
Ovotransferrin as a Multifunctional Bioactive Protein: Unlocking Its Potential in Animal Health and Wellness



Simple Summary
Abstract
1. Introduction
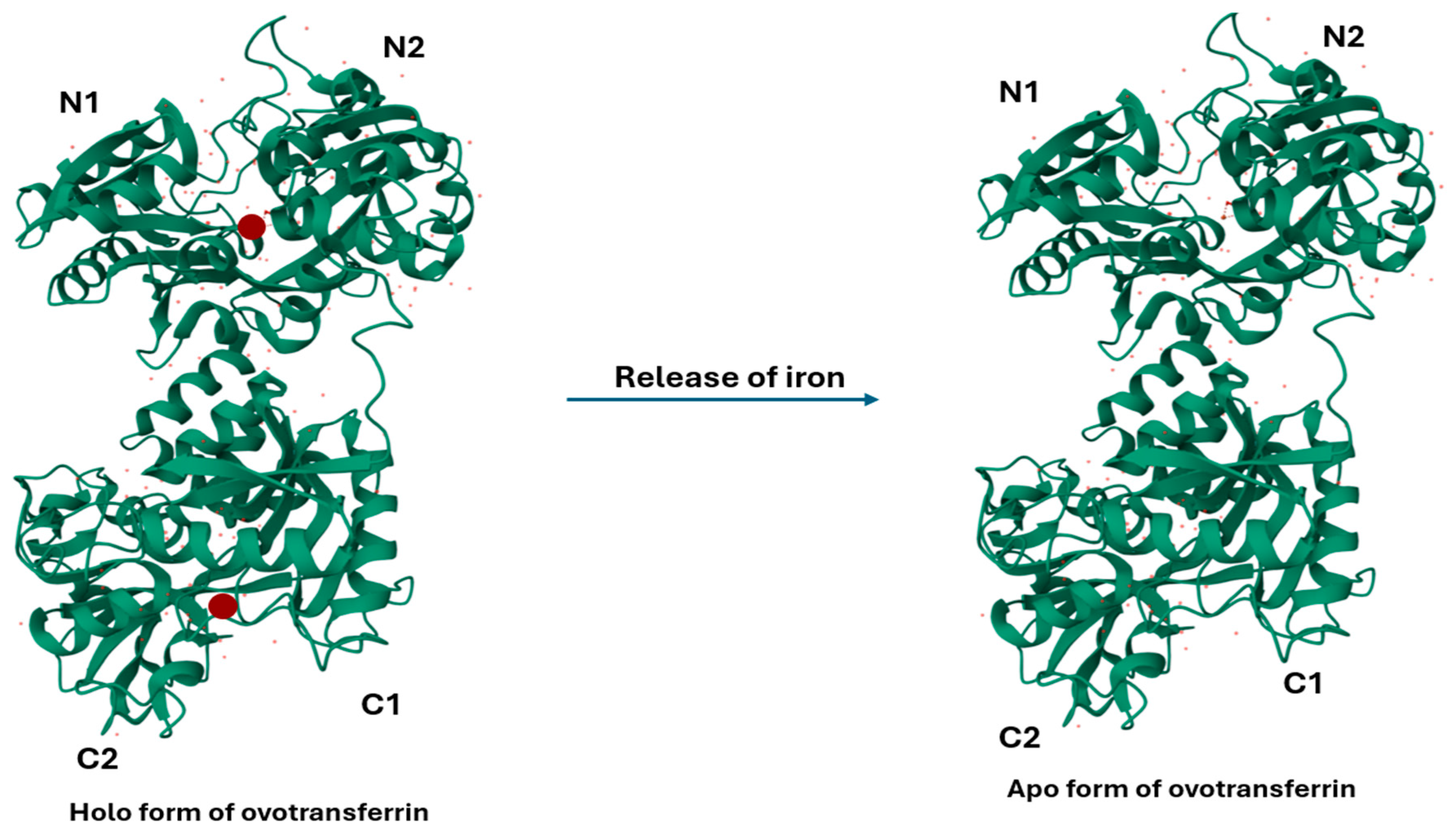
2. Structure of OVT
3. Stability and Bioavailability of OVT
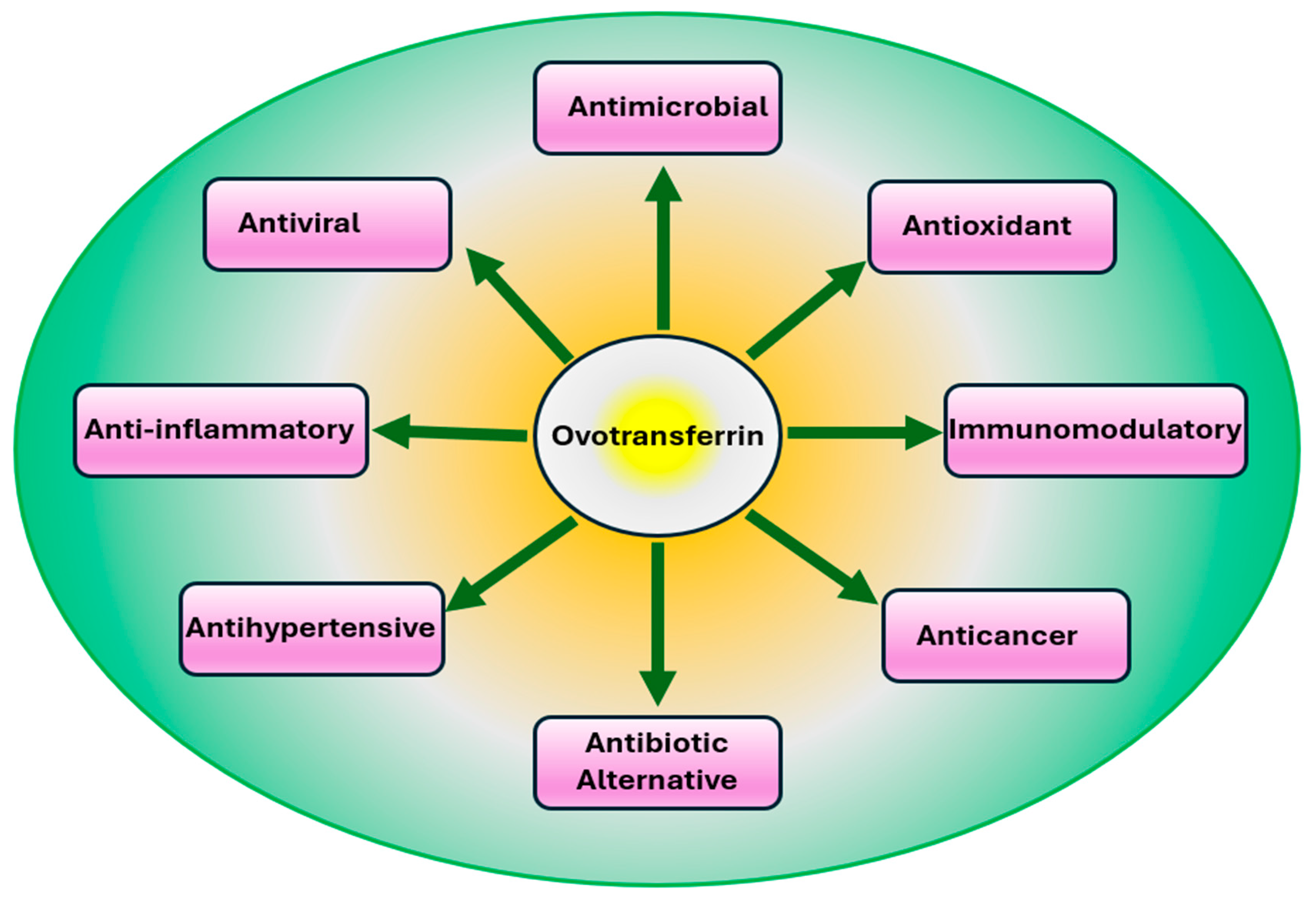
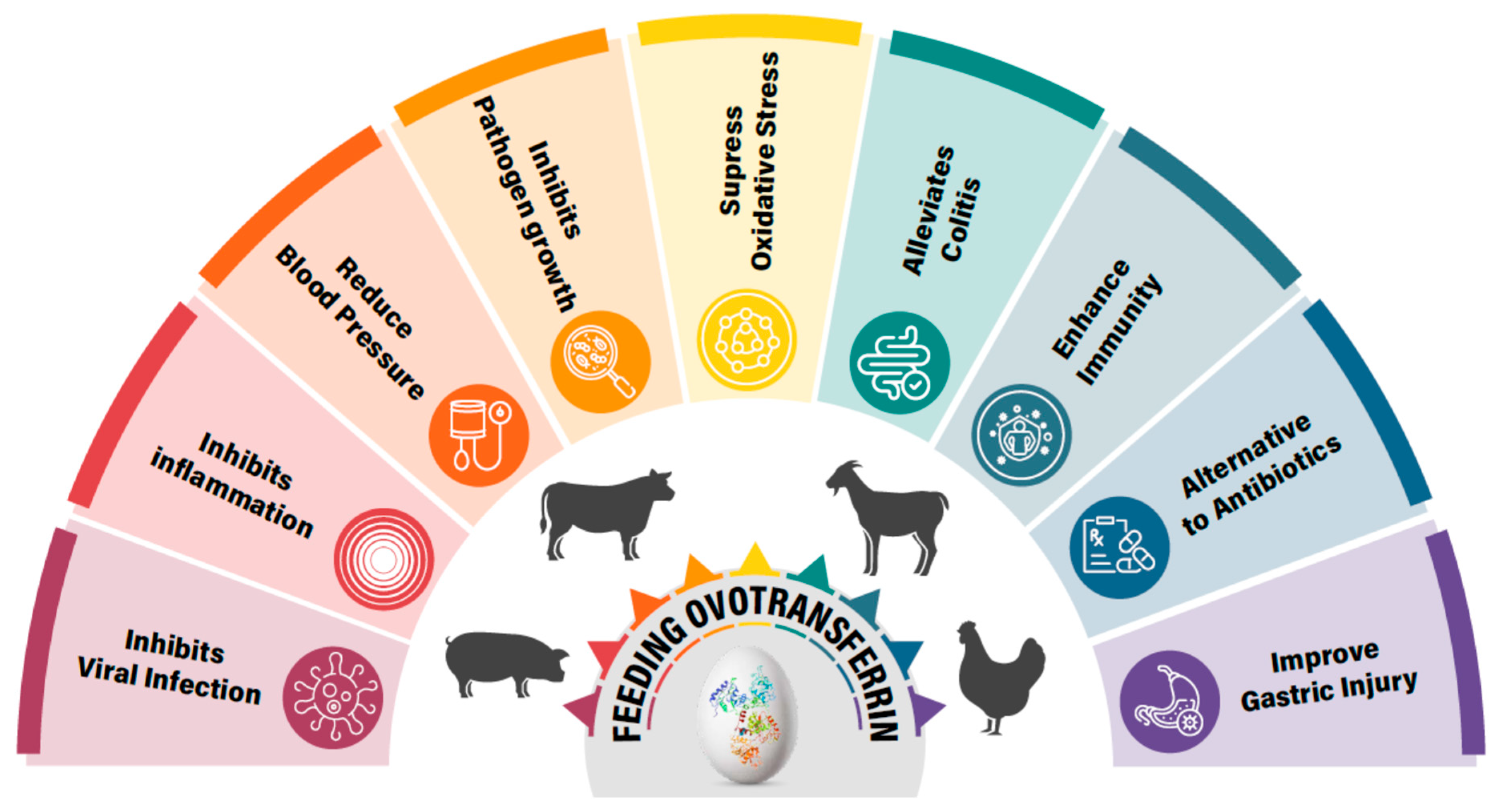
4. Physiological Properties of OVT
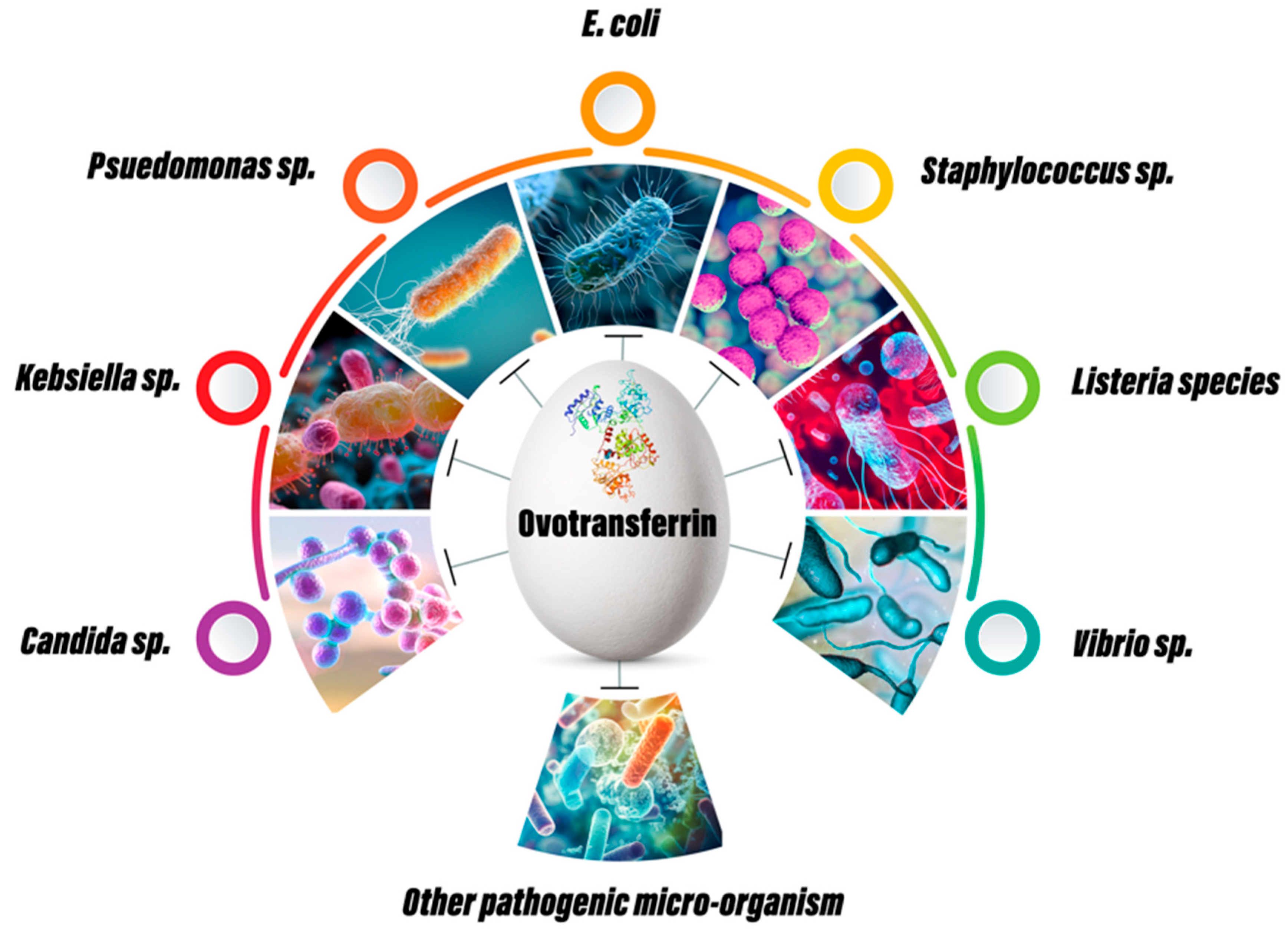
5. Antimicrobial Effects of OVT
5.1. In Vitro Studies of OVT
5.2. In Vivo Studies
6. Antiviral Properties of OVT
7. Antioxidant Properties of OVT
7.1. In Vitro Studies
7.2. In Vivo Studies
8. Anti-Inflammatory Properties of OVT
8.1. In Vitro Studies
8.2. In Vivo Studies
9. Immunomodulatory Effects of OVT
9.1. In Vitro Studies
9.2. In Vivo Studies
10. Others
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rehault-Godbert, S.; Guyot, N.; Nys, Y. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients 2019, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Liu, J.; Li, W.; Xu, G.; Yang, N. Divergent Proteome Patterns of Egg Albumen from Domestic Chicken, Duck, Goose, Turkey, Quail and Pigeon. Proteomics 2017, 17. [Google Scholar] [CrossRef]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, H.; Mikami, B.; Hirose, M. Crystal structure of diferric hen ovotransferrin at 2.4 A resolution. J. Mol. Biol. 1995, 254, 196–207. [Google Scholar] [CrossRef]
- Kurokawa, H.; Dewan, J.C.; Mikami, B.; Sacchettini, J.C.; Hirose, M. Crystal structure of hen apo-ovotransferrin. Both lobes adopt an open conformation upon loss of iron. J. Biol. Chem. 1999, 274, 28445–28452. [Google Scholar] [CrossRef]
- Williams, J.; Moreton, K.; Goodearl, A.D. Selective reduction of a disulphide bridge in hen ovotransferrin. Biochem. J. 1985, 228, 661–665. [Google Scholar] [CrossRef]
- Baringer, S.L.; Palsa, K.; Simpson, I.A.; Connor, J.R. Apo- and holo- transferrin differentially interact with ferroportin and hephaestin to regulate iron release at the blood-brain barrier. bioRxiv 2023. [Google Scholar] [CrossRef]
- Rathnapala, E.C.N.; Ahn, D.U.; Abeyrathne, E. Enzymatic Hydrolysis of Ovotransferrin and the Functional Properties of Its Hydrolysates. Food Sci. Anim. Resour. 2021, 41, 608–622. [Google Scholar] [CrossRef]
- Ko, K.Y.; Mendonca, A.F.; Ahn, D.U. Effect of ethylenediaminetetraacetate and lysozyme on the antimicrobial activity of ovotransferrin against Listeria monocytogenes. Poult. Sci. 2008, 87, 1649–1658. [Google Scholar] [CrossRef]
- Tan, A.T.; Woodworth, R.C. Ultraviolet difference spectrl studies of conalbumin complexes with transition metal ions. Biochemistry 1969, 8, 3711–3716. [Google Scholar] [CrossRef]
- Giansanti, F.; Leboffe, L.; Angelucci, F.; Antonini, G. The Nutraceutical Properties of Ovotransferrin and Its Potential Utilization as a Functional Food. Nutrients 2015, 7, 9105–9115. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Liu, Y.; Sun, J.; Jin, Y. Providing New Insights on the Molecular Properties and Thermal Stability of Ovotransferrin and Lactoferrin. Foods 2023, 12, 532. [Google Scholar] [CrossRef] [PubMed]
- Galla, R.; Grisenti, P.; Farghali, M.; Saccuman, L.; Ferraboschi, P.; Uberti, F. Ovotransferrin Supplementation Improves the Iron Absorption: An In Vitro Gastro-Intestinal Model. Biomedicines 2021, 9, 1543. [Google Scholar] [CrossRef]
- Azari, P.R.; Feeney, R.E. Resistance of metal complexes of conalbumin and transferrin to proteolysis and to thermal denaturation. J. Biol. Chem. 1958, 232, 293–302. [Google Scholar] [CrossRef]
- Bejjani, S.; Wu, J. Transport of IRW, an ovotransferrin-derived antihypertensive peptide, in human intestinal epithelial Caco-2 cells. J. Agric. Food Chem. 2013, 61, 1487–1492. [Google Scholar] [CrossRef]
- Majumder, K.; Liang, G.; Chen, Y.; Guan, L.; Davidge, S.T.; Wu, J. Egg ovotransferrin-derived ACE inhibitory peptide IRW increases ACE2 but decreases proinflammatory genes expression in mesenteric artery of spontaneously hypertensive rats. Mol. Nutr. Food Res. 2015, 59, 1735–1744. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Huang, C.; Cai, B.; Guo, F.; Chen, L.; Feng, X.; Ma, M. Improved thermal tolerance of ovotransferrin against pasteurization by phosphorylation. Food Chem. 2023, 405, 135019. [Google Scholar] [CrossRef]
- Pan, F.; Wu, X.; Gong, L.; Xu, H.; Yuan, Y.; Lu, J.; Zhang, T.; Liu, J.; Shang, X. Dextran sulfate acting as a chaperone-like component on inhibition of amorphous aggregation and enhancing thermal stability of ovotransferrin. Food Chem. 2024, 445, 138720. [Google Scholar] [CrossRef]
- Shirkhani, R.; Lee, E.J.; Talukder, J. Mechanism of Absorption and Transportation of Ovotransferrin in the Intestine. Faseb J. 2018, 32, 747.17. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Sugimoto, Y.; Aoki, T. Ovotransferrin antimicrobial peptide (OTAP-92) kills bacteria through a membrane damage mechanism. Biochim. Biophys. Acta 2000, 1523, 196–205. [Google Scholar] [CrossRef]
- Aguilera, O.; Quiros, L.M.; Fierro, J.F. Transferrins selectively cause ion efflux through bacterial and artificial membranes. FEBS Lett. 2003, 548, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Baron, F.; Jan, S.; Gonnet, F.; Pasco, M.; Jardin, J.; Giudici, B.; Gautier, M.; Guerin-Dubiard, C.; Nau, F. Ovotransferrin plays a major role in the strong bactericidal effect of egg white against the Bacillus cereus group. J. Food Prot. 2014, 77, 955–962. [Google Scholar] [CrossRef]
- Ma, B.; Chen, Y.; Hu, G.; Zeng, Q.; Lv, X.; Oh, D.H.; Fu, X.; Jin, Y. Ovotransferrin Antibacterial Peptide Coupling Mesoporous Silica Nanoparticle as an Effective Antibiotic Delivery System for Treating Bacterial Infection In Vivo. ACS Biomater. Sci. Eng. 2022, 8, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Legros, J.; Jan, S.; Bonnassie, S.; Gautier, M.; Croguennec, T.; Pezennec, S.; Cochet, M.F.; Nau, F.; Andrews, S.C.; Baron, F. The Role of Ovotransferrin in Egg-White Antimicrobial Activity: A Review. Foods 2021, 10, 823. [Google Scholar] [CrossRef]
- Valenti, P.; Visca, P.; Antonini, G.; Orsi, N. Antifungal activity of ovotransferrin towards genus Candida. Mycopathologia 1985, 89, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Beeckman, D.S.; Van Droogenbroeck, C.M.; De Cock, B.J.; Van Oostveldt, P.; Vanrompay, D.C. Effect of ovotransferrin and lactoferrins on Chlamydophila psittaci adhesion and invasion in HD11 chicken macrophages. Vet. Res. 2007, 38, 729–739. [Google Scholar] [CrossRef]
- Vandeputte, M.; Verhaeghe, M.; Willocx, L.; Bossier, P.; Vanrompay, D. Bovine Lactoferrin and Hen Ovotransferrin Affect Virulence Factors of Acute Hepatopancreatic Necrosis Disease (AHPND)-Inducing Vibrio parahaemolyticus Strains. Microorganisms 2023, 11, 2912. [Google Scholar] [CrossRef]
- Van Droogenbroeck, C.; Vanrompay, D. Use of ovotransferrin on a turkey farm to reduce respiratory disease. Vet. Rec. 2013, 172, 71. [Google Scholar] [CrossRef]
- Giansanti, F.; Rossi, P.; Massucci, M.T.; Botti, D.; Antonini, G.; Valenti, P.; Seganti, L. Antiviral activity of ovotransferrin discloses an evolutionary strategy for the defensive activities of lactoferrin. Biochem. Cell Biol. 2002, 80, 125–130. [Google Scholar] [CrossRef]
- Zhou, Y.; Qun, T.; Huaying, D.; Yonggang, T.; Shaofu, W.; Wenjun, W.; Xu, M. Antiviral effect of ovotransferrin in mouse peritoneal macrophages by up-regulating type I interferon expression. Food Agric. Immunol. 2018, 29, 600–614. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Hoq, M.I.; Aoki, T. Ovotransferrin possesses SOD-like superoxide anion scavenging activity that is promoted by copper and manganese binding. Int. J. Biol. Macromol. 2007, 41, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Majumder, K.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Structure and activity study of egg protein ovotransferrin derived peptides (IRW and IQW) on endothelial inflammatory response and oxidative stress. J. Agric. Food Chem. 2013, 61, 2120–2129. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Moon, S.H.; Kim, H.S.; Park, E.; Ahn, D.U.; Paik, H.D. Antioxidant and anticancer effects of functional peptides from ovotransferrin hydrolysates. J. Sci. Food Agric. 2017, 97, 4857–4864. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Moon, S.H.; Ahn, D.U.; Paik, H.D.; Park, E. Antioxidant effects of ovotransferrin and its hydrolysates. Poult. Sci. 2012, 91, 2747–2754. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, S.; Yao, Y.; Wu, N.; Xu, M.; Du, H.; Yin, Z.; Zhao, Y.; Tu, Y. Ovotransferrin Inhibits TNF-alpha Induced Inflammatory Response in Gastric Epithelial Cells via MAPK and NF-kappaB Pathway. J. Agric. Food Chem. 2023, 71, 12474–12486. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Rupa, P.; Kovacs-Nolan, J.; Turner, P.V.; Matsui, T.; Mine, Y. Oral administration of hen egg white ovotransferrin attenuates the development of colitis induced by dextran sodium sulfate in mice. J. Agric. Food Chem. 2015, 63, 1532–1539. [Google Scholar] [CrossRef]
- Chai, Y.; Ding, S.; Jiang, L.; Wang, S.; Yuan, X.; Jiang, H.; Fang, J. The mitigative effect of ovotransferrin-derived peptide IQW on DSS-induced colitis via alleviating intestinal injury and reprogramming intestinal microbes. Front. Nutr. 2022, 9, 927363. [Google Scholar] [CrossRef]
- Ru, Z.; Xu, M.; Zhu, G.; Tu, Y.; Jiang, Y.; Du, H. Ovotransferrin exerts bidirectional immunomodulatory activities via TLR4-mediated signal transduction pathways in RAW264.7 cells. Food Sci. Nutr. 2021, 9, 6162–6175. [Google Scholar] [CrossRef]
- Liu, L.; Xu, M.; Tu, Y.; Du, H.; Zhou, Y.; Zhu, G. Immunomodulatory effect of protease hydrolysates from ovotransferrin. Food Funct. 2017, 8, 1452–1459. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, H.J.; Ahn, D.U.; Paik, H.D. Improved immune-enhancing activity of egg white protein ovotransferrin after enzyme hydrolysis. J. Anim. Sci. Technol. 2021, 63, 1159–1168. [Google Scholar] [CrossRef]
- Zhu, G.; Luo, J.; Du, H.; Jiang, Y.; Tu, Y.; Yao, Y.; Xu, M. Ovotransferrin enhances intestinal immune response in cyclophosphamide-induced immunosuppressed mice. Int. J. Biol. Macromol. 2018, 120, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shang, N.; Wu, J. Egg White Ovotransferrin Attenuates RANKL-Induced Osteoclastogenesis and Bone Resorption. Nutrients 2019, 11, 2254. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Kiyono, T. Novel anticancer activity of the autocleaved ovotransferrin against human colon and breast cancer cells. J. Agric. Food Chem. 2009, 57, 11383–11390. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Lee, J.H.; Kim, J.H.; Paik, H.D.; Ahn, D.U. In vitro cytotoxic and ACE-inhibitory activities of promod 278P hydrolysate of ovotransferrin from chicken egg white. Poult. Sci. 2017, 96, 1982–1987. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, S.; Yao, Y.; Wu, N.; Xu, M.; Du, H.; Zhao, Y.; Tu, Y. Ovotransferrin alleviated acute gastric mucosal injury in BALB/c mice caused by ethanol. Food Funct. 2023, 14, 305–318. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G.; Von Hunolstein, C.; Visca, P.; Orsi, N.; Antonini, E. Studies of the antimicrobial activity of ovotransferrin. Int. J. Tissue React. 1983, 5, 97–105. [Google Scholar]
- Ko, K.Y.; Mendoncam, A.F.; Ismail, H.; Ahn, D.U. Ethylenediaminetetraacetate and lysozyme improves antimicrobial activities of ovotransferrin against Escherichia coli O157:H7. Poult. Sci. 2009, 88, 406–414. [Google Scholar] [CrossRef]
- Moon, S.H.; Paik, H.D.; White, S.; Daraba, A.; Mendonca, A.F.; Ahn, D.U. Influence of nisin and selected meat additives on the antimicrobial effect of ovotransferrin against Listeria monocytogenes. Poult. Sci. 2011, 90, 2584–2591. [Google Scholar] [CrossRef]
- Giansanti, F.; Massucci, M.T.; Giardi, M.F.; Nozza, F.; Pulsinelli, E.; Nicolini, C.; Botti, D.; Antonini, G. Antiviral activity of ovotransferrin derived peptides. Biochem. Biophys. Res. Commun. 2005, 331, 69–73. [Google Scholar] [CrossRef]
- Moon, S.H.; Lee, J.H.; Ahn, D.U.; Paik, H.D. In vitro antioxidant and mineral-chelating properties of natural and autocleaved ovotransferrin. J. Sci. Food Agric. 2015, 95, 2065–2070. [Google Scholar] [CrossRef]
- You, J.; Luo, Y.; Wu, J. Conjugation of ovotransferrin with catechin shows improved antioxidant activity. J. Agric. Food Chem. 2014, 62, 2581–2587. [Google Scholar] [CrossRef]
- Huang, W.; Shen, S.; Nimalaratne, C.; Li, S.; Majumder, K.; Wu, J. Effects of addition of egg ovotransferrin-derived peptides on the oxygen radical absorbance capacity of different teas. Food Chem. 2012, 135, 1600–1607. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Zhang, Q.; Lin, Y.; Gao, Y.; Zhang, P. The Ovotransferrin-Derived Peptide IRW Attenuates Lipopolysaccharide-Induced Inflammatory Responses. Biomed. Res. Int. 2019, 2019, 8676410. [Google Scholar] [CrossRef] [PubMed]
- Chiurciu, C.; Chiurciu, V.; Oporanu, M.; Patrascu, I.V.; Mihai, I.; Tablica, M.; Cristina, R.T. PC2 Ovotransferrin: Characterization and Alternative Immunotherapeutic Activity. Evid. Based Complement. Alternat Med. 2017, 2017, 8671271. [Google Scholar] [CrossRef]
- Zhu, G.; Jiang, Y.; Yao, Y.; Wu, N.; Luo, J.; Hu, M.; Tu, Y.; Xu, M. Ovotransferrin ameliorates the dysbiosis of immunomodulatory function and intestinal microbiota induced by cyclophosphamide. Food Funct. 2019, 10, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Lee, J.H.; Lee, Y.J.; Chang, K.H.; Paik, J.Y.; Ahn, D.U.; Paik, H.D. Screening for cytotoxic activity of ovotransferrin and its enzyme hydrolysates. Poult. Sci. 2013, 92, 424–434. [Google Scholar] [CrossRef]
- Lee, N.Y.; Cheng, J.T.; Enomoto, T.; Nakano, Y. One peptide derived from hen ovotransferrin as pro-drug to inhibit angiotensin converting enzyme. J. Food Drug Anal. 2006, 14, 31–35. [Google Scholar] [CrossRef]
- Hernandez-Patlan, D.; Tellez-Isaias, G.; Hernandez-Velasco, X.; Solis-Cruz, B. Editorial: Technological strategies to improve animal health and production. Front. Vet. Sci. 2023, 10, 1206170. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects of OVT | Model | Dose | Reference |
|---|---|---|---|
| Anti-microbial properties | |||
| Killed bacteria by permeation | E. coli | 2 μM | [20] |
| Permeation of K+ ion in the bacterial cell membrane | E coli | 13 μM | [21] |
| Provoked perturbation of the electrochemical potential of the cytoplasmic membrane | Bacillus cereus group | 13 g/L | [22] |
| OVT-derived nanoparticles interacted with bacteria | E. coli | 20 mg/kg | [23] |
| Induced outer membrane permeabilization | Salmonella enteritidis | 13 g/L | [24] |
| Interacted with fungal surface proteins and inhibited growth | Candida albicans and CC. krusei | 1 mg/mL | [25] |
| Inhibited infectivity, adhesion, and invasion in a dose-dependent manner | Chlamydophila Psittaci | 0.5–5 mg/mL | [26] |
| Prolonged the lag phase of bacteria | Vibrio parahaemolyticus | 1–10 mg/mL | [27] |
| Reduced respiratory disease as aerosol | Chlamydia psittaci | 5 mg/animal | [28] |
| Antiviral properties | |||
| Inhibited viral antigen synthesis | HSV-1 and MDV | <3 mg/mL | [29] |
| Downregulated RNF125 and upregulated RIG-I expression by NF-κB; results increased IFN-1 expression in macrophages | VSV | 100 ng/ml | [30] |
| Antioxidant properties | |||
| Acted as SOD mimic protein and scavenged superoxide anion (O2−) | Xanthine/xanthine oxidase coupling system | 6.4 μM | [31] |
| OVT-derived peptides reduced TNF-induced superoxide generation | Endothelial cells | 50 μmol/L | [32] |
| OVT hydrolysates showed strong free radical scavenging activity | Radical scavenging assay | 0.5–2 mg/mL | [33] |
| OVT and OH-OVT protected against oxidative stress-induced DNA damage | Human leukocytes | 500 μg/mL | [34] |
| Anti-inflammatory effects | |||
| Inhibited MAPK/NF-κB Pathway | Gastric epithelial cells | 50−400 μg/mL | [35] |
| Prevented DSS-induced colitis by inhibiting epithelial dysfunction and modulating cytokine profile | BALB/c mice | 50 or 250 mg/kg BW/day) | [36] |
| OVT-derived peptide (IQW) mitigated disulfide sodium-induced colitis | C57BL/6J mice | 20–100 μg/mL of IQW | [37] |
| Immunomodulatory effects | |||
| Regulated TLR4-mediated NF-κB/MAPK signaling | RAW 264.7 Ms. macrophage cells | 50 μg/mL | [38] |
| OVT-hydrolysates inhibited DC maturation by reducing MHC-II, TNF-α, IL-12p70, and RANTES | Bone marrow-derived dendritic cells | 250 μg/mL | [39] |
| OVT hydrolysate promoted phagocytic activity by inducing NO and iNOS | RAW 264.7 macrophages | 250–500 μg/mL | [40] |
| Promoted maturation of intestinal DC and increased cytokine expression | Kunming mouse | 2–200 mg/kg | [41] |
| Other properties | |||
| Induced apoptosis of mature osteoclasts by increasing expression of Bim and Bad | Mouse osteoclast | 1–1000 μg/mL | [42] |
| OVT hydrolysates exhibited cytotoxic properties against cancer cells | Human cancer cell lines | 10 mg/mL | [33] |
| Autocleaved OVT dissipated mitochondrial membrane potential and activation of caspase-9/-6 | HCT-116 and MCF-7 | 250 μg/mL | [43] |
| OVT hydrolysates acted as antihypertension agents by suppressing ACE | Human cancer cell lines | IC50: 1.53 ± 0.20 mg/mL | [44] |
| OVT-derived IRW peptide inhibited blood pressure through ACE reduction | Spontaneously hypertensive rats | 3 and 15 mg/kg | [44] |
| Reduced gastric disease caused by Helicobacter pylori | BALB/c mice | 50−400 μg/mL | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasad, S.; Patel, B.; Kumar, P.; Kaufman, J.; Lall, R. Ovotransferrin as a Multifunctional Bioactive Protein: Unlocking Its Potential in Animal Health and Wellness. Vet. Sci. 2025, 12, 514. https://doi.org/10.3390/vetsci12060514
Prasad S, Patel B, Kumar P, Kaufman J, Lall R. Ovotransferrin as a Multifunctional Bioactive Protein: Unlocking Its Potential in Animal Health and Wellness. Veterinary Sciences. 2025; 12(6):514. https://doi.org/10.3390/vetsci12060514
Chicago/Turabian StylePrasad, Sahdeo, Bhaumik Patel, Prafulla Kumar, Jeffrey Kaufman, and Rajiv Lall. 2025. "Ovotransferrin as a Multifunctional Bioactive Protein: Unlocking Its Potential in Animal Health and Wellness" Veterinary Sciences 12, no. 6: 514. https://doi.org/10.3390/vetsci12060514
APA StylePrasad, S., Patel, B., Kumar, P., Kaufman, J., & Lall, R. (2025). Ovotransferrin as a Multifunctional Bioactive Protein: Unlocking Its Potential in Animal Health and Wellness. Veterinary Sciences, 12(6), 514. https://doi.org/10.3390/vetsci12060514