. Ann. N. Y. Acad. Sci. 2008, 1149, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Lane, R.S.; Fedorova, N.; Koloczek, J.; Piesman, J.; Hojgaard, A.; Sing, A.; Fingerle, V. Borrelia bissettiae sp. nov. and Borrelia californiensis sp. nov. prevail in diverse enzootic transmission cycles. Int. J. Syst. Evol. Microbiol. 2016, 66, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Vollmer, S.A.; Ogden, N.H.; Fish, D. Population genetics, taxonomy, phylogeny and evolution of Borrelia burgdorferi sensu lato. Infect. Genet. Evol. 2011, 11, 1545–1563. [Google Scholar] [CrossRef] [PubMed]
- Schwan, T.G.; Schrumpf, M.E.; Karstens, R.H.; Clover, J.R.; Wong, J.; Daugherty, M.; Struthers, M.; Rosa, P.A. Distribution and molecular analysis of Lyme disease spirochetes, Borrelia burgdorferi, isolated from ticks throughout California. J. Clin. Microbiol. 1993, 31, 3096–3108. [Google Scholar] [PubMed]
- Postic, D.; Assous, M.V.; Grimont, P.A.; Baranton, G. Diversity of Borrelia burgdorferi sensu lato evidenced by restriction fragment length polymorphism of rrf (5S)-rrl (23S) intergenic spacer amplicons. Int. J. Syst. Evol. Microbiol. 1994, 44, 743–752. [Google Scholar]
- Lane, R.S.; Pascocello, J.A. Antigenic characteristics of Borrelia burgdorferi isolates from ixodid ticks in California. J. Clin. Microbiol. 1989, 27, 2344–2349. [Google Scholar] [PubMed]
- Fedorova, N.; Kleinjan, J.E.; James, D.; Hui, L.T.; Peeters, H.; Lane, R.S. Remarkable diversity of tick or mammalian-associated Borreliae in the metropolitan San Francisco Bay Area, California. Ticks Tick-Borne Dis. 2014, 5, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Donahue, J.G.; Piesman, J.; Spielman, A. Reservoir competence of white-footed mice for Lyme disease spirochetes. Am. J. Trop. Med. Hyg. 1987, 36, 92–96. [Google Scholar] [PubMed]
- Massung, R.F.; Priestley, R.A.; Levin, M.L. Transmission route efficacy and kinetics of Anaplasma phagocytophilum infection in white-footed mouse, Peromyscus leucopus. Vector-Borne Zoonotic Dis. 2004, 4, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Bockenstedt, L.K.; Wormser, G.P. Review: Unraveling Lyme disease. Arthritis Rheumatol. 2014, 66, 2313–2323. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Choi, K.S.; Garcia-Garcia, J.C.; Barat, N.S.; Scorpio, D.G.; Garyu, J.W.; Grab, D.J.; Bakken, J.S. Human granulocytic anaplasmosis and Anaplasma phagocytophilum. Emerg. Infect. Dis. 2005, 11, 1828–1834. [Google Scholar] [CrossRef] [PubMed]
- Keesing, F.; Hersh, M.H.; Tibbetts, M.; McHenry, D.J.; Duerr, S.; Brunner, J.; Killilea, M.; LoGiudice, K.; Schmidt, K.A.; Ostfeld, R.S. Reservoir competence of vertebrate hosts for Anaplasma phagocytophilum. Emerg. Infect. Dis. 2012, 18, 2013–2016. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.E.; Kramer, V.L.; Weber, D. Experimental ehrlichiosis in dusky footed woodrats (Neotoma fuscipes). J. Wildl. Dis. 2002, 38, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.; Eisen, R.J.; Mun, J.; Salkeld, D.J.; Lane, R.S. Transmission cycles of Borrelia burgdorferi and B. bissettii in relation to habitat type in northwestern California. J. Vector Ecol. 2009, 34, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Salkeld, D.J.; Leonhard, S.; Girard, Y.A.; Hahn, N.; Mun, J.; Padgett, K.A.; Lane, R.S. Identifying the reservoir hosts of the Lyme disease spirochete Borrelia burgdorferi in California: The role of the western gray squirrel (Sciurus griseus). Am. J. Trop. Med. Hyg. 2008, 79, 535–540. [Google Scholar] [PubMed]
- Lane, R.S.; Mun, J.; Eisen, R.J.; Eisen, L. Western gray squirrel (Rodentia: Sciuridae): A primary reservoir host of Borrelia burgdorferi in Californian oak woodlands? J. Med. Entomol. 2005, 42, 388–396. [Google Scholar] [CrossRef]
- Nieto, N.C.; Foley, J.E. Reservoir competence of the redwood chipmunk (Tamias ochrogenys) for Anaplasma phagocytophilum. Vector-Borne Zoonotic Dis. 2009, 9, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Swei, A.; Bowie, V.C.; Bowie, R.C. Comparative genetic diversity of Lyme disease bacteria in northern Californian ticks and their vertebrate hosts. Ticks Tick-Borne Dis. 2015, 6, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Swei, A.; Ostfeld, R.S.; Lane, R.S.; Briggs, C.J. Effects of an invasive forest pathogen on abundance of ticks and their vertebrate hosts in a California Lyme disease focus. Oecologia 2011, 166, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Nieto, N.C.; Foley, J.E. Evaluation of squirrels (Rodentia: Sciuridae) as ecologically significant hosts for Anaplasma phagocytophilum in California. J. Med. Entomol. 2008, 45, 763–769. [Google Scholar] [CrossRef]
- Foley, J.; Stephenson, N.; Qurollo, B.; Breitschwerdt, E. A putative marker for human pathogenic strains of Anaplasma phagocytophilum correlates with geography and host, but not human tropism. Ticks Tick-Borne Dis. 2016, 7, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, N.; Hodzic, E.; Mapes, S.; Rejmanek, D.; Foley, J. A real-time PCR assay for differentiating pathogenic Anaplasma phagocytophilum from an apathogenic, woodrat-adapted genospecies from North America. Ticks Tick-Borne Dis. 2015, 6, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Hasty, J.; Lane, R. Diversity of rickettsial pathogens in Columbian black-tailed deer and their associated keds (Diptera: Hippoboscidae) and ticks (Acari: Ixodidae). J. Vector Ecol. 2016, 41, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Barlough, J.; Kimsey, R.; Madigan, J.; DeRock, E.; Poland, A. Ehrlichia spp. in cervids from California. J. Wildl. Dis. 1998, 34, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Yabsley, M.J.; Davidson, W.R.; Stallknecht, D.E.; Varela, A.S.; Swift, P.K.; Devos, J.C., Jr.; Dubay, S.A. Evidence of tick-borne organisms in mule deer (Odocoileus hemionus) from the western United States. Vector-Borne Zoonotic Dis. 2005, 5, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.A.; Thompson, M.A.; Miller, M.J.; Knerl, K.M.; Elms, S.L.; Karpowicz, J.C.; Young, J.F.; Kramer, V.L. Ecology of Borrelia burgdorferi in ticks (Acari: Ixodidae), rodents, and birds in the Sierra Nevada foothills, Placer County, California. J. Med. Entomol. 2000, 37, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.; Eisen, R.J.; Lane, R.S. The roles of birds, lizards, and rodents as hosts for the western black-legged tick Ixodes pacificus. J. Vector Ecol. 2004, 29, 295–308. [Google Scholar] [PubMed]
- Newman, E.A.; Eisen, L.; Eisen, R.J.; Fedorova, N.; Hasty, J.M.; Vaughn, C.; Lane, R.S. Borrelia burgdorferi sensu lato spirochetes in wild birds in northwestern California: Associations with ecological factors, bird behavior and tick infestation. PLoS ONE 2015, 10, e0118146. [Google Scholar] [CrossRef] [PubMed]
- Dingler, R.J.; Wright, S.A.; Donahue, A.; Madeco, P.; Foley, J. Surveillance for Ixodes pacificus and the tick-borne pathogens Anaplasma phagocytophilum and Borrelia burgdorferi in birds from California’s Inner Coast Range. Ticks Tick-Borne Dis. 2014, 5, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.N.; Peot, M.A.; Lane, R.S. Sylvatic maintenance of Borrelia burgdorferi (Spirochaetales) in Northern California: Untangling the web of transmission. J. Med. Entomol. 2006, 43, 743–751. [Google Scholar] [CrossRef]
- Salkeld, D.J.; Nieto, N.C.; Carbajales-Dale, P.; Carbajales-Dale, M.; Cinkovich, S.S.; Lambin, E.F. Disease risk & landscape attributes of tick-borne Borrelia pathogens in the San Francisco Bay Area, California. PLoS ONE 2015, 10, e0134812. [Google Scholar]
- Salkeld, D.J.; Cinkovich, S.; Nieto, N.C. Tick-borne pathogens in northwestern California, USA. Emerg. Infect. Dis. 2014, 20, 493–494. [Google Scholar] [CrossRef] [PubMed]
- Crowder, C.D.; Matthews, H.E.; Schutzer, S.; Rounds, M.A.; Luft, B.J.; Nolte, O.; Campbell, S.R.; Phillipson, C.A.; Li, F.; Sampath, R. Genotypic variation and mixtures of Lyme Borrelia in Ixodes ticks from North America and Europe. PLoS ONE 2010, 5, e10650. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L.; Girard, Y.A.; Fedorova, N.; Mun, J.; Slikas, B.; Leonhard, S.; Kitron, U.; Lane, R.S. A spatially-explicit model of acarological risk of exposure to Borrelia burgdorferi-infected Ixodes pacificus nymphs in northwestern California based on woodland type, temperature, and water vapor. Ticks Tick-Borne Dis. 2010, 1, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.S.; Mun, J.; Parker, J.M.; White, M. Columbian black-tailed deer (Odocoileus hemionus columbianus) as hosts for Borrelia spp. in northern California. J. Wildl. Dis. 2005, 41, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.; Eisen, R.J.; Chang, C.C.; Mun, J.; Lane, R.S. Acarologic risk of exposure to Borrelia burgdorferi spirochaetes: Long-term evaluations in north-western California, with implications for Lyme borreliosis risk-assessment models. Med. Vet. Entomol. 2004, 18, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Eshoo, M.W.; Carolan, H.E.; Massire, C.; Chou, D.M.; Crowder, C.D.; Rounds, M.A.; Phillipson, C.A.; Schutzer, S.E.; Ecker, D.J. Survey of Ixodes pacificus ticks in California reveals a diversity of microorganisms and a novel and widespread Anaplasmataceae species. PLoS ONE 2015, 10, e0135828. [Google Scholar] [CrossRef] [PubMed]
- Vredevoe, L.K.; Stevens, J.R.; Schneider, B.S. Detection and characterization of Borrelia bissettii in rodents from the central California coast. J. Med. Entomol. 2004, 41, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Boyce, W.M.; Brown, R.N.; Zingg, B.C.; Lefebvre, R.B.; Lane, R.S. First isolation of Borrelia burgdorferi in southern California. J. Med. Entomol. 1992, 29, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Sholty, K.E. Strain Distribtution of Borrelia burgdorferi and Anaplasma phagocytophilum in Sciurids and Woodrats in Northwestern California. Master’s Thesis, Department of Wildlife Biology, Humboldt State University, Arcata, CA, USA, 2015. [Google Scholar]
- Bissett, M.L.; Hill, W. Characterization of Borrelia burgdorferi strains isolated from Ixodes pacificus ticks in California. J. Clin. Microbiol. 1987, 25, 2296–2301. [Google Scholar] [PubMed]
- Lane, R.S.; Fedorova, N.; Kleinjan, J.E.; Maxwell, M. Eco-epidemiological factors contributing to the low risk of human exposure to ixodid tick-borne borreliae in southern California, USA. Ticks Tick-Borne Dis. 2013, 4, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Ott-Conn, C.; Worth, S.J.; Poulsen, A.; Clifford, D. An Ixodes minor and Borrelia carolinensis enzootic cycle involving a critically endangered Mojave Desert rodent. Ecol. Evol. 2014, 4, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.P.; Loomis, E.C. The Ticks of California (Acari: Ixodida); University of California Press: Berkeley, CA, USA, 1984; Volume 25, p. 239. [Google Scholar]
- Bakken, J.S.; Dumler, S. Human granulocytic anaplasmosis. Inf. Dis. Clin. N. Am. 2008, 22, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Madigan, J. Equine ehrlichiosis. Vet. Clin. N. Am. Equine Pract. 1993, 9, 423–428. [Google Scholar]
- Carrade, D.D.; Foley, J.E.; Borjesson, D.L.; Sykes, J.E. Canine granulocytic anaplasmosis: A review. J. Vet. Int. Sci. 2009, 23, 1129–1141. [Google Scholar] [CrossRef] [PubMed]
- Hardalo, C.; Quagliarello, V.; Dumler, J. Human granulocytic ehrlichiosis in Connecticut: Report of a fatal case. Clin. Inf. Dis. 1995, 21, 910–914. [Google Scholar] [CrossRef]
- Walker, D.H.; Dumler, J.S. Emergence of the ehrlichioses as human health problems. Emerg. Infect. Dis. 1996, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Scorpio, D.G.; Dumler, J.S.; Barat, N.C.; Cook, J.A.; Barat, C.E.; Stillman, B.A.; DeBisceglie, K.C.; Beall, M.J.; Chandrashekar, R. Comparative strain analysis of Anaplasma phagocytophilum infection and clinical outcomes in a canine model of granulocytic anaplasmosis. Vector-Borne Zoonotic Dis. 2011, 11, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Wormser, G.P.; Dattwyler, R.J.; Shapiro, E.D.; Halperin, J.J.; Steere, A.C.; Klempner, M.S.; Krause, P.J.; Bakken, J.S.; Strle, F.; Stanek, G.; et al. The clinical assessment, treatment, and prevention of Lyme disease, human granulocytic anaplasmosis, and babesiosis: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Inf. Dis. 2006, 43, 1089–1134. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.J.; Allan, S.; Jacobson, R.H.; Lauderdale, T.L.; Chang, Y.F.; Shin, S.J.; Thomford, J.W.; Todhunter, R.J.; Summers, B.A. Experimental Lyme disease in dogs produces arthritis and persistent infection. J. Infect. Dis. 1993, 167, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Greene, C.E.; Straubinger, R.; Levy, S. Borreliosis. In Infectious Diseases of the Dog and Cat; Elsevier: Athens, Greece; Atlanta, GA, USA, 2012; pp. 447–464. [Google Scholar]
- Hahn, C.; Mayhew, I.; Mackay, R. Borreliosis. In Equine Medicine and Surgery; Colahan, P., Mayhew, I., Merritt, A., Moore, J., Eds.; Mosby: St. Louis, MI, USA, 1999; p. 895. [Google Scholar]
- Castro, M.B.; Wright, S.A. Vertebrate hosts of Ixodes pacificus (Acari: Ixodidae) in California. J. Vector Ecol. 2007, 32, 140–149. [Google Scholar] [CrossRef]
- Furman, D.; Catts, E. Manual of Medical Entomology, 4th ed.; Cambridge University Press: Cambridge, UK, 1982; p. 207. [Google Scholar]
- Lane, R.S.; Brown, R.N.; Piesman, J.; Peavey, C.A. Vector competence of Ixodes pacificus and Dermacentor occidentalis (Acari: Ixodidae) for various isolates of Lyme disease spirochetes. J. Med. Entomol. 1994, 31, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Gordus, A.G. Prevalence of Lyme Borreliosis in Deer Mice and Ticks from Northeastern California. Ph.D. Thesis, University of California, Davis, CA, USA, 1992; p. 103. [Google Scholar]
- Banerjee, S. Isolation of Borrelia burgdorferi in British Columbia. Can. Commun. Dis. Rep. 1993, 19, 204. [Google Scholar] [PubMed]
- Banerjee, S.; Banerjee, M.; Smith, J.; Fernando, K. Lyme disease in British Columbia—An update. B. C. Med. J. 1994, 36, 540–541. [Google Scholar]
- Damrow, T.; Freedman, H.; Lane, R.; Preston, K. Is Ixodes (Ixodiopsis) angustus a vector of Lyme disease in Washington State? West. J. Med. 1989, 150, 580. [Google Scholar] [PubMed]
- Peavey, C.A.; Lane, R.S.; Damrow, T. Vector competence of Ixodes angustus (Acari: Ixodidae) for Borrelia burgdorferi sensu stricto. Exp. Appl. Acarol. 2000, 24, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Nieto, N.; Foley, J.; Bettaso, J.; Lane, R. Reptile infection with Anaplasma phagocytophilum, the causative agent of granulocytic anaplasmosis. J. Parasitol. 2009, 95, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.S.; Quistad, G. Borreliacidal factor in the blood of the western fence lizard (Sceloporus occidentalis). J. Parasitol. 1998, 84, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Swei, A.; Ostfeld, R.S.; Lane, R.S.; Briggs, C.J. Impact of the experimental removal of lizards on Lyme disease risk. Proc. R. Soc. Lond. B Biol. Sci. 2011, 278, 2970–2978. [Google Scholar] [CrossRef] [PubMed]
- Majláthová, V.; Majláth, I.; Derdáková, M.; Víchová, B.; Pet’ko, B. Borrelia lusitaniae and green lizards (Lacerta viridis), Karst Region, Slovakia. Emerg. Infect. Dis. 2006, 12, 1895–1901. [Google Scholar] [CrossRef] [PubMed]
- Baldridge, G.D.; Scoles, G.A.; Burkhardt, N.Y.; Schloeder, B.; Kurtti, T.J.; Munderloh, U.G. Transovarial transmission of Francisella-like endosymbionts and Anaplasma phagocytophilum variants in Dermacentor albipictus (Acari: Ixodidae). J. Med. Entomol. 2009, 46, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Westrom, D.; Anderson, J. The distribution and seasonal abundance of deer keds (Diptera: Hippoboscidae) on Columbian black-tailed deer (Odocoileus hemionus columbianus) in northern California. Bull. Soc. Vector Ecol. 1992, 17, 57–69. [Google Scholar]
- Massung, R.F.; Mather, T.N.; Levin, M.L. Reservoir competency of goats for the Ap-variant 1 strain of Anaplasma phagocytophilum. Infect. Immun. 2006, 74, 1373–1375. [Google Scholar] [CrossRef] [PubMed]
- Massung, R.F.; Levin, M.L.; Munderloh, U.G.; Silverman, D.J.; Lynch, M.J.; Gaywee, J.K.; Kurtti, T.J. Isolation and propagation of the Ap-Variant 1 strain of Anaplasma phagocytophilum in a tick cell line. J. Clin. Microbiol. 2007, 45, 2138–2143. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H. Borrelia carolinensis sp. nov., a novel species of the Borrelia burgdorferi sensu lato complex isolated from rodents and a tick from the south-eastern USA. Int. J. Syst. Evol. Microbiol. 2011, 61, 381–383. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.L.; Oliver, J.H., Jr.; Grego, J.M.; James, A.M.; Durden, L.A.; Banks, C.W. Host associations of ticks parasitizing rodents at Borrelia burgdorferi enzootic sites in south Carolina. J. Parasitol. 2001, 87, 1379–1386. [Google Scholar] [CrossRef]
- Neumann, L. Notes sur les Ixodides. Arch. Parasitol. 1902, 6, 109–128. [Google Scholar]
- Fairchild, G. An annotated list of the bloodsucking insects, ticks and mites known from Panama. Am. J. Trop. Med. 1943, 23, 569–591. [Google Scholar]
- Poulsen, A.; Conroy, C.; Foley, P.; Ott-Conn, C.; Roy, A.; Brown, R.; Foley, J. Ectoparasites of Microtus californicus and possible emergence of an exotic Ixodes species tick in California. J. Med. Entomol. 2015, 52, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Postic, D.; Garnier, M.; Baranton, G. Multilocus sequence analysis of atypical Borrelia burgdorferi sensu lato isolates—Description of Borrelia californiensis sp. nov., and genomospecies 1 and 2. Int. J. Med. Microbiol. 2007, 297, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.M.; Foley, J. Detection of Borrelia americana in the avian coastal tick, Ixodes auritulus (Acari: Ixodidae), collected from a bird captured in Canada. Open J. Anim. Sci. 2016, 6, 207–216. [Google Scholar] [CrossRef]
- Clark, K.L.; Leydet, B.; Hartman, S. Lyme borreliosis in human patients in Florida and Georgia, USA. Int. J. Med. Sci. 2013, 10, 915–931. [Google Scholar] [CrossRef] [PubMed]
- Strle, F.; Picken, R.N.; Cheng, Y.; Cimperman, J.; Maraspin, V.; Lotric-Furlan, S.; Ruzic-Sabljic, E.; Picken, M.M. Clinical findings for patients with Lyme borreliosis caused by Borrelia burgdorferi sensu lato with genotypic and phenotypic similarities to strain 25015. Clin. Inf. Dis. 1997, 25, 273–280. [Google Scholar]
- Rudenko, N.; Golovchenko, M.; Ruzek, D.; Piskunova, N.; Mallatova, N.; Grubhoffer, L. Molecular detection of Borrelia bissettii DNA in serum samples from patients in the Czech Republic with suspected borreliosis. FEMS Microbiol. Lett. 2009, 292, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Mokráček, A.; Piskunová, N.; Růžek, D.; Mallatová, N.; Grubhoffer, L. Detection of Borrelia bissettii in cardiac valve tissue of a patient with endocarditis and aortic valve stenosis in the Czech Republic. J. Clin. Microbiol. 2008, 46, 3540–3543. [Google Scholar] [CrossRef] [PubMed]
- Fingerle, V.; Schulte-Spechtel, U.C.; Ruzic-Sabljic, E.; Leonhard, S.; Hofmann, H.; Weber, K.; Pfister, K.; Strle, F.; Wilske, B. Epidemiological aspects and molecular characterization of Borrelia burgdorferi sl from southern Germany with special respect to the new species Borrelia spielmanii sp. nov. Int. J. Med. Microbiol. 2008, 298, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Fedorova, N.; Lane, R.S. Genetic diversity of Borrelia burgdorferi and detection of B. bissettii-like DNA in serum of north-coastal California residents. J. Clin. Microbiol. 2011, 49, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Zeidner, N.S.; Burkot, T.R.; Massung, R.; Nicholson, W.L.; Dolan, M.C.; Rutherford, J.S.; Biggerstaff, B.J.; Maupin, G.O. Transmission of the agent of human granulocytic ehrlichiosis by Ixodes spinipalpis ticks: Evidence of an enzootic cycle of dual infection with Borrelia burgdorferi in northern Colorado. J. Infect. Dis. 2000, 182, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Linsdale, J.M.; Tevis, L.P. The Dusky-Footed Wood Rat; A Record of Observations Made on the Hastings Natural History Reservation; University of California Press: Berkeley, CA, USA, 1951. [Google Scholar]
- Durand, J.; Jacquet, M.; Paillard, L.; Rais, O.; Gern, L.; Voordouw, M.J. Cross-immunity and community structure of a multiple-strain pathogen in the tick vector. Appl. Environ. Micrbiol. 2015, 81, 7740–7752. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Rejmanek, D.; Foley, C.W.; Matocq, M. Fine-scale genetic structure of woodrat populations (Genus: Neotoma) and the spatial distribution of their tick-borne pathogens. Ticks Tick-Borne Dis. 2016, 7, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Fleer, K.A.; Foley, P.; Calder, L.; Foley, J.E. Arthropod vectors and vector-borne bacterial pathogens in Yosemite National Park. J. Med. Entomol. 2011, 48, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.S.; Mun, J.; Stubbs, H. Horizontal and vertical movements of host-seeking Ixodes pacificus (Acari: Ixodidae) nymphs in a hardwood forest. J. Vector Ecol. 2009, 34, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Piovia-Scott, J. Vector biodiversity did not associate with tick-borne pathogen prevalence in small mammal communities in northern and central California. Ticks Tick-Borne Dis. 2014, 5, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum—A widespread multi-host pathogen with highly adaptive strategies. Front. Cell. Inf. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Nieto, N.C.; Foley, J.E. Meta-Analysis of coinfection and coexposure with Borrelia burgdorferi and Anaplasma phagocytophilum in humans, domestic animals, wildlife, and Ixodes ricinus-complex ticks. Vector-Borne Zoonotic Dis. 2009, 9, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; Van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; Van Maanen, C.; Butler, C.M.; Földvári, G. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasites Vectors 2014, 7, 365. [Google Scholar] [CrossRef] [PubMed]
- Bown, K.; Begon, M.; Bennett, M.; Woldehiwet, Z.; Ogden, N. Seasonal dynamics of Anaplasma phagocytophila in a rodent-tick (Ixodes trianguliceps) system, United Kingdom. Emerg. Infect. Dis. 2003, 9, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Baráková, I.; Derdáková, M.; Carpi, G.; Rosso, F.; Collini, M.; Tagliapietra, V.; Ramponi, C.; Hauffe, H.C.; Rizzoli, A. Genetic and ecologic variability among Anaplasma phagocytophilum strains, northern Italy. Emerg. Infect. Dis. 2014, 20, 1082. [Google Scholar] [CrossRef] [PubMed]
- Blaňarová, L.; Stanko, M.; Carpi, G.; Miklisová, D.; Víchová, B.; Mošanský, L.; Bona, M.; Derdáková, M. Distinct Anaplasma phagocytophilum genotypes associated with Ixodes trianguliceps ticks and rodents in central Europe. Ticks Tick-Borne Dis. 2014, 5, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.A.; Epikhina, T.I.; Yakimenko, V.V.; Malkova, M.G.; Tancev, A.K.; Bondarenko, E.I.; Ivanov, M.K.; Tikunova, N.V. Genetic variability of Anaplasma phagocytophilum in ticks and voles from Ixodes persulcatus/Ixodes trianguliceps sympatric areas from western Siberia, Russia. Ticks Tick-Borne Dis. 2014, 5, 854–863. [Google Scholar] [CrossRef] [PubMed]
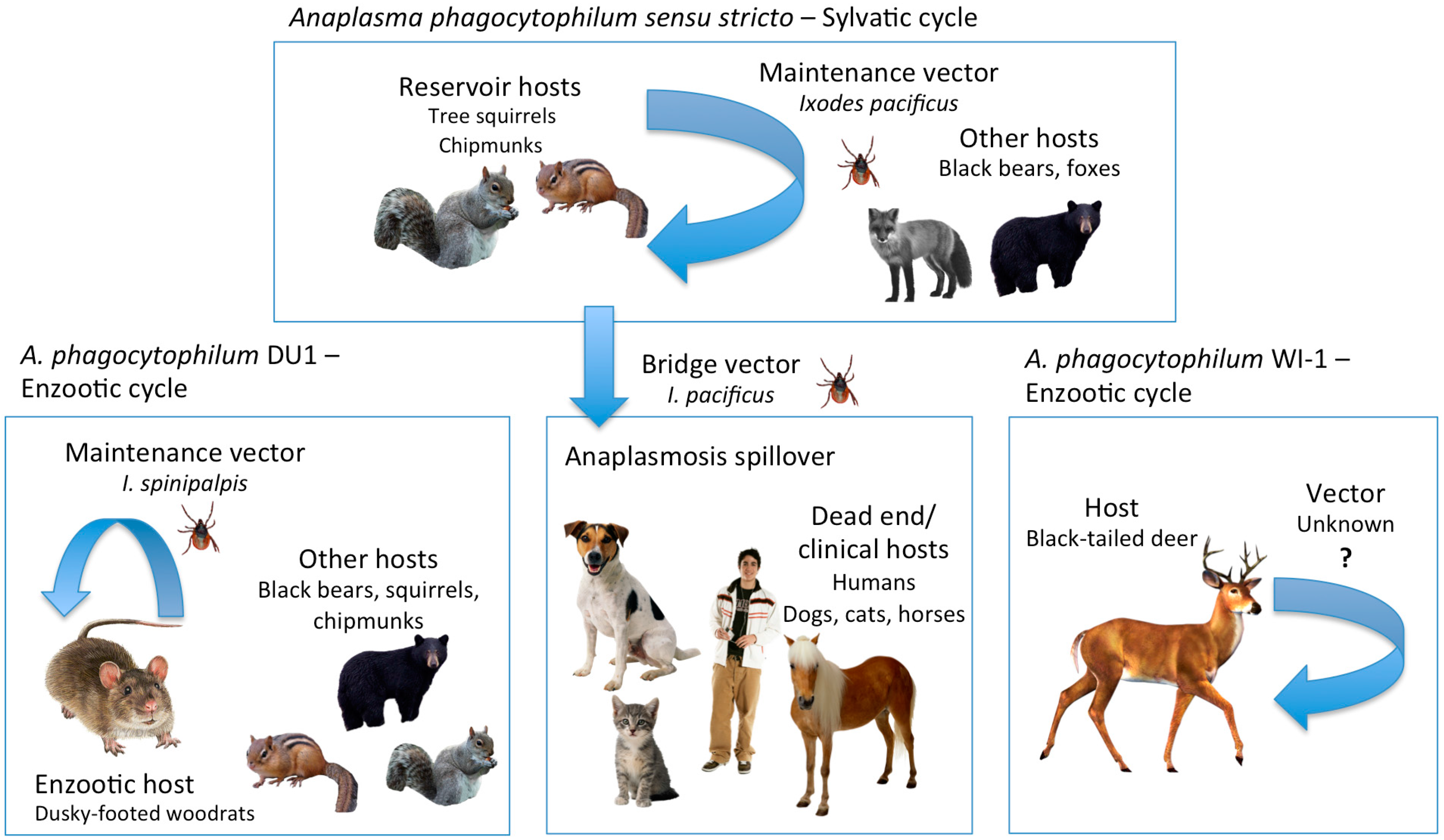


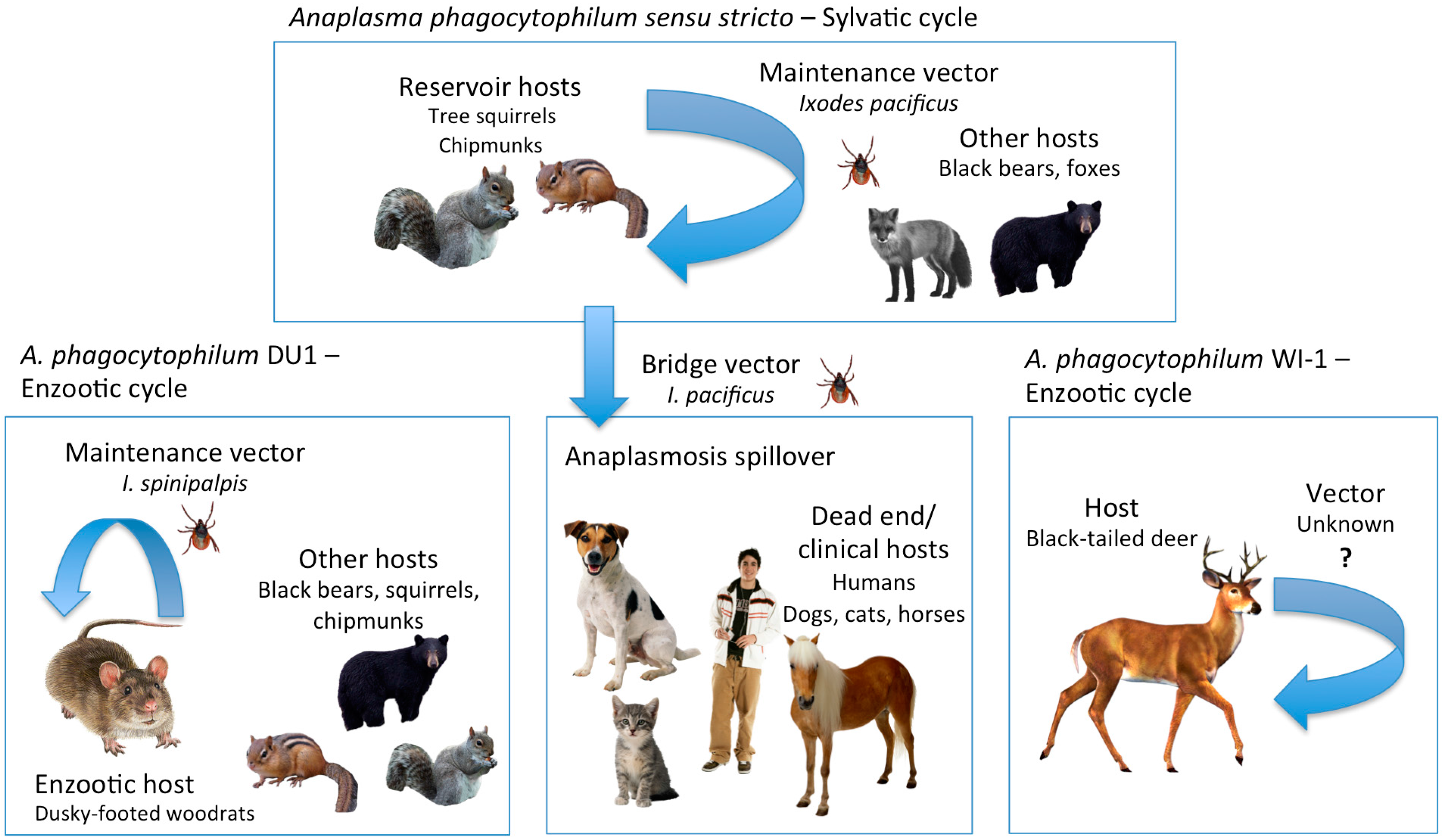{kind=link}
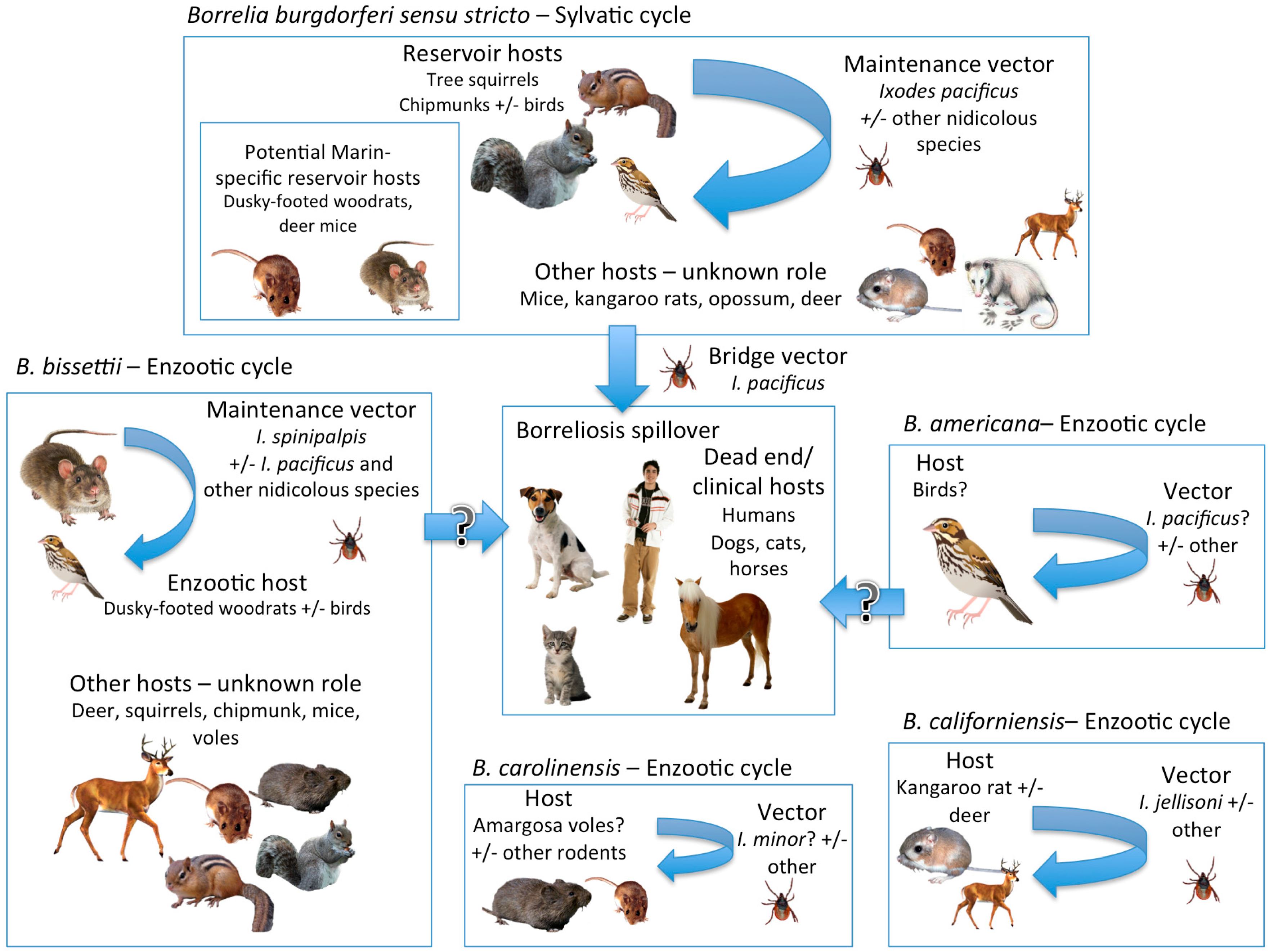{kind=link}
| Strain | Counties | Species (Host or Vector/Host) 1 | Number of Occurrences 2 | References |
|---|---|---|---|---|
| Sensu stricto | Alameda, El Dorado, Humboldt, Placer, Napa, Marin, Mendocino, Santa Cruz, Shasta, Yolo | Ursus americanus | 18 | [7,13,34,35] |
| Equus caballus | 10 | |||
| Canis lupus familiaris | 6 | |||
| Sciurus griseus | 4 | |||
| Tamias sonomae | 2 | |||
| T. ochrogenys | 2 | |||
| Tamiasciurus douglasii | 2 | |||
| S. carolinensis | 2 | |||
| Urocyon cinereoargenteus | 1 | |||
| Ixodes pacificus/flag | 4 | |||
| I. pacificus/Neotoma fuscipes | 1 | |||
| DU1 | Humboldt, Mendocino, Santa Cruz | N. fuscipes | 28 | [7,13,34,35] |
| U. americanus | 8 | |||
| T. ochrogenys | 4 | |||
| S. griseus | 1 | |||
| I. spinipalpis/N. fuscipes | 3 | |||
| I. angustus/N. fuscipes | 1 | |||
| I. woodi/N. fuscipes | 1 | |||
| I. ochotonae/Peromyscus sp. | 1 | |||
| I. pacificus/T. ochrogenys | 1 | |||
| I. pacificus/Homo sapiens | 1 | |||
| WI-1 | Mendocino, Mono, Tehama | Odocoileus hemionus | 16 | [36,37,38] |
| Lipoptena depressa/O. hemionus | 10 |
| Genospecies | Counties | Species (Host or Vector/Host) 1 | Number of Occurrences 2 | References |
|---|---|---|---|---|
| B. burgdorferi | Alameda, Butte, Contra Costa, El Dorado, Humboldt, Lake, Los Angeles, Marin, Mendocino, Napa, Placer, Plumas, Sacramento, San Mateo, Santa Clara, Santa Cruz, Shasta, Sonoma, Tehama, Trinity | Neotoma fuscipes | 95 | [17,20,27,28,29,41,43,44,45,46,47,48,49,50] 3 |
| Peromyscus maniculatus | 59 | |||
| Sciurus griseus | 38 | |||
| S. niger | 19 | |||
| Birds | 14 | |||
| Tamias senex | 4 | |||
| Tamiasciurus douglasii | 2 | |||
| T. ochrogenys | 2 | |||
| P. boylii | 2 | |||
| P. trueii | 2 | |||
| Dipodomys californicus | 1 | |||
| S. carolinensis | 1 | |||
| Didelphis virginiana | 1 | |||
| Odocoileus hemionus | 1 | |||
| Ixodes pacificus/flag | 1095 | |||
| I. pacificus/S. griseus | 25 | |||
| I. pacificus/birds | 3 | |||
| Dermacentor occidentalis/flag | 2 | |||
| I. auritulus/flag | 1 | |||
| I. spinipalpis/flag | 1 | |||
| I. jellisoni/flag | 1 | |||
| B. bissettii | Alameda, Contra Costa, Del Norte, Humboldt, Mendocino, San Bernardino, San Luis Obispo | N. fuscipes | 72 | [15,20,27,29,41,43,48,49,51,52,53,54] 2 |
| Birds | 17 | |||
| P. boylii | 10 | |||
| O. hemionus | 8 | |||
| P. trueii | 7 | |||
| P. maniculatus | 5 | |||
| T. siskiyou | 4 | |||
| N. lepida | 3 | |||
| R. rattus | 3 | |||
| M. californicus | 2 | |||
| T. douglasii | 1 | |||
| I. pacificus/flag | 49 | |||
| I. spinipalpis/flag | 12 | |||
| I. pacificus/bird | 5 | |||
| I. pacificus/S. griseus | 3 | |||
| I. spinipalpis/N. fuscipes | 2 | |||
| I. jellisoni/flag | 1 | |||
| I. auritulus/flag | 1 | |||
| B. californiensis | Alameda, Mendocino | D. californicus | 19 | [15,20,48] |
| O. hemionus | 3 | |||
| I. jellisoni/D. californicus | 1 | |||
| I. spinipalpis/D. californicus | 1 | |||
| I. pacificus/D. californicus | 1 | |||
| I. pacificus/flag | 1 | |||
| B. americana | Alameda, El Dorado, Los Angeles, Orange | I. pacificus/flag | 4 | [17,20,55] |
| B. carolinensis | Inyo | I. minor/Microtus californicus | 1 | [56] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stephenson, N.; Foley, J. Parallelisms and Contrasts in the Diverse Ecologies of the Anaplasma phagocytophilum and Borrelia burgdorferi Complexes of Bacteria in the Far Western United States. Vet. Sci. 2016, 3, 26. https://doi.org/10.3390/vetsci3040026
Stephenson N, Foley J. Parallelisms and Contrasts in the Diverse Ecologies of the Anaplasma phagocytophilum and Borrelia burgdorferi Complexes of Bacteria in the Far Western United States. Veterinary Sciences. 2016; 3(4):26. https://doi.org/10.3390/vetsci3040026
Chicago/Turabian StyleStephenson, Nicole, and Janet Foley. 2016. "Parallelisms and Contrasts in the Diverse Ecologies of the Anaplasma phagocytophilum and Borrelia burgdorferi Complexes of Bacteria in the Far Western United States" Veterinary Sciences 3, no. 4: 26. https://doi.org/10.3390/vetsci3040026
APA StyleStephenson, N., & Foley, J. (2016). Parallelisms and Contrasts in the Diverse Ecologies of the Anaplasma phagocytophilum and Borrelia burgdorferi Complexes of Bacteria in the Far Western United States. Veterinary Sciences, 3(4), 26. https://doi.org/10.3390/vetsci3040026