

Lactobacillus Re-Engineers Gut Microbiota to Overcome E. coli Colonization Resistance in Mice

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Models and Experimental Design
2.3. Sample Collection
2.4. Sample Measurement
2.5. Statistical Analysis
3. Results
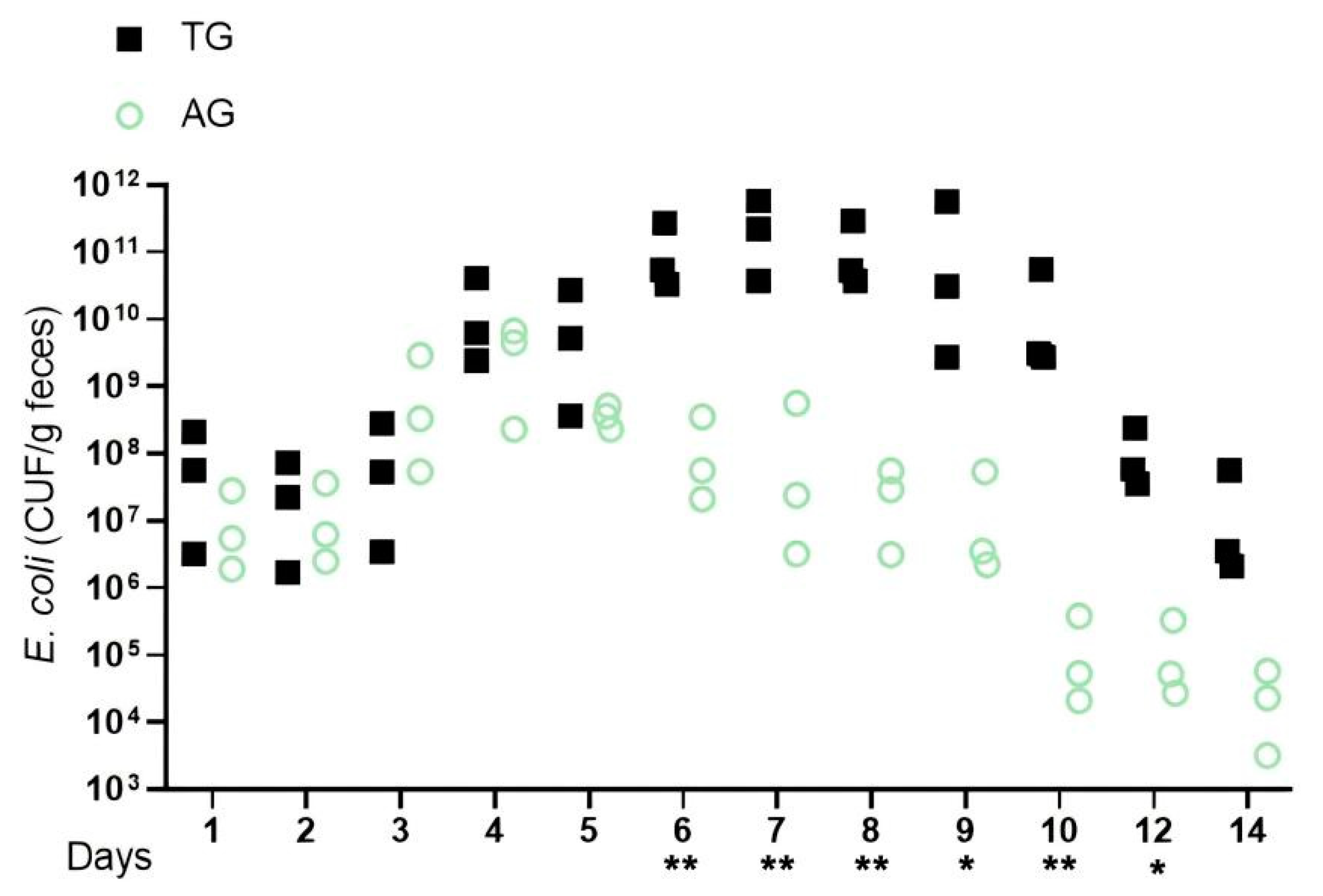
3.1. E. coli O78 Strain Overcomes Microbiota-Mediated Growth Inhibition
3.2. A Change in Mouse Weight Is Linked to Lactobacillus Broken E. coli Colonization Resistance
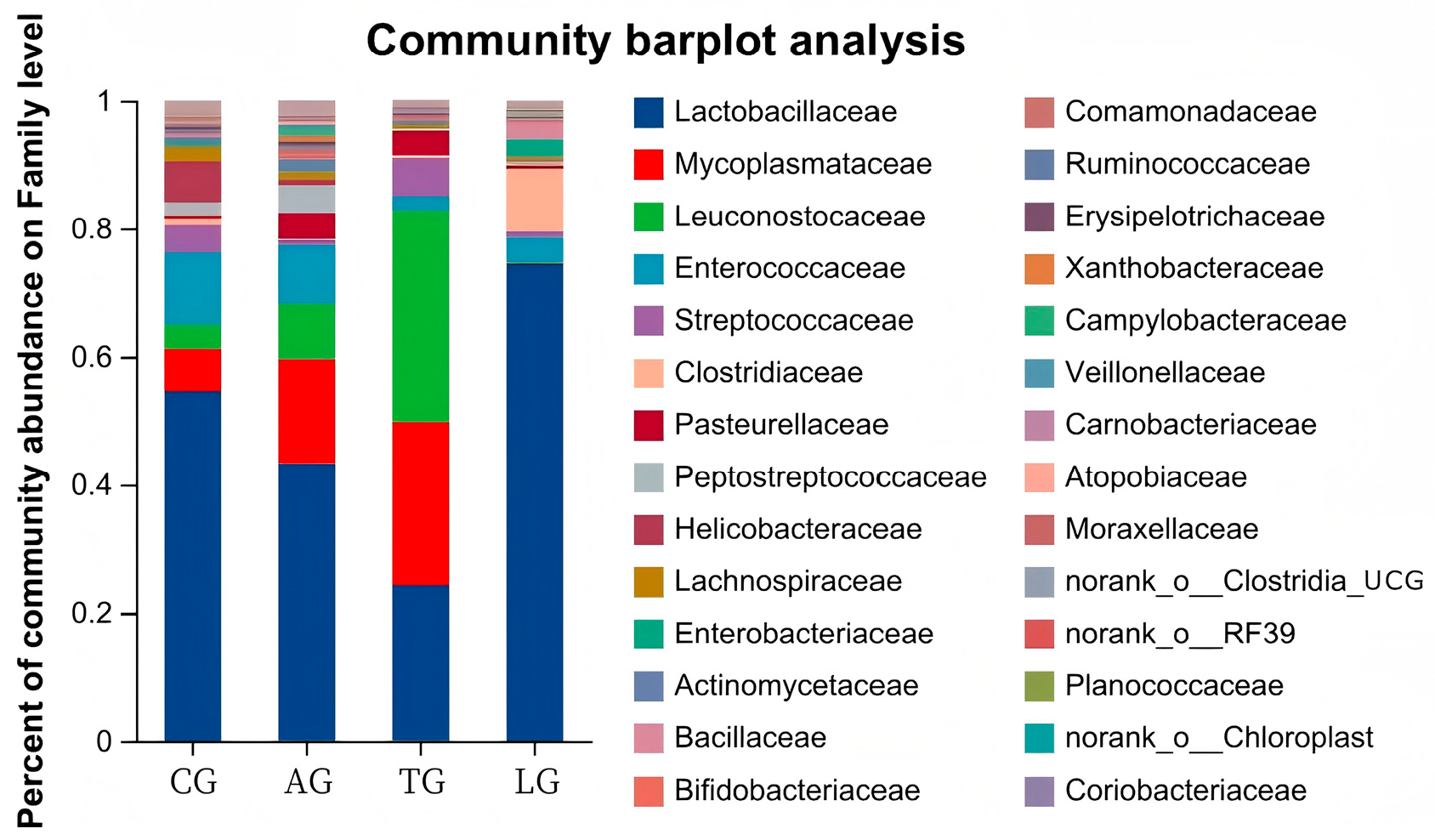
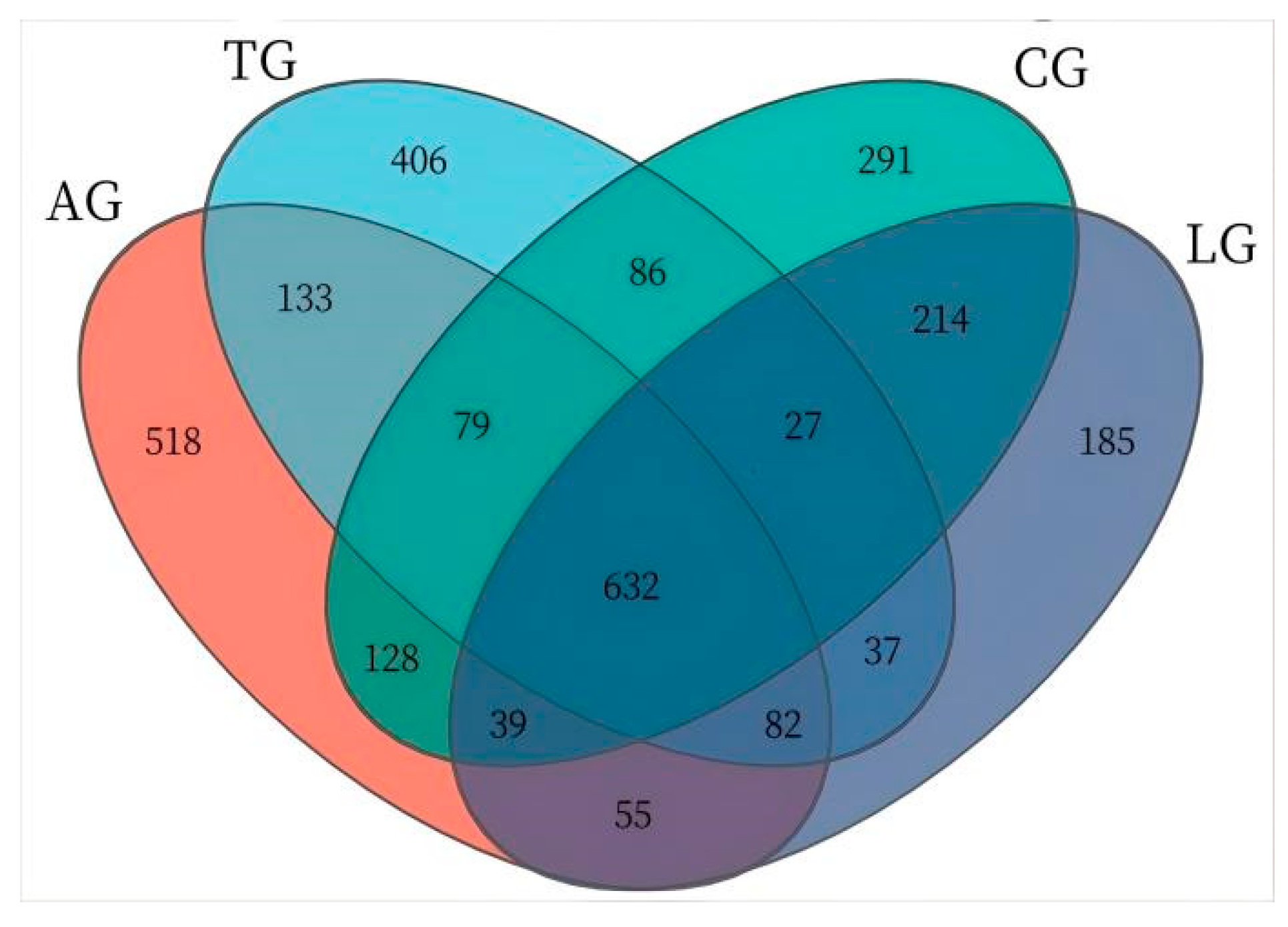
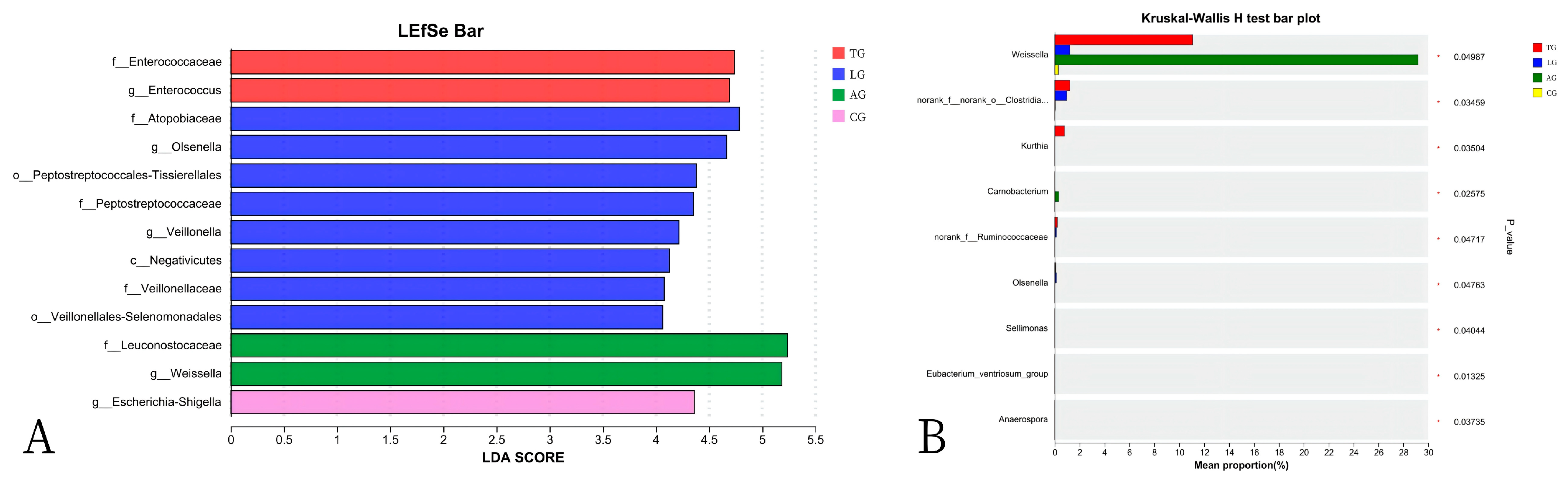
3.3. E. coli Infestation and Lactobacillus Intervention Are Linked to Compositional Gut Microbiota Changes
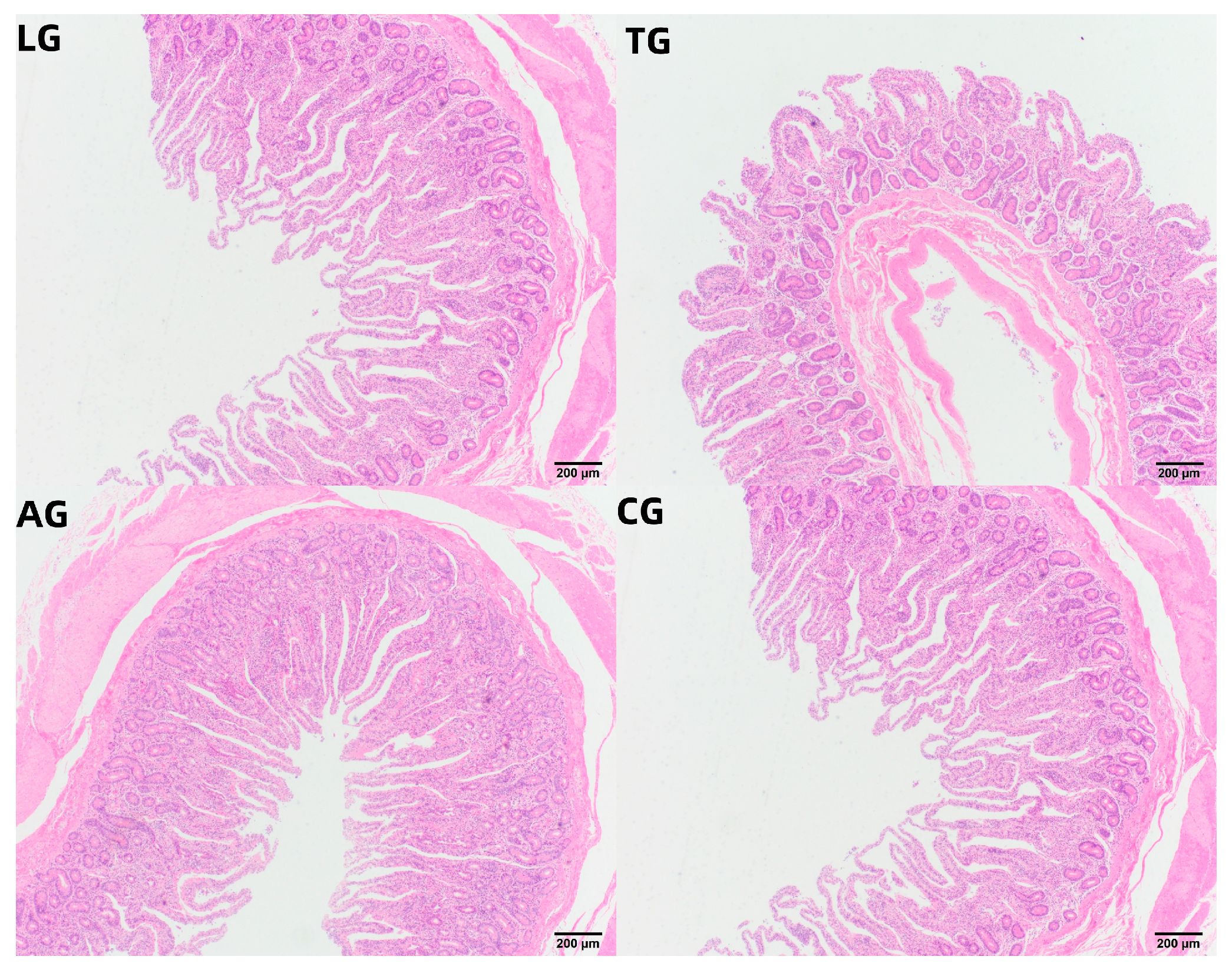
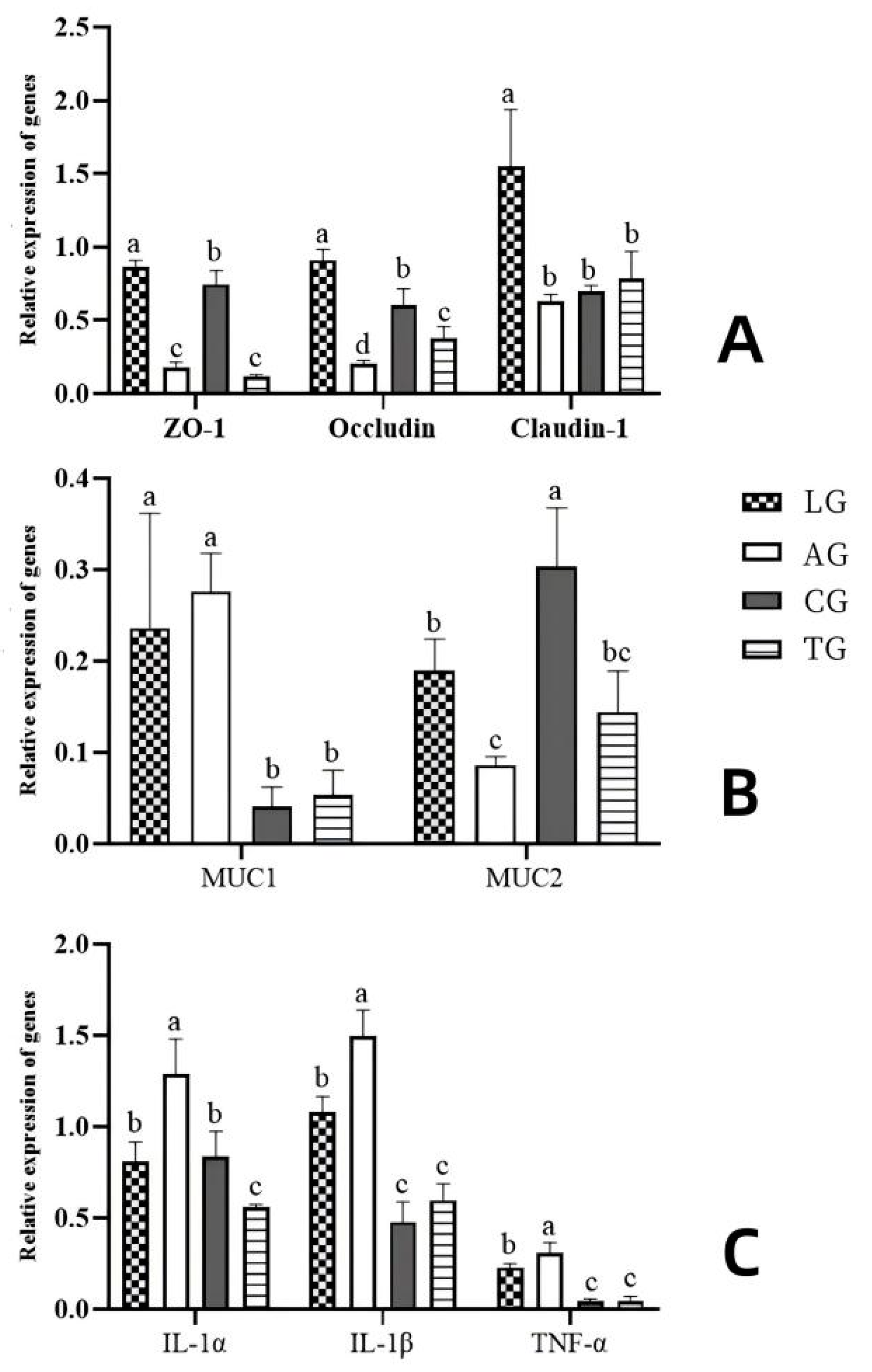
3.4. Jejunal Inflammation Elicited During E. coli Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. A Health Perspective on the Role of the Environment in One Health; No. WHO/EURO: 2022-5290-45054-64214; World Health Organization. Regional Office for Europe: Geneva, Switzerland, 2022. [Google Scholar]
- Khan, I.M.; Nassar, N.; Chang, H.; Khan, S.; Cheng, M.; Wang, Z.; Xiang, X. The microbiota: A key regulator of health, productivity, and reproductive success in mammals. Front. Microbiol. 2024, 15, 1480811. [Google Scholar] [CrossRef] [PubMed]
- Mosa, M.I.; Salem, H.M.; Bastamy, M.A.; Amer, M.M. Pathogenic and non-pathogenic factors; especially infectious bursal disease viruses; affect chicken digestive system microbiota and methods of its evaluation and recovery: A review. Egypt. J. Vet. Sci. 2023, 54, 733–760. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, X.R.; Huang, J.Q.; Li, B. Pathogenic Escherichia coli. In Molecular Medical Microbiology, 3rd ed.; Academic Press: Cambridge, MA, USA, 2024; Chapter 53; pp. 1065–1096. [Google Scholar] [CrossRef]
- Wang, D.; Zheng, Y.; Fan, Y.; He, Y.; Liu, K.; Deng, S.; Liu, Y. Sodium humate-derived gut microbiota ameliorates intestinal dysfunction induced by Salmonella Typhimurium in mice. Microbiol. Spectr. 2023, 11, e05348-22. [Google Scholar] [CrossRef] [PubMed]
- Ormsby, M.J.; White, H.L.; Metcalf, R.; Oliver, D.M.; Quilliam, R.S. Clinically important E. coli strains can persist, and retain their pathogenicity, on environmental plastic and fabric waste. Environ. Pollut. 2023, 326, 121466. [Google Scholar] [CrossRef]
- Koosha, R.Z.; Fazel, P.; Sedighian, H.; Behzadi, E.; Ch, M.H.; Fooladi, A.A.I. The impact of the gut microbiome on toxigenic bacteria. Microb. Pathog. 2021, 160, 105188. [Google Scholar] [CrossRef]
- Gieryńska, M.; Szulc-Dąbrowska, L.; Struzik, J.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P. Integrity of the intestinal barrier: The involvement of epithelial cells and microbiota—A mutual relationship. Animals 2022, 12, 145. [Google Scholar] [CrossRef]
- Endale, H.; Mathewos, M.; Abdeta, D. Potential causes of spread of antimicrobial resistance and preventive measures in one health perspective-a review. Infect. Drug Resist. 2023, 16, 7515–7545. [Google Scholar] [CrossRef]
- Zhang, T.; Nickerson, R.; Zhang, W.; Peng, X.; Shang, Y.; Zhou, Y.; Luo, Q.; Wen, G.; Cheng, Z. The impacts of animal agriculture on One Health—Bacterial zoonosis, antimicrobial resistance, and beyond. One Health 2024, 18, 100748. [Google Scholar] [CrossRef]
- Haque, H.; Sarker, S.; Islam, S.; Islam, A.; Karim, R.; Kayesh, M.E.H.; Shiddiky, M.J.A.; Anwer, M.S. Sustainable antibiotic-free broiler meat production: Current trends, challenges, and possibilities in a developing country perspective. Biology 2020, 9, 411. [Google Scholar] [CrossRef]
- Serwecińska, L. Antimicrobials and antibiotic-resistant bacteria: A risk to the environment and to public health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- Arnold, B.J.; Huang, I.T.; Hanage, W.P. Horizontal gene transfer and adaptive evolution in bacteria. Nat. Rev. Microbiol. 2022, 20, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Gemeda, B.A.; Wieland, B.; Alemayehu, G.; Knight-Jones, T.J.D.; Wodajo, H.D.; Tefera, M.; Kumbe, A.; Olani, A.; Abera, S.; Amenu, K. Antimicrobial resistance of Escherichia coli isolates from Livestock and the environment in extensive smallholder Livestock production systems in Ethiopia. Antibiotics 2023, 12, 941. [Google Scholar] [CrossRef] [PubMed]
- Kakakhel, M.A.; Bibi, N.; Mahboub, H.H.; Wu, F.; Sajjad, W.; Din, S.Z.U.; Hefny, A.A.; Wang, W. Influence of biosynthesized nanoparticles exposure on mortality, residual deposition, and intestinal bacterial dysbiosis in Cyprinus carpio. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 263, 109473. [Google Scholar] [CrossRef] [PubMed]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: A door to the body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Liu, S.; Cheng, L.; Liu, Y.; Zhan, S.; Wu, Z.; Zhang, X. Relationship between dietary polyphenols and gut microbiota: New clues to improve cognitive disorders, mood disorders and circadian rhythms. Foods 2023, 12, 1309. [Google Scholar] [CrossRef]
- Shi, Y.; Luo, J.; Narbad, A.; Chen, Q. Advances in lactobacillus restoration for β-Lactam antibiotic-Induced dysbiosis: A system review in intestinal microbiota and immune homeostasis. Microorganisms 2023, 11, 179. [Google Scholar] [CrossRef]
- Zou, T.; Xie, F.; Liang, P.; Chen, J.; Wang, Z.; Du, M.; You, J. Polysaccharide-rich fractions from Enteromorpha prolifera improve hepatic steatosis and gut barrier integrity in high-fat diet-induced obese mice linking to modulation of gut microbiota. Biomed. Pharmacother. 2023, 157, 114034. [Google Scholar] [CrossRef]
- Chen, D.-W.; Chen, C.-M.; Qu, H.-X.; Ren, C.-Y.; Yan, X.-T.; Huang, Y.-J.; Guan, C.-R.; Zhang, C.-C.; Li, Q.-M.; Gu, R.-X. Screening of Lactobacillus strains that enhance SCFA uptake in intestinal epithelial cells. Eur. Food Res. Technol. 2021, 247, 1049–1060. [Google Scholar] [CrossRef]
- Kaur, H.; Ali, S.A. Probiotics and gut microbiota: Mechanistic insights into gut immune homeostasis through TLR pathway regulation. Food Funct. 2022, 13, 7423–7447. [Google Scholar] [CrossRef]
- Rogers, A.W.; Radlinski, L.C.; Nguyen, H.; Tiffany, C.R.; Carvalho, T.P.; Masson, H.L.; Goodson, M.L.; Bechtold, L.; Yamazaki, K.; Liou, M.J.; et al. Salmonella re-engineers the intestinal environment to break colonization resistance in the presence of a compositionally intact microbiota. Cell Host Microbe 2024, 32, 1774–1786. [Google Scholar] [CrossRef]
- Barathan, M.; Ng, S.L.; Lokanathan, Y.; Ng, M.H.; Law, J.X. The Profound Influence of Gut Microbiome and Extracellular Vesicles on Animal Health and Disease. Int. J. Mol. Sci. 2024, 25, 4024. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, D.; Saadah, O.; Mosli, M.; Edris, S.; Alhindi, R.; Bahieldin, A. Dysbiosis of gut microbiota in inflammatory bowel disease: Current therapies and potential for microbiota-modulating therapeutic approaches. Bosn. J. Basic Med. Sci. 2021, 21, 270. [Google Scholar] [CrossRef] [PubMed]
- Flandroy, L.; Poutahidis, T.; Berg, G.; Clarke, G.; Dao, M.-C.; Decaestecker, E.; Furman, E.; Haahtela, T.; Massart, S.; Plovier, H.; et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci. Total Environ. 2018, 627, 1018–1038. [Google Scholar] [CrossRef] [PubMed]
- Kakakhel, M.A.; Jamil, A.; Narwal, N. Effects of Silver Nanoparticles on Ctenopharyngodon idella: Synthesis, Characterization, Antibacterial Activity, and Toxicological Assessment. Microsc. Res. Tech. 2024, 88, 686–695. [Google Scholar] [CrossRef]
- Liu, N.; Feng, G.; Zhang, X.; Hu, Q.; Sun, S.; Sun, J.; Sun, Y.; Wang, R.; Zhang, Y.; Wang, P.; et al. The functional role of lactoferrin in intestine mucosal immune system and inflammatory bowel disease. Front. Nutr. 2021, 8, 759507. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Ni, J.; Zhang, C.; Jia, J.; Wu, G.; Sun, H.; Wang, S. Berberine relieves metabolic syndrome in mice by inhibiting liver inflammation caused by a high-fat diet and potential association with gut microbiota. Front. Microbiol. 2022, 12, 752512. [Google Scholar] [CrossRef]
- Sharma, C.; Rokana, N.; Chandra, M.; Singh, B.P.; Gulhane, R.D.; Gill, J.P.S.; Ray, P.; Puniya, A.K.; Panwar, H. Antimicrobial resistance: Its surveillance, impact, and alternative management strategies in dairy animals. Front. Vet. Sci. 2018, 4, 237. [Google Scholar] [CrossRef]
- Hernández-González, J.C.; Martínez-Tapia, A.; Lazcano-Hernández, G.; García-Pérez, B.E.; Castrejón-Jiménez, N.S. Bacteriocins from lactic acid bacteria. A powerful alternative as antimicrobials, probiotics, and immunomodulators in veterinary medicine. Animals 2021, 11, 979. [Google Scholar] [CrossRef]
- Feng, W.; Liu, J.; Cheng, H.; Zhang, D.; Tan, Y.; Peng, C. Dietary compounds in modulation of gut microbiota-derived metabolites. Front. Nutr. 2022, 9, 939571. [Google Scholar] [CrossRef]
- Barbara, G.; Barbaro, M.R.; Fuschi, D.; Palombo, M.; Falangone, F.; Cremon, C.; Marasco, G.; Stanghellini, V. Inflammatory and microbiota-related regulation of the intestinal epithelial barrier. Front. Nutr. 2021, 8, 718356. [Google Scholar] [CrossRef]
- Anee, I.J.; Alam, S.; Begum, R.A.; Shahjahan, R.M.; Khandaker, A.M. The role of probiotics on animal health and nutrition. J. Basic Appl. Zool. 2021, 82, 52. [Google Scholar] [CrossRef]
- Dong, H.; Liu, B.; Li, A.; Iqbal, M.; Mehmood, K.; Jamil, T.; Chang, Y.-F.; Zhang, H.; Wu, Q. Microbiome analysis reveals the attenuation effect of lactobacillus from yaks on diarrhea via modulation of gut microbiota. Front. Cell. Infect. Microbiol. 2021, 10, 610781. [Google Scholar] [CrossRef] [PubMed]
- Pogačar, M.Š.; Langerholc, T.; Mičetić-Turk, D.; Možina, S.S.; Klančnik, A. Effect of Lactobacillus spp. on adhesion, invasion, and translocation of Campylobacter jejuni in chicken and pig small-intestinal epithelial cell lines. BMC Vet. Res. 2020, 16, 34. [Google Scholar] [CrossRef]
- Čoklo, M.; Maslov, D.R.; Kraljević Pavelić, S. Modulation of gut microbiota in healthy rats after exposure to nutritional supplements. Gut Microbes 2020, 12, 1779002. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequences (5′→3′) | |
|---|---|---|
| ZO-1 | Primer F | GTGGAATGATGTCGGAATA |
| Primer R | CTACAATGCGGCGATAAA | |
| Claudin-1 | Primer F | TCTTCGACTCCTTGCTGAATCTGAAC |
| Primer R | CCATCCACATCTTCTGCACCTCATC | |
| Occludin | Primer F | TGGATCTATGTACGGCTCAC |
| Primer R | CCATCTTTCTTCGGGTTT | |
| MUC1 | Primer F | AGCCACCAGTCCAGACCACAG |
| Primer R | TAGGTAGCACCGAGGAGCCATTG | |
| MUC2 | Primer F | GCTGACGAGTGGTTGGTGAATG |
| Primer R | GATGAGGTGGCAGACGGAGAC | |
| IL-1β | Primer F | CCGTGGACCTTCCAGGATGA |
| Primer R | GGGAACGTCACACACCAGCA | |
| IL-1α | Primer F | CAAACTGATGAAGCTCGTCA |
| Primer R | TCTCCTTGAGCGCTCACGAA | |
| TNF-α | Primer F | CCCTCACACTCAGATCATCTTCT |
| Primer R | GCTACGACGTGGGCTACAG | |
| GAPDH | Primer F | AGGTCGGTTGTGACGGATTTG |
| Primer R | TGTAGACCATGTAGTTGAGGTCA |
| CG | LG | AG | TG | |
|---|---|---|---|---|
| Initial Body Weight (IBW, g) | 19.20 ± 1.05 | 19.05 ± 0.75 | 18.95 ± 0.95 | 19.15 ± 1.20 |
| Final Body Weight (FBW, g) | 24.00 ± 0.95 b | 26.30 ± 0.80 a | 23.45 ± 1.00 b | 22.45 ± 0.85 b |
| Weight Gain (WG, g) | 4.80 ± 0.35 b | 7.25 ± 1.15 a | 4.50 ± 0.60 b | 3.30 ± 0.95 c |
| Weight Gain rate (WGR, %) | 24.87 ± 1.21 b | 37.49 ± 0.97 a | 24.15 ± 1.03 b | 17.62 ± 1.07 c |
| Treatment | ACE | Chao1 | Shannon | Simpson |
|---|---|---|---|---|
| TG | 278.92 ± 36.28 a | 263.75 ± 13.52 a | 0.32 ± 0.03 a | 1.56 ± 0.27 b |
| LG | 221.3 ± 29.61 b | 225.17 ± 17.02 b | 0.26 ± 0.05 a | 1.18 ± 0.16 b |
| AG | 287.06 ± 33.05 a | 280.89 ± 17.09 a | 0.29 ± 0.08 a | 1.42 ± 0.28 b |
| CG | 188.74 ± 21.05 c | 179.29 ± 16.32 c | 0.18 ± 0.04 b | 2.82 ± 0.21 a |
| Treatment | Villus Height (VH) | Crypt Depth (CD) | VH/CD |
|---|---|---|---|
| TG | 321.05 ± 19.05 c | 165.85 ± 9.55 c | 1.94 ± 0.25 b |
| LG | 463.15 ± 13.35 a | 220.50 ± 11.35 a | 2.10 ± 0.12 b |
| AG | 398.25 ± 11.55 b | 165.75 ± 10.50 c | 2.40 ± 0.23 a |
| CG | 409.20 ± 15.60 b | 198.05 ± 12.85 b | 2.07 ± 0.43 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, J.; Bao, P.; Yu, Q.; Li, N.; Ren, H.; Chen, Q.; Yan, P. Lactobacillus Re-Engineers Gut Microbiota to Overcome E. coli Colonization Resistance in Mice. Vet. Sci. 2025, 12, 484. https://doi.org/10.3390/vetsci12050484
Jia J, Bao P, Yu Q, Li N, Ren H, Chen Q, Yan P. Lactobacillus Re-Engineers Gut Microbiota to Overcome E. coli Colonization Resistance in Mice. Veterinary Sciences. 2025; 12(5):484. https://doi.org/10.3390/vetsci12050484
Chicago/Turabian StyleJia, Jianlei, Pengjia Bao, Qinran Yu, Ning Li, Hao Ren, Qian Chen, and Ping Yan. 2025. "Lactobacillus Re-Engineers Gut Microbiota to Overcome E. coli Colonization Resistance in Mice" Veterinary Sciences 12, no. 5: 484. https://doi.org/10.3390/vetsci12050484
APA StyleJia, J., Bao, P., Yu, Q., Li, N., Ren, H., Chen, Q., & Yan, P. (2025). Lactobacillus Re-Engineers Gut Microbiota to Overcome E. coli Colonization Resistance in Mice. Veterinary Sciences, 12(5), 484. https://doi.org/10.3390/vetsci12050484