
Polyhexamethylene Biguanide Nanoparticles Inhibit Biofilm Formation by Mastitis-Causing Staphylococcus aureus
 , ,
, ,  ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthesis of Polyhexamethylene Biguanide Nanoparticles
2.2. Selection and Identification of Staphylococcus aureus Isolates
2.3. Antimicrobial Susceptibility
2.4. Intracellular Infection of MAC-T Cells
2.5. Biofilm-Forming Ability
2.6. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
Toxicity to Bovine Mammary Epithelial Cells
2.7. Experiment 1: Evaluation of PHMB NP Toxicity and Antibiofilm Activities
2.7.1. Biofilm Inhibition
2.7.2. Evaluation of Preformed Biofilm
2.8. Experiment 2: Evaluation of the Antimicrobial Activity of PHMB NPs on Teat Skin
Antimicrobial Activity of PHMB NPs Using the Excised Teat Model
2.9. Statistical Analyses
3. Results
3.1. Nanoparticle Characteristics
3.2. Antimicrobial Susceptibility Results
3.3. Intracellular Infection Results
3.4. Biofilm-Forming Ability Results
3.5. Minimum Inhibitory Concentration and Minimum Bactericidal Concentration Results
3.6. Experiment 1: Evaluation of PHMB NP Toxicity and Antibiofilm Activities Results
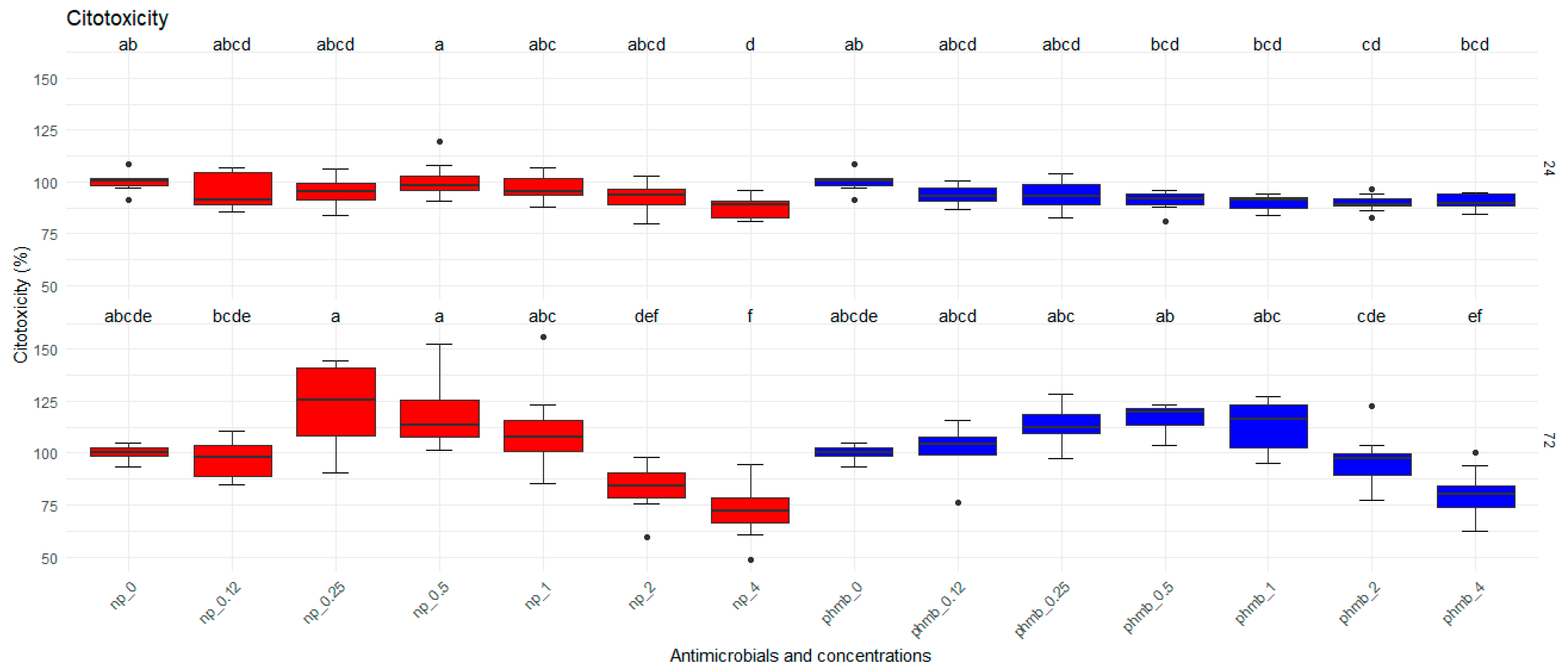
3.6.1. Toxicity to Bovine Mammary Epithelial Cells
3.6.2. Evaluation of PHMB NPs and PHMB on Biofilm Inhibition
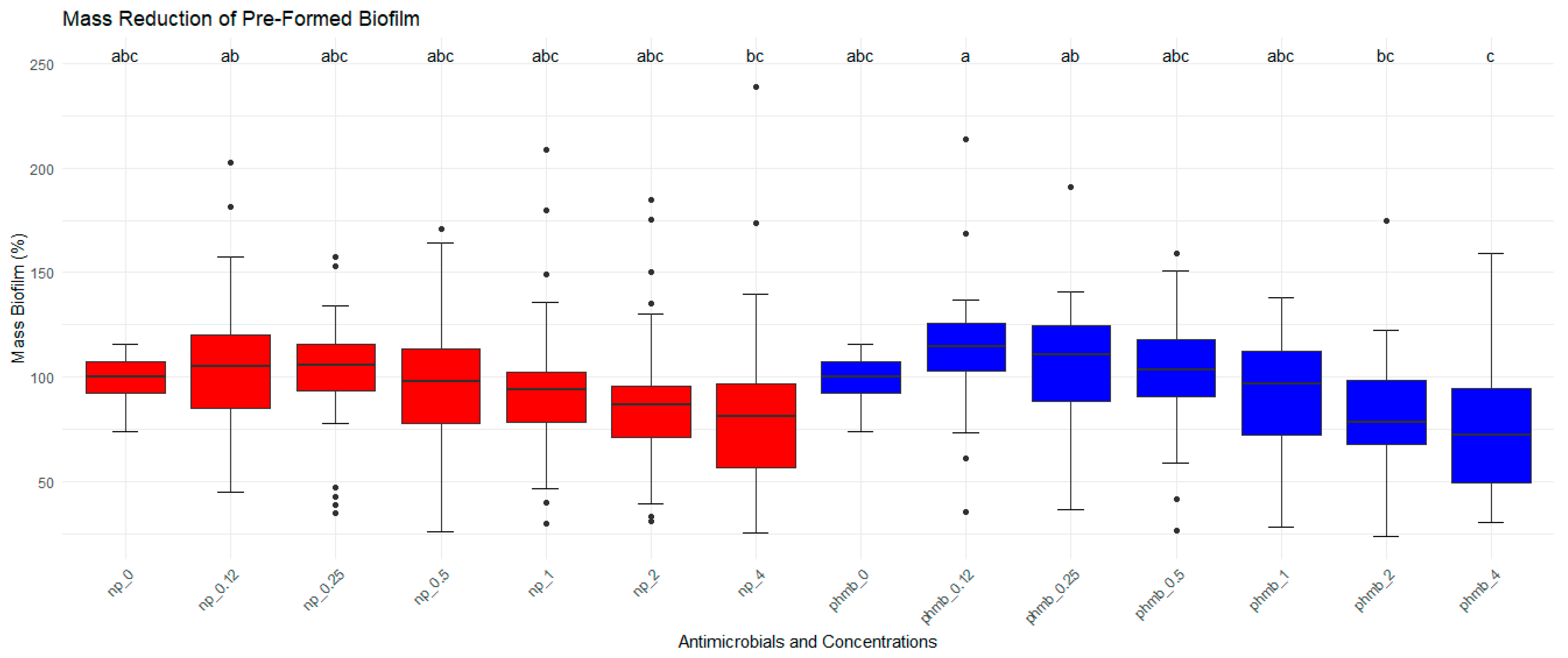
3.6.3. Evaluation of PHMB NPs and PHMB on Preformed Biofilm
3.7. Experiment 2: Evaluation of the Antimicrobial Activity of PHMB NPs on Teat Skin Results
Antimicrobial Activity of PHMB NPs and PHMB Using the Excised Teat Model
4. Discussion
4.1. Experiment 1: Evaluation of PHMB NP Toxicity and Antibiofilm Activities
4.2. Experiment 2: Evaluation of the Antimicrobial Activity of PHMB NPs on Teat Skin
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMR | Antimicrobial resistance |
| ATCC | American Type Culture Collection |
| CHG | Chlorhexidine digluconate |
| CIA | Critically important antimicrobials |
| CV | Crystal violet |
| DLS | Dynamic light scattering |
| EUCAST | European Committee on Antimicrobial Susceptibility Testing |
| IMI | Intramammary infections |
| LMM | Linear mixed-effects model |
| MAC-T cells | Bovine mammary epithelial cells |
| MALDI-TOF MS | Matrix-assisted laser desorption/ionization time-of-flight |
| MBC | Minimum bactericidal concentration |
| MDR | Multidrug-resistant |
| MG | Mammary gland |
| MHB | Mueller–Hinton broth |
| MIC | Minimal inhibitory concentrations |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| MTT | Thiazolyl Blue Tetrazolium Bromide |
| NaDCC | Sodium dichloroisocyanurate |
| NPs | Nanoparticles |
| OD | Optical density |
| ODNC | Optical density of negative control |
| PHMB | Polyhexamethylene biguanide |
| PVP-I | Povidone iodine |
| TSB | Tryptic soy broth |
References
- Pedersen, R.R.; Krömker, V.; Bjarnsholt, T.; Dahl-Pedersen, K.; Buhl, R.; Jørgensen, E. Biofilm Research in Bovine Mastitis. Front. Vet. Sci. 2021, 8, 656810. [Google Scholar] [CrossRef] [PubMed]
- van Soest, F.J.S.; Santman-Berends, I.M.G.A.; Lam, T.J.G.M.; Hogeveen, H. Failure and Preventive Costs of Mastitis on Dutch Dairy Farms. J. Dairy Sci. 2016, 99, 8365–8374. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, A.A.; Rocha, C.M.B.M.; Bruhn, F.R.P.; Custódio, D.A.C.; Braz, M.S.; Pinto, S.M.; Silva, D.B.; Costa, G.M. Staphylococcus aureus and Streptococcus agalactiae: Prevalence, Resistance to Antimicrobials, and Their Relationship with the Milk Quality of Dairy Cattle Herds in Minas Gerais State, Brazil. Pesqui. Veterinária Bras. 2019, 39, 308–316. [Google Scholar] [CrossRef]
- De Buck, J.; Ha, V.; Naushad, S.; Nobrega, D.B.; Luby, C.; Middleton, J.R.; De Vliegher, S.; Barkema, H.W. Non-Aureus Staphylococci and Bovine Udder Health: Current Understanding and Knowledge Gaps. Front. Vet. Sci. 2021, 8, 658031. [Google Scholar] [CrossRef]
- Barkema, H.W.; Schukken, Y.H.; Zadoks, R.N. Invited Review: The Role of Cow, Pathogen, and Treatment Regimen in the Therapeutic Success of Bovine Staphylococcus aureus Mastitis. J. Dairy Sci. 2006, 89, 1877–1895. [Google Scholar] [CrossRef]
- Kümmel, J.; Stessl, B.; Gonano, M.; Walcher, G.; Bereuter, O.; Fricker, M.; Grunert, T.; Wagner, M.; Ehling-Schulz, M. Staphylococcus aureus Entrance into the Dairy Chain: Tracking S. aureus from Dairy Cow to Cheese. Front. Microbiol. 2016, 7, 1603. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis Detection, Management, and Prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.H.; Lacy-Hulbert, S.J. Effect of Disinfecting Teats Post-Milking or Pre- and Post-Milking on Intramammary Infection and Somatic Cell Count. N. Z. Vet. J. 2013, 61, 262–268. [Google Scholar] [CrossRef]
- Fitzpatrick, S.R.; Garvey, M.; Flynn, J.; Jordan, K.; Gleeson, D. Are Some Teat Disinfectant Formulations More Effective Against Specific Bacteria Isolated on Teat Skin than Others? Acta Vet. Scand. 2019, 61, 21. [Google Scholar] [CrossRef]
- Neijenhuis, F.; Klungel, G.H.; Hogeveen, H. Recovery of Cow Teats after Milking as Determined by Ultrasonographic Scanning. J. Dairy Sci. 2001, 84, 2599–2606. [Google Scholar] [CrossRef]
- Berg, W.; Rose-Meierhöfer, S.; Ammon, C.; Kobbe, C. Short Communication: Dipping Efficiency and Teat Dip Residues in Milk Using an Automatic Dipping System. J. Dairy Sci. 2014, 97, 3689–3693. [Google Scholar] [CrossRef] [PubMed]
- Côté-Gravel, J.; Malouin, F. Symposium Review: Features of Staphylococcus Aureus Mastitis Pathogenesis That Guide Vaccine Development Strategies. J. Dairy Sci. 2019, 102, 4727–4740. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, Invasion and Evasion: The Many Functions of the Surface Proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef]
- Kamaruzzaman, N.F.; Chong, S.Q.Y.; Edmondson-Brown, K.M.; Ntow-Boahene, W.; Bardiau, M.; Good, L. Bactericidal and Anti-Biofilm Effects of Polyhexamethylene Biguanide in Models of Intracellular and Biofilm of Staphylococcus aureus Isolated from Bovine Mastitis. Front. Microbiol. 2017, 8, 1518. [Google Scholar] [CrossRef] [PubMed]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus Biofilms: Recent Developments in Biofilm Dispersal. Front. Cell Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef]
- Malaekeh-Nikouei, B.; Fazly Bazzaz, B.S.; Mirhadi, E.; Tajani, A.S.; Khameneh, B. The Role of Nanotechnology in Combating Biofilm-Based Antibiotic Resistance. J. Drug Deliv. Sci. Technol. 2020, 60, 101880. [Google Scholar] [CrossRef]
- Hensen, S.M.; Pavičić, M.J.A.M.P.; Lohuis, J.A.C.M.; De Hoog, J.A.M.; Poutrel, B. Location of Staphylococcus aureus within the Experimentally Infected Bovine Udder and the Expression of Capsular Polysaccharide Type 5 in Situ. J. Dairy Sci. 2000, 83, 1966–1975. [Google Scholar] [CrossRef] [PubMed]
- Schönborn, S.; Krömker, V. Detection of the Biofilm Component Polysaccharide Intercellular Adhesin in Staphylococcus aureus Infected Cow Udders. Vet. Microbiol. 2016, 196, 126–128. [Google Scholar] [CrossRef]
- El-Sayed, A.; Kamel, M. Bovine Mastitis Prevention and Control in the Post-Antibiotic Era. Trop. Anim. Health Prod. 2021, 53, 236. [Google Scholar] [CrossRef]
- Buzea, C.; Pacheco, I.I.; Robbie, K. Nanomaterials and Nanoparticles: Sources and Toxicity. Biointerphases 2007, 2, MR17–MR71. [Google Scholar] [CrossRef]
- Bai, X.; Gao, Y.; Liu, H.G.; Zheng, L. Synthesis of Amphiphilic Ionic Liquids Terminated Gold Nanorods and Their Superior Catalytic Activity for the Reduction of Nitro Compounds. J. Phys. Chem. C 2009, 113, 17730–17736. [Google Scholar] [CrossRef]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of Nanoparticle Design for Overcoming Biological Barriers to Drug Delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef]
- Seleem, M.N.; Munusamy, P.; Ranjan, A.; Alqublan, H.; Pickrell, G.; Sriranganathan, N. Silica-Antibiotic Hybrid Nanoparticles for Targeting Intracellular Pathogens. Antimicrob. Agents Chemother. 2009, 53, 4270–4274. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, S.; Khan, A.U. Nanoparticles vs. Biofilms: A Battle Against Another Paradigm of Antibiotic Resistance. MedChemComm 2016, 7, 1479–1498. [Google Scholar] [CrossRef]
- Fidelis, C.E.; de Freitas Leite, R.; Garcia, B.L.N.; Gonçalves, J.L.; Good, L.; dos Santos, M.V. Antimicrobial Activities of Polyhexamethylene Biguanide Against Biofilm-Producing Prototheca bovis Causing Bovine Mastitis. J. Dairy Sci. 2022, 106, 1383–1393. [Google Scholar] [CrossRef]
- Gilbert, P.; Moore, L.E. Cationic Antiseptics: Diversity of Action under a Common Epithet. J. Appl. Microbiol. 2005, 99, 703–715. [Google Scholar] [CrossRef]
- Chindera, K.; Mahato, M.; Kumar Sharma, A.; Horsley, H.; Kloc-Muniak, K.; Kamaruzzaman, N.F.; Kumar, S.; McFarlane, A.; Stach, J.; Bentin, T.; et al. The Antimicrobial Polymer PHMB Enters Cells and Selectively Condenses Bacterial Chromosomes. Sci. Rep. 2016, 6, 23121. [Google Scholar] [CrossRef] [PubMed]
- Leite, R.F.; Gonçalves, J.L.; Buanz, A.; Febraro, C.; Craig, D.; Van Winden, S.; Good, L.; Santos, M.V. Antimicrobial Activity of Polyhexamethylene Biguanide Nanoparticles Against Mastitis-Causing Staphylococcus aureus. JDS Commun. 2021, 2, 262–265. [Google Scholar] [CrossRef]
- Zielinska, A.; Carreiró, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Nagasamy Venkatesh, D.; Durazzo, A.; Lucarini, M.; Eder, P.; Silva, A.M.; et al. Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Molecules 2020, 25, 3731. [Google Scholar] [CrossRef]
- Leite, R.F.; Gonçalves, J.L.; Peti, A.P.F.; Figueiró, F.S.; Moraes, L.A.B.; Santos, M.V. Antimicrobial Activity of Crude Extracts from Actinomycetes Against Mastitis Pathogens. J. Dairy Sci. 2018, 101, 10116–10125. [Google Scholar] [CrossRef]
- Tomazi, T.; Ferreira, G.C.; Orsi, A.M.; Gonçalves, J.L.; Ospina, P.A.; Nydam, D.V.; Moroni, P.; dos Santos, M.V. Association of Herd-Level Risk Factors and Incidence Rate of Clinical Mastitis in 20 Brazilian Dairy Herds. Prev. Vet. Med. 2018, 161, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.; Barkema, H.W.; De Buck, J.; De Vliegher, S.; Chaffer, M.; Lewis, J.; Keefe, G.P. Identification of Bovine-Associated Coagulase-Negative Staphylococci by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Using a Direct Transfer Protocol. J. Dairy Sci. 2017, 100, 2137–2147. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). EUCAST Reading Guide for Broth Microdilution. Version 2.0, Valid from March 2020. Available online: www.eucast.org (accessed on 5 November 2024).
- Tomazi, T.; dos Santos, M.V. Antimicrobial Use for Treatment of Clinical Mastitis in Dairy Herds from Brazil and Its Association with Herd-Level Descriptors. Prev. Vet. Med. 2020, 176, 104937. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.; Gonçalves, J.L.; Leite, R.F.; Tomazi, T.; Rall, V.L.M.; Santos, M.V. Association Between Antimicrobial Use and Antimicrobial Resistance of Streptococcus uberis Causing Clinical Mastitis. J. Dairy Sci. 2021, 104, 12030–12041. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). EUCAST Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 15.0, Valid from January 2025. Available online: www.eucast.org (accessed on 5 November 2024).
- Prenafeta, A.; Sitjà, M.; Holmes, M.A.; Paterson, G.K. Short Communication: Biofilm Production Characterization of MecA and MecC Methicillin-Resistant Staphylococcus aureus Isolated from Bovine Milk in Great Britain. J. Dairy Sci. 2014, 97, 4838–4841. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćircović, I.; Ruzicka, F. Quantification of Biofilm in Microtiter Plates: Overview of Testing Conditions and Practical Recommendations for Assessment of Biofilm Production by Staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Orellano, M.S.; Isaac, P.; Breser, M.L.; Bohl, L.P.; Conesa, A.; Falcone, R.D.; Porporatto, C. Chitosan Nanoparticles Enhance the Antibacterial Activity of the Native Polymer Against Bovine Mastitis Pathogens. Carbohydr. Polym. 2019, 213, 1–9. [Google Scholar] [CrossRef]
- Breser, M.L.; Felipe, V.; Bohl, L.P.; Orellano, M.S.; Isaac, P.; Conesa, A.; Rivero, V.E.; Correa, S.G.; Bianco, I.D.; Porporatto, C. Chitosan and Cloxacillin Combination Improve Antibiotic Efficacy Against Different Lifestyle of Coagulase-Negative Staphylococcus Isolates from Chronic Bovine Mastitis. Sci. Rep. 2018, 8, 5081. [Google Scholar] [CrossRef]
- Zhang, A.; Mu, H.; Zhang, W.; Cui, G.; Zhu, J.; Duan, J. Chitosan Coupling Makes Microbial Biofilms Susceptible to Antibiotics. Sci. Rep. 2013, 3, 3364. [Google Scholar] [CrossRef]
- Enger, B.D.; Fox, L.K.; Gay, J.M.; Johnson, K.A. Reduction of Teat Skin Mastitis Pathogen Loads: Differences Between Strains, Dips, and Contact Times. J. Dairy Sci. 2015, 98, 1354–1361. [Google Scholar] [CrossRef]
- Neelam; Jain, V.K.; Singh, M.; Joshi, V.G.; Chhabra, R.; Singh, K.; Rana, Y.S. Virulence and Antimicrobial Resistance Gene Profiles of Staphylococcus aureus Associated with Clinical Mastitis in Cattle. PLoS ONE 2022, 17, e0264762. [Google Scholar] [CrossRef]
- Santos, R.P.; Souza, F.N.; Oliveira, A.C.D.; de Souza Filho, A.F.; Aizawa, J.; Moreno, L.Z.; da Cunha, A.F.; Cortez, A.; Della Libera, A.M.M.P.; Heinemann, M.B.; et al. Molecular Typing and Antimicrobial Susceptibility Profile of Staphylococcus aureus Isolates Recovered from Bovine Mastitis and Nasal Samples. Animals 2020, 10, 2143. [Google Scholar] [CrossRef] [PubMed]
- Brahma, U.; Suresh, A.; Murthy, S.; Bhandari, V.; Sharma, P. Antibiotic Resistance and Molecular Profiling of the Clinical Isolates of Staphylococcus aureus Causing Bovine Mastitis from India. Microorganisms 2022, 10, 833. [Google Scholar] [CrossRef] [PubMed]
- Khanal, S.; Boonyayatra, S.; Awaiwanont, N. Prevalence of Methicillin-Resistant Staphylococcus aureus in Dairy Farms: A Systematic Review and Meta-Analysis. Front. Vet. Sci. 2022, 9, 947154. [Google Scholar] [CrossRef] [PubMed]
- O’Dea, M.; Abraham, R.J.; Sahibzada, S.; Lee, T.; Jordan, D.; Laird, T.; Pang, S.; Buller, N.; Stegger, M.; Coombs, G.W.; et al. Antimicrobial Resistance and Genomic Insights into Bovine Mastitis-Associated Staphylococcus aureus in Australia. Vet. Microbiol. 2020, 250, 108850. [Google Scholar] [CrossRef]
- Duse, A.; Persson-waller, K.; Pedersen, K. Pathogen-Specific Risk Factors for Udder Pathogens from Clinical Mastitis in Dairy Cows. Animals 2021, 11, 2113. [Google Scholar] [CrossRef]
- Kaczorek-Łukowska, E.; Małaczewska, J.; Sowińska, P.; Szymańska, M.; Wójcik, E.A.; Siwicki, A.K. Staphylococcus aureus from Subclinical Cases of Mastitis in Dairy Cattle in Poland, What Are They Hiding? Antibiotic Resistance and Virulence Profile. Pathogens 2022, 11, 1404. [Google Scholar] [CrossRef]
- Nobrega, D.B.; Naqvi, S.A.; Dufour, S.; Deardon, R.; Kastelic, J.P.; De Buck, J.; Barkema, H.W. Critically Important Antimicrobials Are Generally Not Needed to Treat Nonsevere Clinical Mastitis in Lactating Dairy Cows: Results from a Network Meta-Analysis. J. Dairy Sci. 2020, 103, 10585–10603. [Google Scholar] [CrossRef]
- Pérez, V.K.C.; da Costa, G.M.; Guimarães, A.S.; Heinemann, M.B.; Lage, A.P.; Dorneles, E.M.S. Relationship Between Virulence Factors and Antimicrobial Resistance in Staphylococcus aureus from Bovine Mastitis. J. Glob. Antimicrob. Resist. 2020, 22, 792–802. [Google Scholar] [CrossRef]
- Wang, X.; Liu, M.; Geng, N.; Du, Y.; Li, Z.; Gao, X.; Han, B.; Liu, J.; Liu, Y. Staphylococcus aureus Mediates Pyroptosis in Bovine Mammary Epithelial Cell via Activation of NLRP3 Inflammasome. Vet. Res. 2022, 53, 10. [Google Scholar] [CrossRef]
- Yan, L.; Yang, Y.; Ma, X.; Wei, L.; Wan, X.; Zhang, Z.; Ding, J.; Peng, J.; Liu, G.; Gou, H.; et al. Effect of Two Different Drug-Resistant Staphylococcus aureus Strains on the Physiological Properties of MAC-T Cells and Their Transcriptome Analysis. Front. Vet. Sci. 2022, 9, 818928. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P.; Niedziela, D.A.; Leonard, F.C.; Keane, O.M. The in Vitro Host Cell Immune Response to Bovine-Adapted Staphylococcus aureus Varies According to Bacterial Lineage. Sci. Rep. 2019, 9, 6134. [Google Scholar] [CrossRef] [PubMed]
- Mello, P.L.; Riboli, D.F.M.; de Almeida Martins, L.; Brito, M.A.V.P.; Victória, C.; Romero, L.C.; da Cunha, M.d.L.R.d.S. Staphylococcus spp. Isolated from Bovine Subclinical Mastitis in Different Regions of Brazil: Molecular Typing and Biofilm Gene Expression Analysis by Rt-Qpcr. Antibiotics 2020, 9, 888. [Google Scholar] [CrossRef]
- Thiran, E.; Di Ciccio, P.A.; Graber, H.U.; Zanardi, E.; Ianieri, A.; Hummerjohann, J. Biofilm Formation of Staphylococcus aureus Dairy Isolates Representing Different Genotypes. J. Dairy Sci. 2018, 101, 1000–1012. [Google Scholar] [CrossRef]
- Ren, Q.; Liao, G.; Wu, Z.; Lv, J.; Chen, W. Prevalence and Characterization of Staphylococcus aureus Isolates from Subclinical Bovine Mastitis in Southern Xinjiang, China. J. Dairy Sci. 2020, 103, 3368–3380. [Google Scholar] [CrossRef]
- da Costa Krewer, C.; Santos Amanso, E.; Veneroni Gouveia, G.; de Lima Souza, R.; da Costa, M.M.; Aparecido Mota, R. Resistance to Antimicrobials and Biofilm Formation in Staphylococcus spp. Isolated from Bovine Mastitis in the Northeast of Brazil. Trop. Anim. Health Prod. 2015, 47, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Marques, V.F.; da Motta, C.C.; Soares, B.d.S.; de Melo, D.A.; Coelho, S.d.M.d.O.; Coelho, I.d.S.; Barbosa, H.S.; de Souza, M.M.S. Biofilm Production and Beta-Lactamic Resistance in Brazilian Staphylococcus aureus Isolates from Bovine Mastitis. Braz. J. Microbiol. 2017, 48, 118–124. [Google Scholar] [CrossRef]
- Trubenová, B.; Roizman, D.; Moter, A.; Rolff, J.; Regoes, R.R. Population Genetics, Biofilm Recalcitrance, and Antibiotic Resistance Evolution. Trends Microbiol. 2022, 30, 841–852. [Google Scholar] [CrossRef]
- Balaure, P.C.; Grumezescu, A.M. Recent Advances in Surface Nanoengineering for Biofilm Prevention and Control. Part II: Active, Combined Active and Passive, and Smart Bacteria-Responsive Antibiofilm Nanocoatings. Nanomaterials 2020, 10, 1527. [Google Scholar] [CrossRef]
- Puertas-Bartolomé, M.; Mora-Boza, A.; García-Fernández, L. Emerging Biofabrication Techniques: A Review on Natural Polymers for Biomedical Applications. Polymers 2021, 13, 1209. [Google Scholar] [CrossRef]
- Orellano, M.S.; Bohl, L.P.; Breser, M.L.; Isaac, P.; Falcone, R.D.; Porporatto, C. A Comparative Study of Antimicrobial Activity of Differently-Synthesized Chitosan Nanoparticles Against Bovine Mastitis Pathogens. Soft Matter 2021, 17, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Acosta, A.C.; Santos, A.S.; Silva, F.A.G.; Medeiros, E.S.; de Oliveira, H.P.; Costa, M.M.; Fernandes, A.W.C.; Pinheiro Júnior, J.W.; Mota, R.A. Atividade Antibacteriana de Nanopartículas de Polipirrol Diante de Staphylococcus aureus Isolados de Amostras de Leite de Vacas e Cabras Com Mastite. Arq. Bras. Med. Vet. Zootec. 2020, 72, 1047–1050. [Google Scholar] [CrossRef]
- Manan, A.; Aqib, A.I.; Shahbaz, A.; Khan, S.R.; Akram, K.; Majeed, H.; Muneer, A.; Murtaza, M.; Afrasiab, M.; Merola, C.; et al. Modification of the Drug Resistance of Emerging Milk-Borne Pathogens Through Sodium Alginate-Based Antibiotics and Nanoparticles. Front. Vet. Sci. 2023, 10, 1130130. [Google Scholar] [CrossRef]
- Niculescu, A.G.; Grumezescu, A.M. Applications of Chitosan-Alginate-Based Nanoparticles—An Up-to-Date Review. Nanomaterials 2022, 12, 186. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.F. Fighting Back Against Antimicrobial Resistance with Comprehensive Policy and Education: A Narrative Review. Antibiotics 2022, 11, 644. [Google Scholar] [CrossRef]
- Feng, P.; Luo, Y.; Ke, C.; Qiu, H.; Wang, W.; Zhu, Y.; Hou, R.; Xu, L.; Wu, S. Chitosan-Based Functional Materials for Skin Wound Repair: Mechanisms and Applications. Front. Bioeng. Biotechnol. 2021, 9, 650598. [Google Scholar] [CrossRef]
- Huang, Y.W.; Cambre, M.; Lee, H.J. The Toxicity of Nanoparticles Depends on Multiple Molecular and Physicochemical Mechanisms. Int. J. Mol. Sci. 2017, 18, 2702. [Google Scholar] [CrossRef]
- Rao, H.; Choo, S.; Mahalingam, S.R.R.; Adisuri, D.S.; Madhavan, P.; Akim, A.M.; Chong, P.P. Approaches for Mitigating Microbial Biofilm-Related Drug Resistance: A Focus on Micro- and Nanotechnologies. Molecules 2021, 26, 1870. [Google Scholar] [CrossRef]
- Mohanta, Y.K.; Chakrabartty, I.; Mishra, A.K.; Chopra, H.; Mahanta, S.; Avula, S.K.; Patowary, K.; Ahmed, R.; Mishra, B.; Mohanta, T.K.; et al. Nanotechnology in Combating Biofilm: A Smart and Promising Therapeutic Strategy. Front. Microbiol. 2023, 13, 1028086. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Mohammadi, M.C.; Anzabi, Y.; Shayegh, J. Comparison of the Frequency of Biofilm-Forming Genes (IcaABCD) in Methicillin-Resistant S. aureus Strains Isolated from Human and Livestock. Arch. Razi Inst. 2021, 76, 1655–1663. [Google Scholar] [CrossRef]
- Aguayo, P.R.; Larenas, T.B.; Godoy, C.A.; Rivas, B.C.; González-Casanova, J.; Rojas-Gómez, D.; Fuentes, N.C. Antimicrobial and Antibiofilm Capacity of Chitosan Nanoparticles Against Wild Type Strain of Pseudomonas sp. Isolated from Milk of Cows Diagnosed with Bovine Mastitis. Antibiotics 2020, 9, 551. [Google Scholar] [CrossRef] [PubMed]
- Cacciatore, F.A.; Brandelli, A.; Malheiros, P. da S. Combining Natural Antimicrobials and Nanotechnology for Disinfecting Food Surfaces and Control Microbial Biofilm Formation. Crit. Rev. Food Sci. Nutr. 2021, 61, 3771–3782. [Google Scholar] [CrossRef] [PubMed]
- Saeki, E.K.; Yamada, A.Y.; de Araujo, L.A.; Anversa, L.; de Oliveira Garcia, D.; de Souza, R.L.B.; Martins, H.M.; Kobayashi, R.K.T.; Nakazato, G. Subinhibitory Concentrations of Biogenic Silver Nanoparticles Affect Motility and Biofilm Formation in Pseudomonas aeruginosa. Front. Cell Infect. Microbiol. 2021, 11, 656984. [Google Scholar] [CrossRef]
- Swolana, D.; Kępa, M.; Kruszniewska-Rajs, C.; Wojtyczka, R.D. Antibiofilm Effect of Silver Nanoparticles in Changing the Biofilm-Related Gene Expression of Staphylococcus epidermidis. Int. J. Mol. Sci. 2022, 23, 9257. [Google Scholar] [CrossRef]
- Ouyang, K.; Mortimer, M.; Holden, P.A.; Cai, P.; Wu, Y.; Gao, C.; Huang, Q. Towards a Better Understanding of Pseudomonas putida Biofilm Formation in the Presence of ZnO Nanoparticles (NPs): Role of NP Concentration. Environ. Int. 2020, 137, 105485. [Google Scholar] [CrossRef]
- Park, K.H.; Jung, M.; Kim, D.Y.; Lee, Y.M.; Lee, M.S.; Ryu, B.H.; Hong, S.I.; Hong, K.W.; Bae, I.G.; Cho, O.H. Effects of Subinhibitory Concentrations of Chlorhexidine and Mupirocin on Biofilm Formation in Clinical Meticillin-Resistant Staphylococcus aureus. J. Hosp. Infect. 2020, 106, 295–302. [Google Scholar] [CrossRef]
- Ebrahimi, A.; Hemati, M.; Dehkordi, S.H.; Bahadoran, S.; Khoshnood, S.; Khubani, S.; Faraj, M.D.; Alni, R.H. Chlorhexidine Digluconate Effects on Planktonic Growth and Biofilm Formation in Some Field Isolates of Animal Bacterial Pathogens. Jundishapur J. Nat. Pharm. Prod. 2014, 9, e14298. [Google Scholar] [CrossRef]
- Bottery, M.J.; Pitchford, J.W.; Friman, V.P. Ecology and Evolution of Antimicrobial Resistance in Bacterial Communities. ISME J. 2021, 15, 939–948. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Alonso-Calleja, C.; García-Fernández, C.; Carballo, J.; Capita, R. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) for Twelve Antimicrobials (Biocides and Antibiotics) in Eight Strains of Listeria Monocytogenes. Biology 2021, 11, 46. [Google Scholar] [CrossRef]
- Nguyen, P.T.M.; Nguyen, M.T.H.; Bolhuis, A. Inhibition of Biofilm Formation by Alpha-Mangostin Loaded Nanoparticles against Staphylococcus aureus. Saudi J. Biol. Sci. 2021, 28, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lin, Q.; Sheng, M.; Ding, T.; Li, B.; Gao, Y.; Tan, Y. Antibiofilm Effect of Cinnamaldehyde-Chitosan Nanoparticles Against the Biofilm of Staphylococcus aureus. Antibiotics 2022, 11, 1403. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Ma, S.; Leonhard, M.; Moser, D.; Haselmann, G.M.; Wang, J.; Eder, D.; Schneider-Stickler, B. Enhancing Antibiofilm Activity with Functional Chitosan Nanoparticles Targeting Biofilm Cells and Biofilm Matrix. Carbohydr. Polym. 2018, 200, 35–42. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, L.; Su, L.; Van der Mei, H.C.; Jutte, P.C.; Ren, Y.; Busscher, H.J. Nanotechnology-Based Antimicrobials and Delivery Systems for Biofilm-Infection Control. Chem. Soc. Rev. 2019, 48, 428–446. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering Precision Nanoparticles for Drug Delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.L.; Oliver, S.P.; Fydenkevez, M.E. Evaluation of Experimental Teat Dip Containing Sodium Chlorite and Lactic Acid by Excised Teat Assay. J. Dairy Sci. 1984, 67, 3075–3080. [Google Scholar] [CrossRef]
- West, A.M.; Teska, P.J.; Lineback, C.B.; Oliver, H.F. Strain, Disinfectant, Concentration, and Contact Time Quantitatively Impact Disinfectant Efficacy. Antimicrob. Resist. Infect. Control 2018, 7, 49. [Google Scholar] [CrossRef]
- Herdiana, Y.; Wathoni, N.; Shamsuddin, S.; Muchtaridi, M. Drug Release Study of the Chitosan-Based Nanoparticles. Heliyon 2022, 8, e08674. [Google Scholar] [CrossRef] [PubMed]
- Schwenker, J.A.; Schotte, U.; Hölzel, C.S. Minimum Inhibitory Concentrations of Chlorhexidine- and Lactic Acid-Based Teat Disinfectants: An Intervention Trial Assessing Bacterial Selection and Susceptibility. J. Dairy Sci. 2022, 105, 734–747. [Google Scholar] [CrossRef]
- Rozman, U.; Pušnik, M.; Kmetec, S.; Duh, D.; Turk, S.Š. Reduced Susceptibility and Increased Resistance of Bacteria Against Disinfectants: A Systematic Review. Microorganisms 2021, 9, 2550. [Google Scholar] [CrossRef]
- Klostermann, K.; Crispie, F.; Flynn, J.; Meaney, W.J.; Paul Ross, R.; Hill, C. Efficacy of a Teat Dip Containing the Bacteriocin Lacticin 3147 to Eliminate Gram-Positive Pathogens Associated with Bovine Mastitis. J. Dairy Res. 2010, 77, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, S.R.; Garvey, M.; Flynn, J.; O’Brien, B.; Gleeson, D. The Effect of Disinfectant Ingredients on Teat Skin Bacteria Associated with Mastitis in Irish Dairy Herds. Ir. Vet. J. 2021, 74, 1. [Google Scholar] [CrossRef] [PubMed]
- Oura, L.Y.; Fox, L.K.; Warf, C.C.; Kemp, G.K. Efficacy of Two Acidified Chlorite Postmilking Teat Disinfectants with Sodium Dodecylbenzene Sulfonic Acid on Prevention of Contagious Mastitis Using an Experimental Challenge Protocol. J. Dairy Sci. 2002, 85, 252–257. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Antimicrobial | Tested Isolate | Resistance (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1650 | 1651 | 1658 | 1663 | 1666 | 1667 | 1677 | 1680 | 1684 | 1688 | ||
| Amoxicillin | 0.25 | 1 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.06 | 90 |
| Ampicillin | 0.25 | 0.5 | 0.12 | 0.12 | 0.12 | 0.25 | 0.25 | 0.12 | 0.12 | 0.06 | 40 |
| Cefalexin | 0.5 | 1 | 2 | 2 | 2 | 8 | 2 | 1 | 2 | 0.5 | 10 |
| Cefoxitin | 2 | 2 | 2 | 2 | 2 | 8 | 2 | 2 | 2 | 2 | 10 |
| Ceftiofur | 1 | 2 | 2 | 2 | 2 | 8 | 2 | 4 | 2 | 1 | 20 |
| Ciprofloxacin | 0.06 | 0.12 | 0.25 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.06 | 0 |
| Enrofloxacin | 0.12 | 0.06 | 0.12 | 0.12 | 0.06 | 0.12 | 0.06 | 0.12 | 0.06 | 0.06 | 0 |
| Erythromycin | 0.12 | 0.12 | 4 | 1 | 1 | 1 | 0.5 | 1 | 1 | 1 | 10 |
| Gentamicin | 0.25 | 0.25 | 0.12 | 1 | 0.5 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0 |
| Lincomycin | 0.12 | 0.25 | 0.25 | 0.12 | 0.25 | 0.12 | 0.12 | 0.25 | 0.12 | 0.12 | 0 |
| Oxytetracycline | 1 | 8 | 8 | 8 | 8 | 4 | 4 | 4 | 8 | 4 | 90 |
| Penicillin G | 0.25 | 0.25 | 0.5 | 0.25 | 0.03 | 0.5 | 0.25 | 0.25 | 0.25 | 0.03 | 80 |
| Tetracycline | 1 | 1 | 1 | 4 | 1 | 4 | 1 | 1 | 64 | 1 | 30 |
| Tested Isolate | PHMB NPs 1 | PHMB 2 | ||
|---|---|---|---|---|
| MIC 3 | MBC 4 | MIC 3 | MBC 4 | |
| 1650 | 0.25 | 2 | 0.5 | 2 |
| 1651 | 0.25 | 1 | 0.5 | 8 |
| 1658 | 0.5 | 4 | 0.5 | 8 |
| 1663 | 0.25 | 4 | 0.5 | 4 |
| 1666 | 0.25 | 2 | 1 | 8 |
| 1667 | 0.25 | 4 | 0.5 | 8 |
| 1677 | 0.25 | 1 | 1 | 8 |
| 1680 | 0.5 | 4 | 0.5 | 8 |
| 1684 | 1 | 8 | 0.5 | 8 |
| 1688 | 0.5 | 4 | 0.5 | 8 |
| ATCC 29213 5 | 0.5 | 4 | 0.5 | 8 |
| Concentration | Preformed Biofilm | Biofilm Inhibition | ||
|---|---|---|---|---|
| Treatments | ||||
| PHMB NPs | PHMB | PHMB NPs | PHMB | |
| 0 | 100 1 | 100 | 100 | 100 |
| 0.12 | 105.3 | 114.5 | 82.9 | 108.6 |
| 0.25 | 101 | 106.2 | 64.9 | 105.5 |
| 0.5 | 97 | 100.9 | 53.6 | 106.9 |
| 1 | 96.3 | 91.2 | 46.8 | 102 |
| 2 | 89.6 | 83.4 | 43.9 | 84.2 |
| 4 | 84.4 | 73.9 | 43.5 | 56.7 |
| Tested Isolate | Control (Log) 1 | PHMB NPs 1 | PHMB 1 | CHG 1 | NaDCC 1 | PVP-I 1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Log 2 | % Log Reduction 3 | Log 2 | % Log Reduction 3 | Log 2 | % Log Reduction 3 | Log 2 | % Log Reduction 3 | Log 2 | % Log Reduction 3 | ||
| 1650 | 2.60 A | 1.76 | 32.27 AB | 0.15 | 94.21 B | 0.23 | 91.32 B | 0.56 | 78.32 AB | 0 | 100 B |
| 1651 | 1.33 A | 1.30 | 1.98 AB | 0.42 | 68.32 AB | 0 | 100 B | 0 | 100 B | 0.19 | 85.34 AB |
| 1658 | 2.79 A | 1.30 | 53.37 AB | 0 | 100 B | 0.24 | 91.45 B | 0 | 100 B | 0.12 | 95.72 B |
| 1663 | 2.58 A | 1.76 | 31.49 AB | 0.33 | 87.37 AB | 0 | 100 B | 0 | 100 B | 0.19 | 92.45 AB |
| 1666 | 2.64 A | 1.54 | 41.77 AB | 0.46 | 82.42 B | 0.31 | 88.12 B | 0.71 | 73.06 AB | 0.33 | 87.68 B |
| 1667 | 2.88 A | 1.81 | 37.05 AB | 0 | 100 B | 0 | 100 B | 0.37 | 87.19 B | 0 | 100 B |
| 1677 | 2.90 A | 1.62 | 44.10 AB | 0 | 100 B | 0.08 | 97.40 B | 0 | 100 B | 0.08 | 97.40 B |
| 1680 | 3.45 A | 1.77 | 48.70 AB | 0 | 100 B | 0 | 100 B | 0.12 | 96.55 B | 0.08 | 97.82 B |
| 1684 | 2.76 A | 1.39 | 49.58 AB | 0.69 | 74.84 AB | 0.67 | 75.72 AB | 0.42 | 84.78 AB | 0.08 | 97.27 B |
| 1688 | 2.47 A | 1.60 | 35.35 AB | 0 | 100 B | 0.08 | 96.96 B | 0.19 | 92.13 B | 0 | 100 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite, R.d.F.; Garcia, B.L.N.; Barbosa, K.d.S.; Mitsunaga, T.M.; Fidelis, C.E.; Dias, B.J.M.; Miranda, R.R.d.; Zucolotto, V.; Good, L.; Santos, M.V.d. Polyhexamethylene Biguanide Nanoparticles Inhibit Biofilm Formation by Mastitis-Causing Staphylococcus aureus. Vet. Sci. 2025, 12, 507. https://doi.org/10.3390/vetsci12050507
Leite RdF, Garcia BLN, Barbosa KdS, Mitsunaga TM, Fidelis CE, Dias BJM, Miranda RRd, Zucolotto V, Good L, Santos MVd. Polyhexamethylene Biguanide Nanoparticles Inhibit Biofilm Formation by Mastitis-Causing Staphylococcus aureus. Veterinary Sciences. 2025; 12(5):507. https://doi.org/10.3390/vetsci12050507
Chicago/Turabian StyleLeite, Renata de Freitas, Breno Luis Nery Garcia, Kristian da Silva Barbosa, Thatiane Mendes Mitsunaga, Carlos Eduardo Fidelis, Bruna Juliana Moreira Dias, Renata Rank de Miranda, Valtencir Zucolotto, Liam Good, and Marcos Veiga dos Santos. 2025. "Polyhexamethylene Biguanide Nanoparticles Inhibit Biofilm Formation by Mastitis-Causing Staphylococcus aureus" Veterinary Sciences 12, no. 5: 507. https://doi.org/10.3390/vetsci12050507
APA StyleLeite, R. d. F., Garcia, B. L. N., Barbosa, K. d. S., Mitsunaga, T. M., Fidelis, C. E., Dias, B. J. M., Miranda, R. R. d., Zucolotto, V., Good, L., & Santos, M. V. d. (2025). Polyhexamethylene Biguanide Nanoparticles Inhibit Biofilm Formation by Mastitis-Causing Staphylococcus aureus. Veterinary Sciences, 12(5), 507. https://doi.org/10.3390/vetsci12050507