
Breaking the PRRSV-2 Life Cycle in Porcine Alveolar Macrophages: Tylvalosin’s Multi-Stage Inhibition

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Reagent and Antibody Suppliers
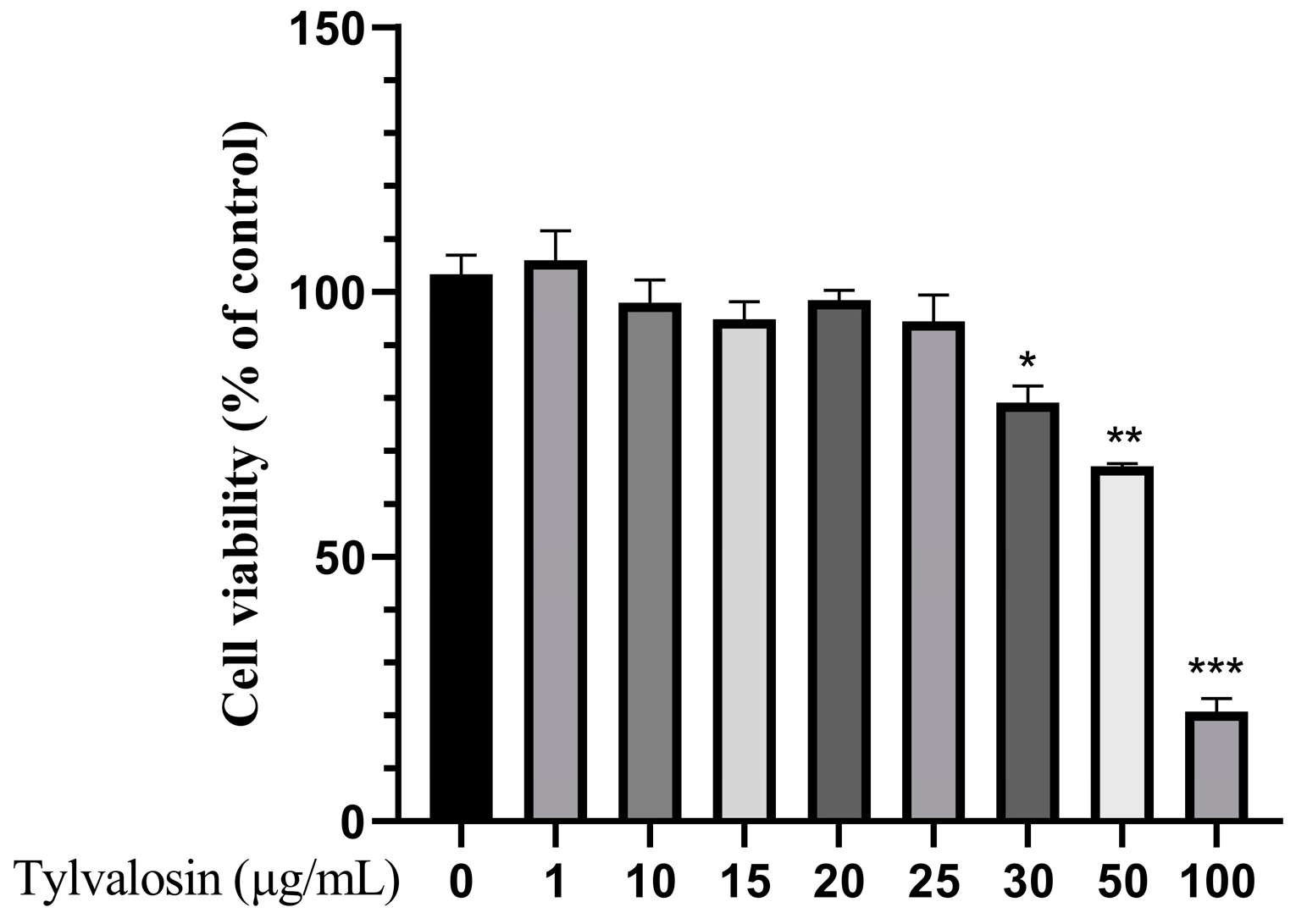
2.3. Cell Viability Assay
2.4. Assay for Testing the Inhibition of PRRSV Replication
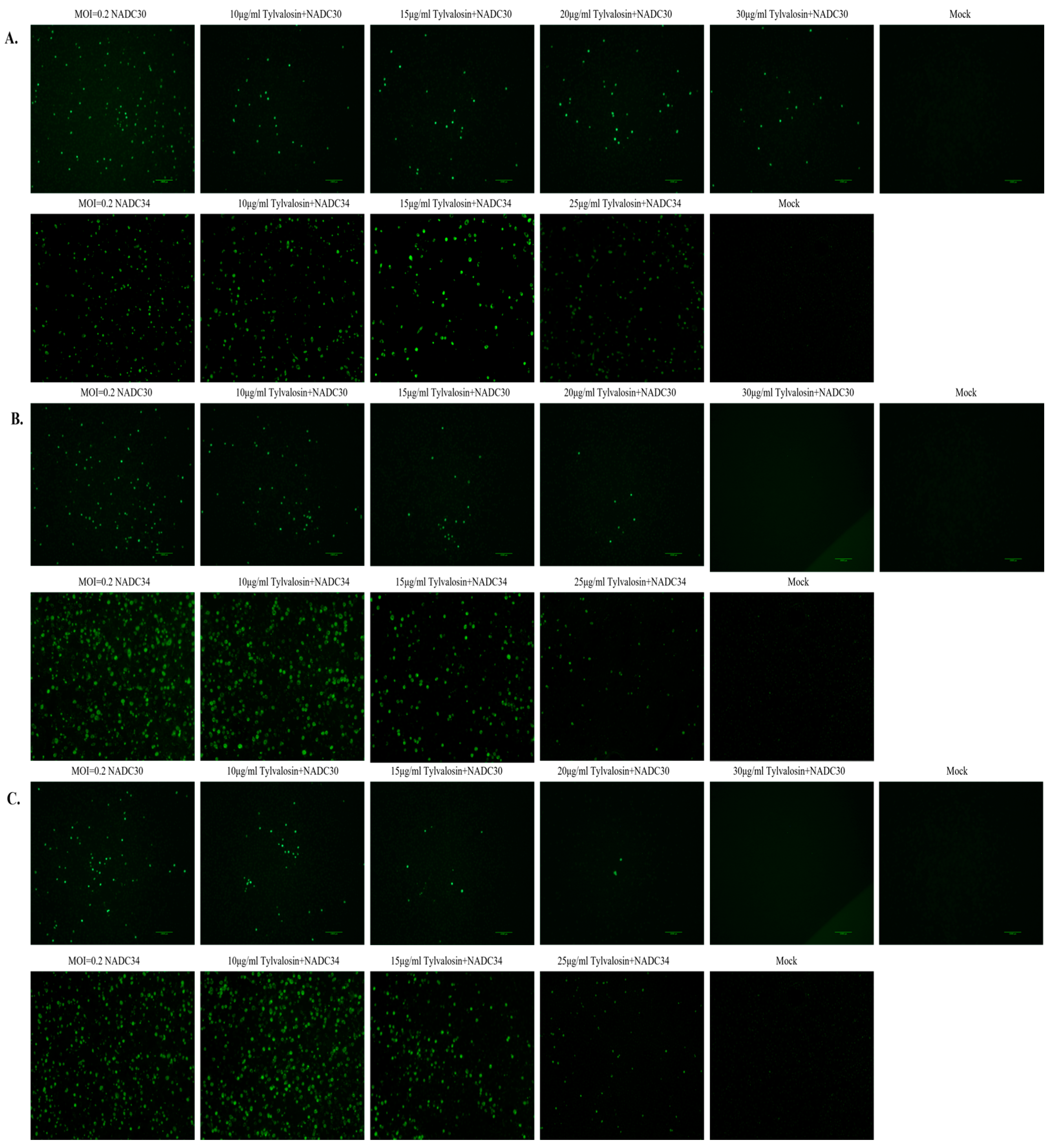
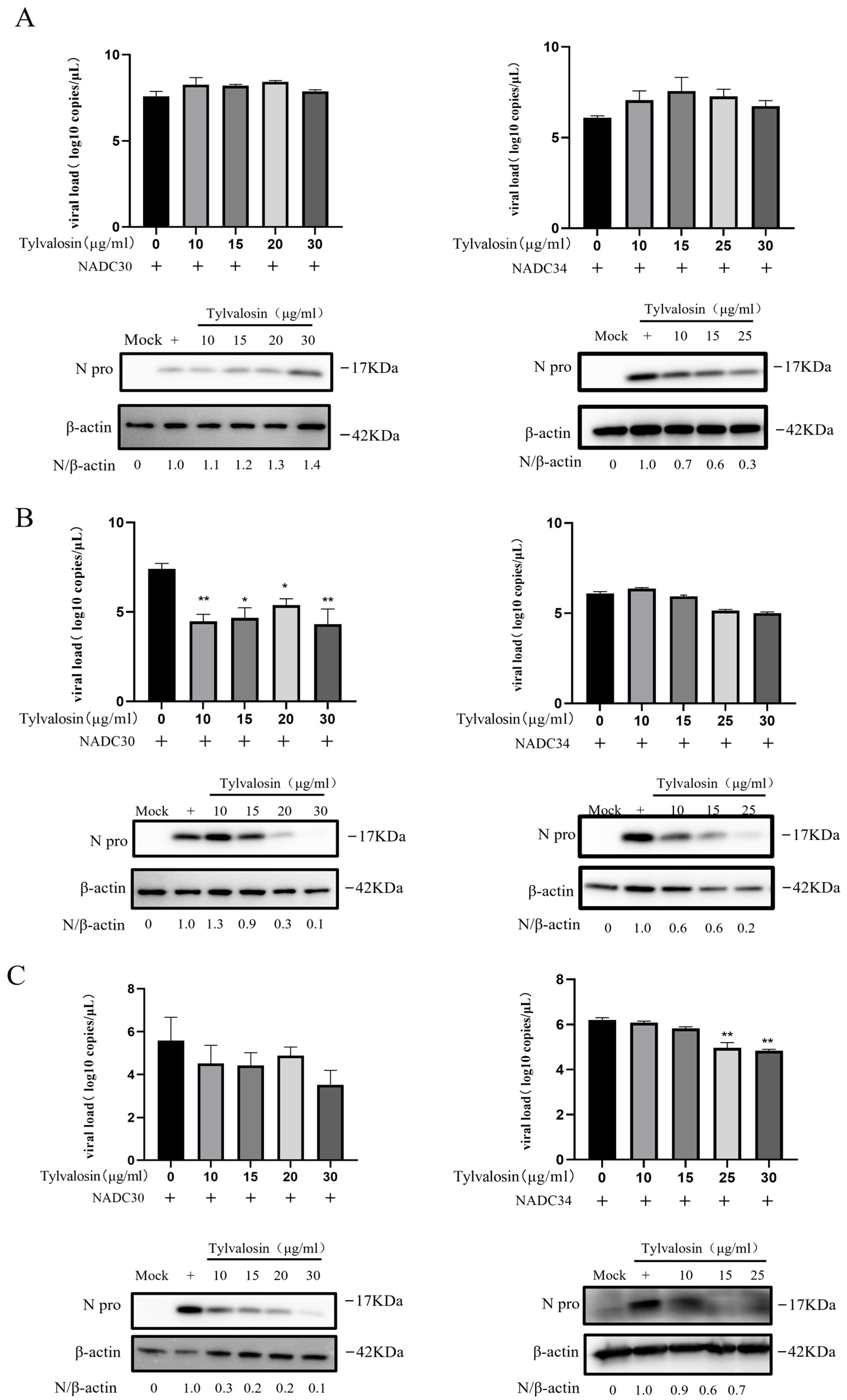
2.5. Effect of Tylvalosin on the Life Cycle of PRRSV
2.6. Immunofluorescence Assay (IFA)
2.7. Quantitative Real-Time PCR
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Cytotoxicity Test of Tylvalosin in PAMs
3.2. Tylvalosin Inhibits PRRSV Replication In Vitro
3.3. Tylvalosin Inhibits Multiple Stages of PRRSV Replication
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Benfield, D.A.; Rowland, R.R. Porcine reproductive and respiratory syndrome virus: An update on an emerging and re-emerging viral disease of swine. Virus Res. 2010, 154, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Chen, X.X.; Li, R.; Qiao, S.; Zhang, G. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Bo, K.; Wang, X.; Tang, B.; Yang, B.; Jiang, W.; Jiang, P. Emergence of a highly pathogenic porcine reproductive and respiratory syndrome virus in the Mid-Eastern region of China. Vet. J. 2007, 174, 577–584. [Google Scholar] [CrossRef]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef] [PubMed]
- Tong, G.Z.; Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; An, T.Q.; Qiu, H.J. Highly pathogenic porcine reproductive and respiratory syndrome, China. Emerg. Infect. Dis. 2007, 13, 1434–1436. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef]
- Han, G.; Lei, K.; Xu, H.; He, F. Genetic characterization of a novel recombinant PRRSV2 from lineage 8, 1 and 3 in China with significant variation in replication efficiency and cytopathic effects. Transbound. Emerg. Dis. 2020, 67, 1574–1584. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Ma, X.; Chen, X.M.; Song, Y.P.; Zheng, L.L.; Ma, S.J.; Chen, H.Y. Molecular detection and genetic characteristics of porcine reproductive and respiratory syndrome virus in central China. Microb. Pathog. 2024, 197, 107024. [Google Scholar] [CrossRef]
- Huang, B.; Xu, T.; Luo, Z.; Deng, L.; Jian, Z.; Lai, S.; Ai, Y.; Zhou, Y.; Ge, L.; Xu, Z.; et al. Prevalence and genetic diversity of PRRSV in Sichuan province of China from 2021 to 2023: Evidence of an ongoing epidemic transition. Virology 2024, 600, 110213. [Google Scholar] [CrossRef]
- Zhou, L.; Han, J.; Yang, H. The evolution and diversity of porcine reproductive and respiratory syndrome virus in China. Vet. Microbiol. 2024, 298, 110252. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.; Liu, Z.; Sun, Z.; Su, L.; Zhang, C.; Huang, L. Efficacy of two porcine reproductive and respiratory syndrome (PRRS) modified-live virus (MLV) vaccines against heterologous NADC30-like PRRS virus challenge. Vet. Microbiol. 2020, 248, 108805. [Google Scholar] [CrossRef]
- Sun, Y.F.; Zhou, L.; Bian, T.; Tian, X.X.; Ren, W.K.; Lu, C.; Zhang, L.; Li, X.L.; Cui, M.S.; Yang, H.C.; et al. Efficacy evaluation of two commercial modified-live virus vaccines against a novel recombinant type 2 porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2018, 216, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ge, X.; Yang, H. Porcine Reproductive and Respiratory Syndrome Modified Live Virus Vaccine: A “Leaky” Vaccine with Debatable Efficacy and Safety. Vaccines 2021, 9, 362. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, B.; Xu, L.; Jin, H.; Ge, X.; Guo, X.; Han, J.; Yang, H. Efficacy evaluation of three modified-live virus vaccines against a strain of porcine reproductive and respiratory syndrome virus NADC30-like. Vet. Microbiol. 2017, 207, 108–116. [Google Scholar] [CrossRef]
- Duan, E.; Wang, D.; Fang, L.; Ma, J.; Luo, J.; Chen, H.; Li, K.; Xiao, S. Suppression of porcine reproductive and respiratory syndrome virus proliferation by glycyrrhizin. Antivir. Res. 2015, 120, 122–125. [Google Scholar] [CrossRef]
- Lee, Y.J.; Lee, C. Ivermectin inhibits porcine reproductive and respiratory syndrome virus in cultured porcine alveolar macrophages. Arch. Virol. 2016, 161, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Stuart, A.D.; Brown, T.D.K.; Mockett, A.P.A. Tylvalosin, a macrolide antibiotic, nhibits the in-vitro replication of European and American porcine reproductive and respiratory syndrome virus (PRRS) viruses. Pig J. 2008, 61, 42–48. [Google Scholar]
- Zhao, Z.; Tang, X.; Zhao, X.; Zhang, M.; Zhang, W.; Hou, S.; Yuan, W.; Zhang, H.; Shi, L.; Jia, H.; et al. Tylvalosin exhibits anti-inflammatory property and attenuates acute lung injury in different models possibly through suppression of NF-κB activation. Biochem. Pharmacol. 2014, 90, 73–87. [Google Scholar] [CrossRef]
- Zhang, D.; Xia, Q.; Wu, J.; Liu, D.; Wang, X.; Niu, Z. Construction and immunogenicity of DNA vaccines encoding fusion protein of murine complement C3d-p28 and GP5 gene of porcine reproductive and respiratory syndrome virus. Vaccine 2011, 29, 629–635. [Google Scholar] [CrossRef]
- Lopez Rodriguez, A.; Berge, A.C.; Ramage, C.; Saltzman, R.; Domangue, R.J.; Gnozzio, M.J.; Muller, A.; Sierra, P.; Benchaoui, H.A. Evaluation of the clinical efficacy of a water soluble formulation of tylvalosin in the control of enzootic pneumonia associated with Mycoplasma hyopneumoniae and Pasteurella multocida in pigs. Porc. Health Manag. 2020, 6, 39. [Google Scholar] [CrossRef]
- Arsic, B.; Barber, J.; Čikoš, A.; Mladenovic, M.; Stankovic, N.; Novak, P. 16-membered macrolide antibiotics: A review. Int. J. Antimicrob. Agents 2018, 51, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Du, K.; Xia, Y.; Wu, Q.; Yin, M.; Zhao, H.; Chen, X.W. Analysis of whole transcriptome reveals the immune response to porcine reproductive and respiratory syndrome virus infection and tylvalosin tartrate treatment in the porcine alveolar macrophages. Front. Immunol. 2024, 15, 1506371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cui, C.; Zhang, S.; Deng, X.; Cai, X.; Wang, G. Tylvalosin Tartrate Improves the Health Status of Swine Herds during Immunization with Porcine Reproductive and Respiratory Syndrome Virus-Inactivated Vaccine. Vet. Sci. 2022, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Zhang, W.L.; Xiang, L.R.; Leng, C.L.; Tian, Z.J.; Tang, Y.D.; Cai, X.H. Emergence of novel porcine reproductive and respiratory syndrome viruses (ORF5 RFLP 1-7-4 viruses) in China. Vet. Microbiol. 2018, 222, 105–108. [Google Scholar] [CrossRef]
- Zhao, K.; Ye, C.; Chang, X.B.; Jiang, C.G.; Wang, S.J.; Cai, X.H.; Tong, G.Z.; Tian, Z.J.; Shi, M.; An, T.Q. Importation and Recombination Are Responsible for the Latest Emergence of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus in China. J. Virol. 2015, 89, 10712–10716. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Z.; Ding, Y.; Ge, X.; Guo, X.; Yang, H. NADC30-like Strain of Porcine Reproductive and Respiratory Syndrome Virus, China. Emerg. Infect. Dis. 2015, 21, 2256–2257. [Google Scholar] [CrossRef]
- Tang, X.; Wang, C.; Sun, W.; Wu, W.; Sun, S.; Wan, J.; Zhu, G.; Ma, N.; Ma, X.; Xu, R.; et al. Evaluating anti-viral effect of Tylvalosin tartrate on porcine reproductive and respiratory syndrome virus and analyzing the related gene regulation by transcriptomics. Virol. J. 2023, 20, 79. [Google Scholar] [CrossRef]
- Zheng, S.; An, H.; Xu, F.; Meng, F.; Cai, X.; Tian, Z.; Peng, J.; Deng, X.; Shang, Y.; Wang, G. Tylvalosin tartrate inhibits the replication stage of the porcine reproductive and respiratory syndrome virus life cycle. Microb. Pathog. 2024, 196, 106988. [Google Scholar] [CrossRef]
- Cui, M.; Qiu, M.; Yang, S.; Qiu, Y.; Qi, W.; Lin, H.; Sun, Z.; Zheng, W.; Zhu, J.; Chen, N. The replication efficacy of NADC34-like porcine reproductive and respiratory syndrome virus 2 is not directly associated with the pathogenicity. Vet. Microbiol. 2025, 301, 110367. [Google Scholar] [CrossRef]
- Yu, P.W.; Fu, P.F.; Zeng, L.; Qi, Y.L.; Li, X.Q.; Wang, Q.; Yang, G.Y.; Li, H.W.; Wang, J.; Chu, B.B.; et al. EGCG Restricts PRRSV Proliferation by Disturbing Lipid Metabolism. Microbiol. Spectr. 2022, 10, e0227621. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Shi, Y.; Xiao, S.; Li, N.; Zhao, Q.; Zhang, A.; Nan, Y.; Mu, Y.; Sun, Y.; Wu, C.; et al. Curcumin is a promising inhibitor of genotype 2 porcine reproductive and respiratory syndrome virus infection. BMC Vet. Res. 2017, 13, 298. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Meng, X.; Huang, Q.; Lei, N.; Zeng, L.; Jiang, X.; Guo, X. Spiramycin and azithromycin, safe for administration to children, exert antiviral activity against enterovirus A71 in vitro and in vivo. Int. J. Antimicrob. Agents 2019, 53, 362–369. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Primer and Probe | Primer and Probe Sequence | Amplification Position (bp) | Target Gene | Amplification Size (bp) |
|---|---|---|---|---|
| NL-PRRSV-2-F | GGACGACAAATGCGTGGTT | 14321–14339 14370–14388 | ORF6 | 68 |
| NL-PRRSV-2-R | CCACCACGTTGAAAGTGCC | ORF6 | ||
| NL-PRRSV-2-P | FAM-CTTGCCGTTATCGGATGA-MGB | 14348–14365 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, H.; Zhao, Y.; Deng, X.; Hu, W.; Zhang, X.; Zheng, S.; Yao, L.; Meng, F.; Fang, Z.; Xu, F.; et al. Breaking the PRRSV-2 Life Cycle in Porcine Alveolar Macrophages: Tylvalosin’s Multi-Stage Inhibition. Vet. Sci. 2025, 12, 348. https://doi.org/10.3390/vetsci12040348
An H, Zhao Y, Deng X, Hu W, Zhang X, Zheng S, Yao L, Meng F, Fang Z, Xu F, et al. Breaking the PRRSV-2 Life Cycle in Porcine Alveolar Macrophages: Tylvalosin’s Multi-Stage Inhibition. Veterinary Sciences. 2025; 12(4):348. https://doi.org/10.3390/vetsci12040348
Chicago/Turabian StyleAn, Hui, Yuhan Zhao, Xiaohong Deng, Wei Hu, Xia Zhang, Shuo Zheng, Longshuai Yao, Fanliang Meng, Zheng Fang, Fanghua Xu, and et al. 2025. "Breaking the PRRSV-2 Life Cycle in Porcine Alveolar Macrophages: Tylvalosin’s Multi-Stage Inhibition" Veterinary Sciences 12, no. 4: 348. https://doi.org/10.3390/vetsci12040348
APA StyleAn, H., Zhao, Y., Deng, X., Hu, W., Zhang, X., Zheng, S., Yao, L., Meng, F., Fang, Z., Xu, F., Qiu, J., Li, N., & Wang, G. (2025). Breaking the PRRSV-2 Life Cycle in Porcine Alveolar Macrophages: Tylvalosin’s Multi-Stage Inhibition. Veterinary Sciences, 12(4), 348. https://doi.org/10.3390/vetsci12040348