

An Inducible CYP19A1 Excision Model for Sexual Differentiation in Chicken (Gallus gallus) via the CRISPR/Cas9 System

 , ,
, ,  , and
, and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
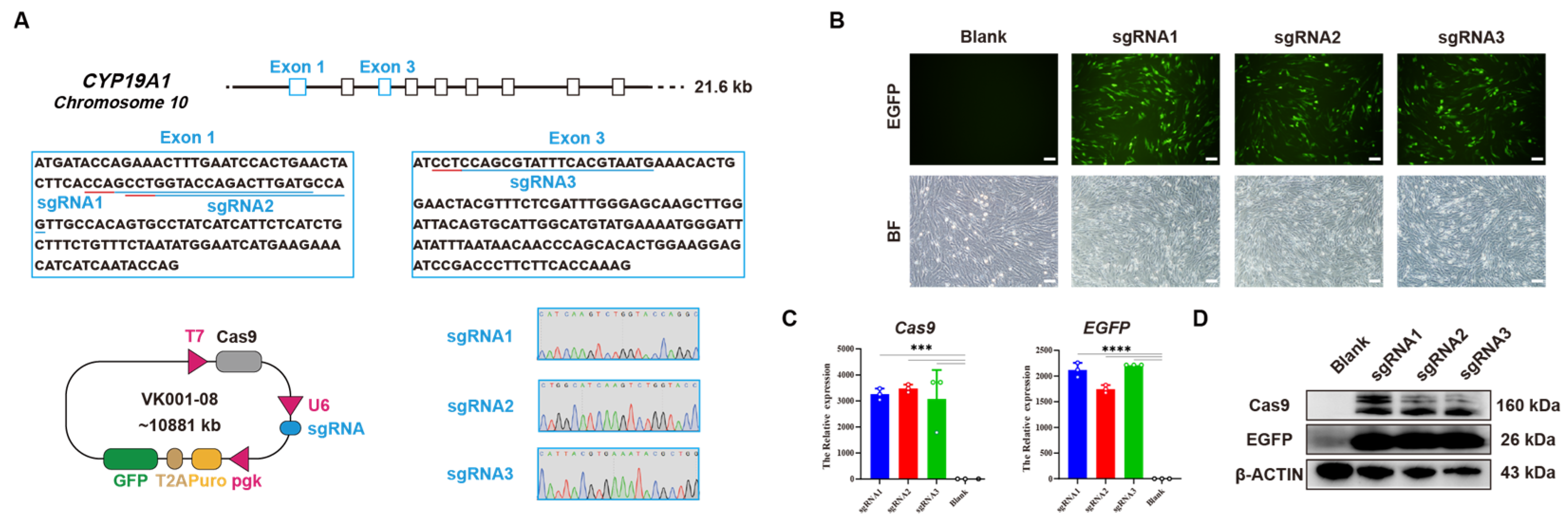
2.2. Construction of CYP19A1 Knockout Vector
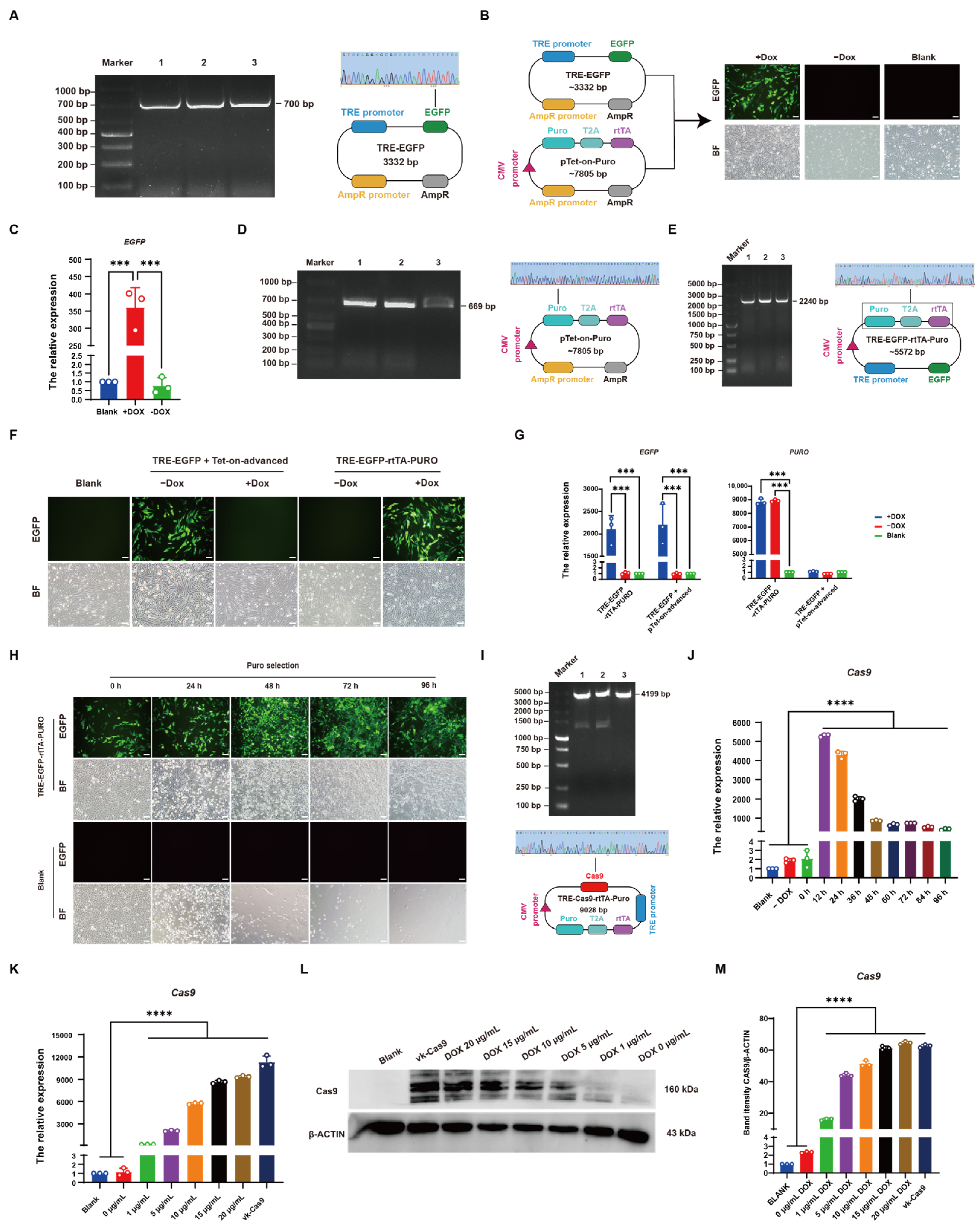
2.3. Construction of the TRE-EGFP Vector
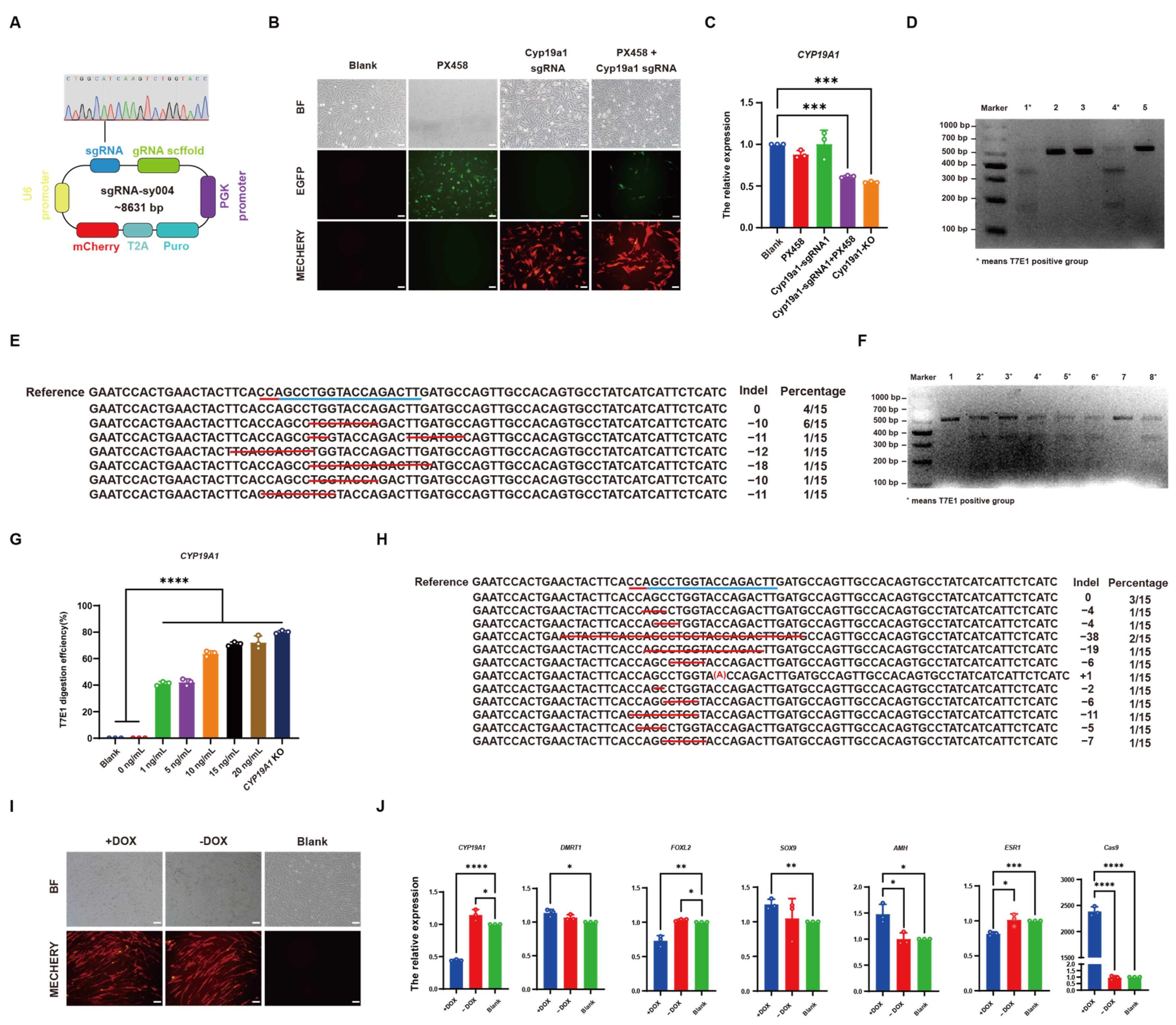
2.4. Construction of TRE-Cas9-rtTA-Puro and sgRNA-sy004 Vectors
2.5. Cell Transfection
2.6. RNA Isolation and Reverse Transcription
2.7. qRT-PCR
2.8. Western Blot (Supplementary Materials)
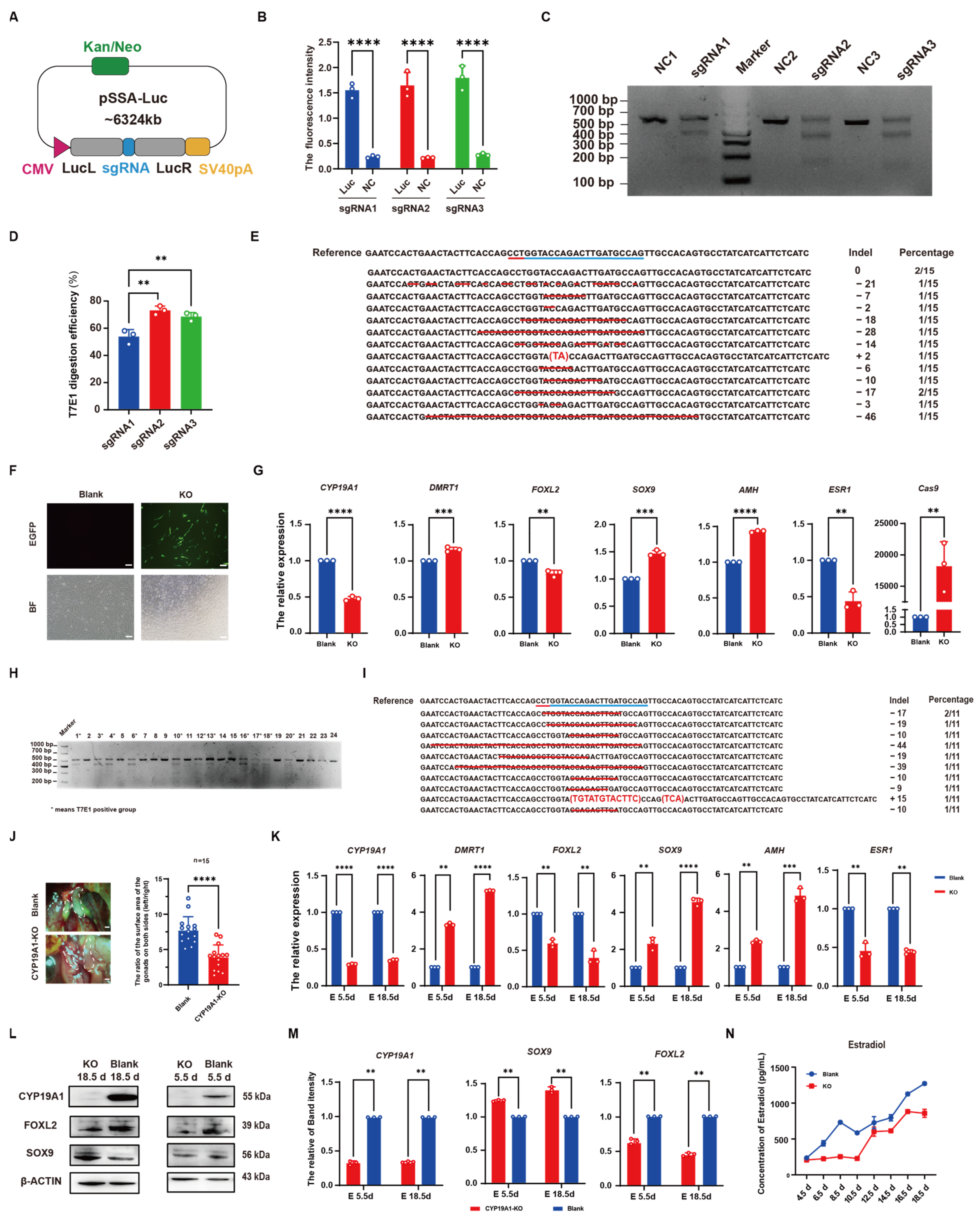
2.9. Luciferase-SSA Activity Assay
2.10. T7E1 Enzyme Digestion Assay
2.11. TA Cloning
2.12. PEI-Mediated Injection
2.13. ELISA Detection
2.14. Statistical Analysis
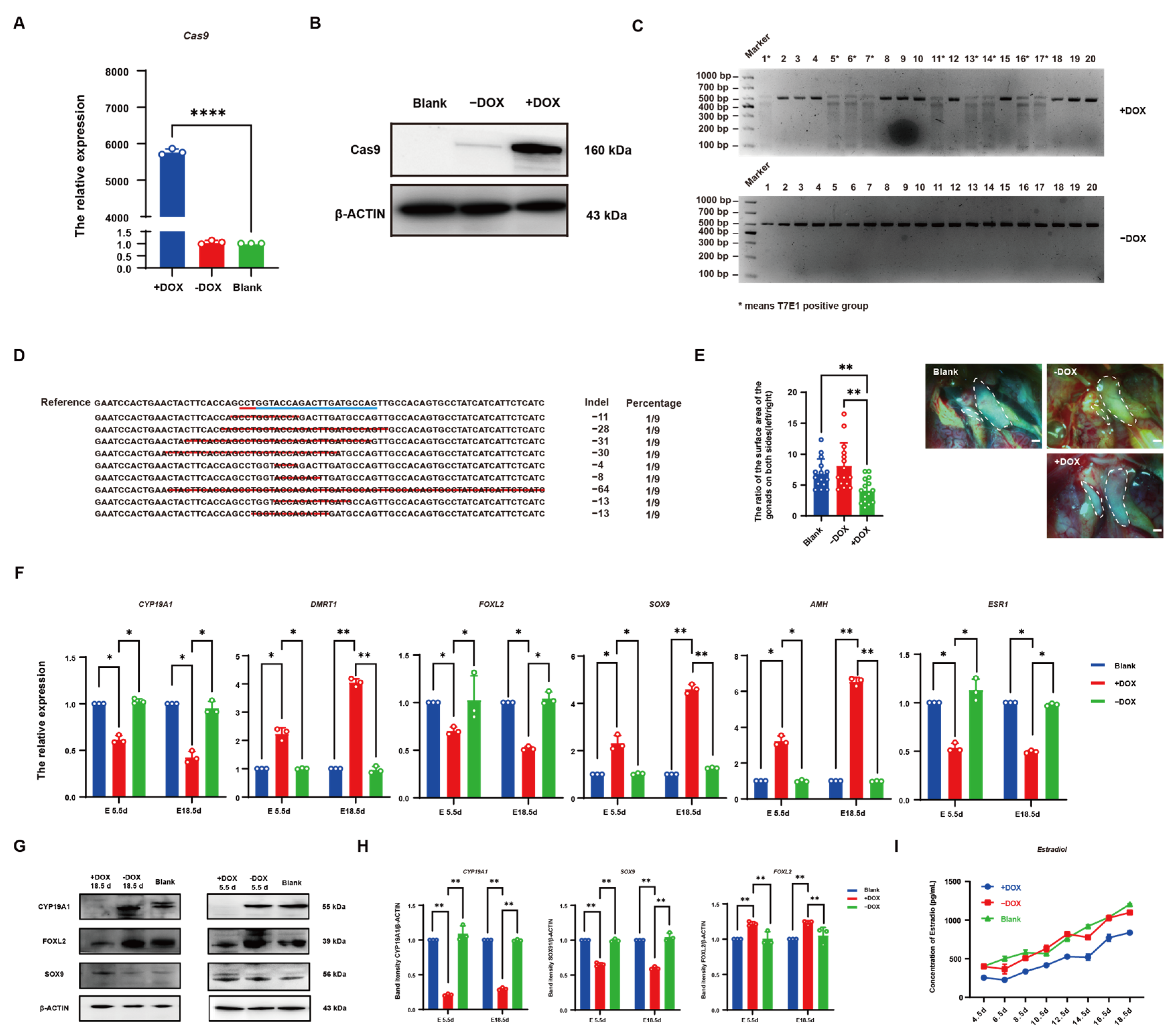
3. Results
3.1. Establishment of CYP19A1 Knockout System In Vitro
3.2. Identification of CYP19A1 Knockout System
3.3. Establishment of CRISPR/Cas9-Mediated Induced Knockout System in Chicken DF-1 Cells
3.4. Establishment of CRISPR/Cas9-Mediated DOX-Induced CYP19A1 Knockout System In Vitro
3.5. CRISPR/Cas9 Effectively Knocked out CYP19A1 via DOX Induction In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Eising, C.M.; Müller, W.; Dijkstra, C.; Groothuis, T.G.G. Maternal Androgens in Egg Yolks: Relation with Sex, Incubation Time and Embryonic Growth. Gen. Comp. Endocrinol. 2003, 132, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kobayashi, A.; Sekido, R.; DiNapoli, L.; Brennan, J.; Chaboissier, M.-C.; Poulat, F.; Behringer, R.R.; Lovell-Badge, R.; Capel, B. Fgf9 and Wnt4 Act as Antagonistic Signals to Regulate Mammalian Sex Determination. PLoS Biol. 2006, 4, e187. [Google Scholar] [CrossRef]
- Rutt, R.L.; Jakobsen, J. The ‘Brother Layer Problem’: Routine Killing, Biotechnology and the Pursuit of ‘Ethical Sustainability’ in Industrial Poultry. Environ. Plan. E Nat. Space 2023, 6, 1785–1803. [Google Scholar] [CrossRef]
- Petitte, J.N.; Kegelmeyer, A.E. Rapid Sex Determination of Chick Embryos Using the Polymerase Chain Reaction 1. Anim. Biotechnol. 1995, 6, 119–130. [Google Scholar] [CrossRef]
- Arnold, K.E.; Orr, K.J.; Griffiths, R. Primary Sex Ratios in Birds: Problems with Molecular Sex Identification of Undeveloped Eggs. Mol. Ecol. 2003, 12, 3451–3458. [Google Scholar] [CrossRef]
- Weissmann, A.; Reitemeier, S.; Hahn, A.; Gottschalk, J.; Einspanier, A. Sexing Domestic Chicken before Hatch: A New Method for in Ovo Gender Identification. Theriogenology 2013, 80, 199–205. [Google Scholar] [CrossRef]
- Zhang, W.; Pan, L.; Tu, K.; Zhang, Q.; Liu, M. Comparison of Spectral and Image Morphological Analysis for Egg Early Hatching Property Detection Based on Hyperspectral Imaging. PLoS ONE 2014, 9, e88659. [Google Scholar] [CrossRef]
- Xie, C.; Tang, W.; Yang, C. A Review of the Recent Advances for the in Ovo Sexing of Chicken Embryos Using Optical Sensing Techniques. Poult. Sci. 2023, 102, 102906. [Google Scholar] [CrossRef]
- Elbrecht, A.; Smith, R.G. Aromatase Enzyme Activity and Sex Determination in Chickens. Science 1992, 255, 467–470. [Google Scholar] [CrossRef]
- Crews, D.; Bergeron, J.M. Role of Reductase and Aromatase in Sex Determination in the Red-Eared Slider (Trachemys scripta), a Turtle with Temperature-Dependent Sex Determination. J. Endocrinol. 1994, 143, 279–289. [Google Scholar] [CrossRef]
- Kitano, T.; Takamune, K.; Nagahama, Y.; Abe, S.-I. Aromatase Inhibitor and 17β-Methyltestosterone Cause Sex-Reversal from Genetical Females to Phenotypic Males and Suppression of P450 Aromatase Gene Expression in Japanese Flounder (Paralichthys olivaceus). Mol. Reprod. Dev. 2000, 56, 1–5. [Google Scholar] [CrossRef]
- Sakata, N.; Tamori, Y.; Wakahara, M. P450 Aromatase Expression in the Temperature-Sensitive Sexual Differentiation of Salamander (Hynobius retardatus) Gonads. Int. J. Dev. Biol. 2005, 49, 417–425. [Google Scholar] [CrossRef]
- Nakabayashi, O.; Kikuchi, H.; Kikuchi, T.; Mizuno, S. Differential Expression of Genes for Aromatase and Estrogen Receptor during the Gonadal Development in Chicken Embryos. J. Mol. Endocrinol. 1998, 20, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Zuo, Q.; Song, J.; Zhang, Y.; Chen, G.; Li, B. CYP19A1 (Aromatase) Dominates Female Gonadal Differentiation in Chicken (Gallus gallus) Embryos Sexual Differentiation. Biosci. Rep. 2020, 40, BSR20201576. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, L.S.; Cummins, D.M.; Doran, T.J.; Sinclair, A.H.; Smith, C.A. Overexpression of Aromatase Alone Is Sufficient for Ovarian Development in Genetically Male Chicken Embryos. PLoS ONE 2013, 8, e68362. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef]
- Yosef, I.; Edry-Botzer, L.; Globus, R.; Shlomovitz, I.; Munitz, A.; Gerlic, M.; Qimron, U. A Genetic System for Biasing the Sex Ratio in Mice. EMBO Rep. 2019, 20, e48269. [Google Scholar] [CrossRef]
- Gamez, S.; Chaverra-Rodriguez, D.; Buchman, A.; Kandul, N.P.; Mendez-Sanchez, S.C.; Bennett, J.B.; Sánchez, C.H.M.; Yang, T.; Antoshechkin, I.; Duque, J.E.; et al. Exploiting a Y Chromosome-Linked Cas9 for Sex Selection and Gene Drive. Nat. Commun. 2021, 12, 7202. [Google Scholar] [CrossRef]
- Urlinger, S.; Baron, U.; Thellmann, M.; Hasan, M.T.; Bujard, H.; Hillen, W. Exploring the Sequence Space for Tetracycline-Dependent Transcriptional Activators: Novel Mutations Yield Expanded Range and Sensitivity. Proc. Natl. Acad. Sci. USA 2000, 97, 7963–7968. [Google Scholar] [CrossRef]
- Zhou, X.; Vink, M.; Klaver, B.; Berkhout, B.; Das, A.T. Optimization of the Tet-On System for Regulated Gene Expression through Viral Evolution. Gene Ther. 2006, 13, 1382–1390. [Google Scholar] [CrossRef]
- Peng, L.; Pan, P.; Chen, J.; Yu, X.; Wu, J.; Chen, Y. A Tetracycline-inducible CRISPR/Cas9 System, Targeting Two Long Non-coding RNAs, Suppresses the Malignant Behavior of Bladder Cancer Cells. Oncol. Lett. 2018, 16, 4309–4316. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Bai, Y.; Chen, Y.; Wang, K. Knockout of the Transducin-Like Enhancer of Split 6 Gene Affects the Proliferation and Cell Cycle Process of Mouse Spermatogonia. Int. J. Mol. Sci. 2020, 21, 5827. [Google Scholar] [CrossRef]
- Watanabe, T.; Saito, D.; Tanabe, K.; Suetsugu, R.; Nakaya, Y.; Nakagawa, S.; Takahashi, Y. Tet-on Inducible System Combined with in Ovo Electroporation Dissects Multiple Roles of Genes in Somitogenesis of Chicken Embryos. Dev. Biol. 2007, 305, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Gao, X.; Zhao, J.; Zhi, Q.; Liu, X.; Zuo, Q.; Jin, K.; Zhang, Y.; Niu, Y.; Han, W.; et al. H3K4me2 Cooperates with Wnt/TCF7L2 to Regulate TDRD1 and Promote Chicken Spermatogonia Stem Cell Formation. Poult. Sci. 2023, 102, 102552. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, S.; Lian, Z.; Jiang, J.; Gao, X.; Hu, C.; Zuo, Q.; Zhang, Y.; Chen, G.; Jin, K.; et al. Tle4z1 Facilitate the Male Sexual Differentiation of Chicken Embryos. Front. Physiol. 2022, 13, 856980. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Komatsu, T.; Kasahara, M.; Katoh-Fukui, Y.; Ogawa, H.; Toyama, Y.; Maekawa, M.; Toshimori, K.; Chandraratna, R.A.S.; Morohashi, K.; et al. Mechanism of Asymmetric Ovarian Development in Chick Embryos. Development 2008, 135, 677–685. [Google Scholar] [CrossRef]
- Smith, C.A.; Andrews, J.E.; Sinclair, A.H. Gonadal Sex Differentiation in Chicken Embryos: Expression of Estrogen Receptor and Aromatase Genes. J. Steroid Biochem. Mol. Biol. 1997, 60, 295–302. [Google Scholar] [CrossRef]
- Shimada, K. Gene Expression of Steroidogenic Enzymes in Chicken Embryonic Gonads. J. Exp. Zool. 1998, 281, 450–456. [Google Scholar] [CrossRef]
- Bruggeman, V.; Van As, P.; Decuypere, E. Developmental Endocrinology of the Reproductive Axis in the Chicken Embryo. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 839–846. [Google Scholar] [CrossRef]
- Liang, H.W.; Meng, Y.; Cao, L.H.; Li, X.; Zou, G.W. Expression and Characterization of the Cyp19a Gene and Its Responses to Estradiol/Letrozole Exposure in Chinese Soft-shelled Turtle (Pelodiscus Sinensis). Mol. Reprod. Devel. 2019, 86, 480–490. [Google Scholar] [CrossRef]
- Lau, E.S.-W.; Zhang, Z.; Qin, M.; Ge, W. Knockout of Zebrafish Ovarian Aromatase Gene (Cyp19a1a) by TALEN and CRISPR/Cas9 Leads to All-Male Offspring Due to Failed Ovarian Differentiation. Sci. Rep. 2016, 6, 37357. [Google Scholar] [CrossRef] [PubMed]
- Hudson, Q.J.; Smith, C.A.; Sinclair, A.H. Aromatase Inhibition Reduces Expression of FOXL2 in the Embryonic Chicken Ovary. Dev. Dyn. 2005, 233, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.; Zhang, J.; Wang, Y. Drug Inducible CRISPR/Cas Systems. Comput. Struct. Biotechnol. J. 2019, 17, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Kayo, D.; Kimura, S.; Yamazaki, T.; Naruse, K.; Takeuchi, H.; Ansai, S. Spatio-temporal Control of Targeted Gene Expression in Combination with CRISPR/Cas and Tet-On Systems in Medaka. Genesis 2024, 62, e23519. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sgRNA Type | Target Region | Sequences: 5′-3′ | PAM |
|---|---|---|---|
| sgRNA 1 | Exon 1 | CATCAAGTCTGGTACCAGGC | TGG |
| sgRNA 2 | Exon 1 | CTGGCATCAAGTCTGGTACC | AGG |
| sgRNA 3 | Exon 3 | CATTACGTGAAATACGCTGG | AGG |
| Gene | Primer Sequences (5′-3′) |
|---|---|
| CYP19A1-1 | F: CTCTTAGTCCATCAAGTCTGGTACCAGGC R: CTCTAAAACGCCTGGTACCAGACTTGATG |
| CYP19A1-2 | F: CTCTTAGTCCTGGCATCAAGTCTGGTACC R: CTCTAAAACGGTACCAGACTTGATGCCAG |
| CYP19A1-3 | F: CTCTTAGTCCATTACGTGAAATACGCTGG R: CTCTAAAACCCAGCGTATTTCACGTAATG |
| β-ACTIN | F: CAGCCATCTTTCTTGGGTAT R: CTGTGATCTCCTTCTGCATCC |
| AMH | F: GAAGCATTTTGGGGACTGG R: GGGTGGTAGCAGAAGCTGAG |
| SOX9 | F: AAGGAGAGCGACGAGGACAAA R: GGTCCAGTCGTAGCCCTTGA |
| FOXL2 | F: CCTCAACGAGTGCTTCATCA R: ACATCTGGCAAGAGGCGTAG |
| ESR1 | F: TAGCCTGAAGCATAGTCGTT R: TTTGGGATGAGGGAAGC |
| CYP19A1 | F: CTCCAGCAGGTTGAAGGTA R: ATGAGAATGATGATAGGCAC |
| DMRT1 | F: CCCTGCTCACTCCACG R: CACTGGTTTCCTCCCT |
| Cas9 | F: GGACATCGGCACCAAC R: CTCGTGCTTCTTATCCTCTT |
| Puro | F: CGCAGCAACAGATGGAA R: TCGGCGGTGACGGTGAA |
| CYP19A1sg1 | F: CAGCAGGTTGAAAGGTAC R: GCATATACACAATAGAAGCAAAAC |
| CYP19A1sg2 | F: CAGCAGGTTGAAAGGTAC R: GCATATACACAATAGAAGCAAAAC |
| CYP19A1sg3 | F: CATAATAGAAGCAACAATGTC R: GTGAGAGCATGTTTTTTTTG |
| EGFP-1 | F: CCGGAATTCCGCCACCATGGTGAGCAAG T R: CGCGGATCCTTACTTGTACAGCTCGTCCATGC |
| Puro-1 | F: CCGGAATTCATGACCGAGTACAAGCCC R: GCTCTAGACATTGGGCCAGGATTCTC |
| Puro+rtTA-1 | F: ACATGTGACATTGATTATTGACTAGTTATTAATAG R: ACATGTAACTTGTTTATTGCAGCTTATAATG |
| Cas9-1 | F: CGGGGTACCCCCCAAAGAAGAAGCGGAAG R: ATTTGCGGCCGCTTTCTTTTTCTTAGCTTGACCAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Peng, Y.; Li, Z.; Zhu, X.; Sun, H.; Chen, G.; Zuo, Q.; Niu, Y.; Song, J.; Han, W.; et al. An Inducible CYP19A1 Excision Model for Sexual Differentiation in Chicken (Gallus gallus) via the CRISPR/Cas9 System. Vet. Sci. 2025, 12, 296. https://doi.org/10.3390/vetsci12040296
Liu G, Peng Y, Li Z, Zhu X, Sun H, Chen G, Zuo Q, Niu Y, Song J, Han W, et al. An Inducible CYP19A1 Excision Model for Sexual Differentiation in Chicken (Gallus gallus) via the CRISPR/Cas9 System. Veterinary Sciences. 2025; 12(4):296. https://doi.org/10.3390/vetsci12040296
Chicago/Turabian StyleLiu, Guangzheng, Yixiu Peng, Zeyu Li, Xilin Zhu, Hongyan Sun, Guohong Chen, Qisheng Zuo, Yingjie Niu, Jiuzhou Song, Wei Han, and et al. 2025. "An Inducible CYP19A1 Excision Model for Sexual Differentiation in Chicken (Gallus gallus) via the CRISPR/Cas9 System" Veterinary Sciences 12, no. 4: 296. https://doi.org/10.3390/vetsci12040296
APA StyleLiu, G., Peng, Y., Li, Z., Zhu, X., Sun, H., Chen, G., Zuo, Q., Niu, Y., Song, J., Han, W., Wei, W., Li, B., & Jin, K. (2025). An Inducible CYP19A1 Excision Model for Sexual Differentiation in Chicken (Gallus gallus) via the CRISPR/Cas9 System. Veterinary Sciences, 12(4), 296. https://doi.org/10.3390/vetsci12040296