
Epigallocatechin Gallate Alleviates Lipopolysaccharide-Induced Intestinal Inflammation in Wenchang Chicken by Inhibiting the TLR4/MyD88/NF-κB Signaling Pathway

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
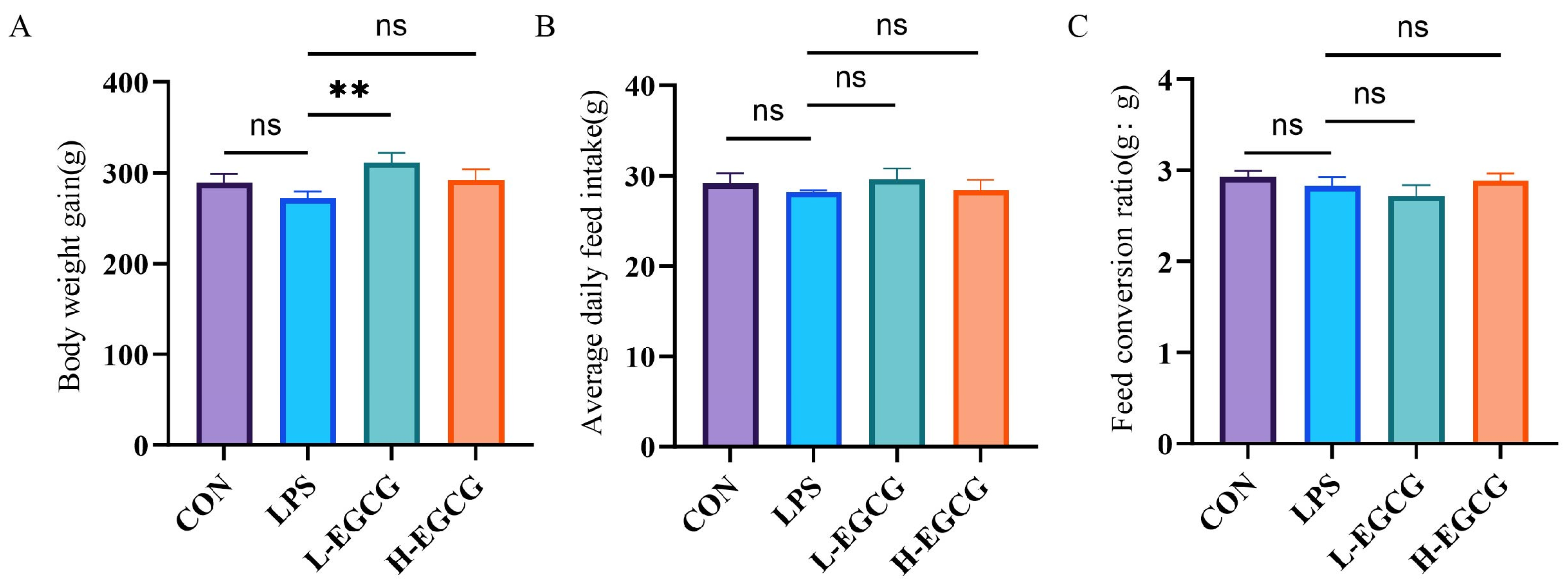
2.2. Growth Performance
2.3. Sample Collection
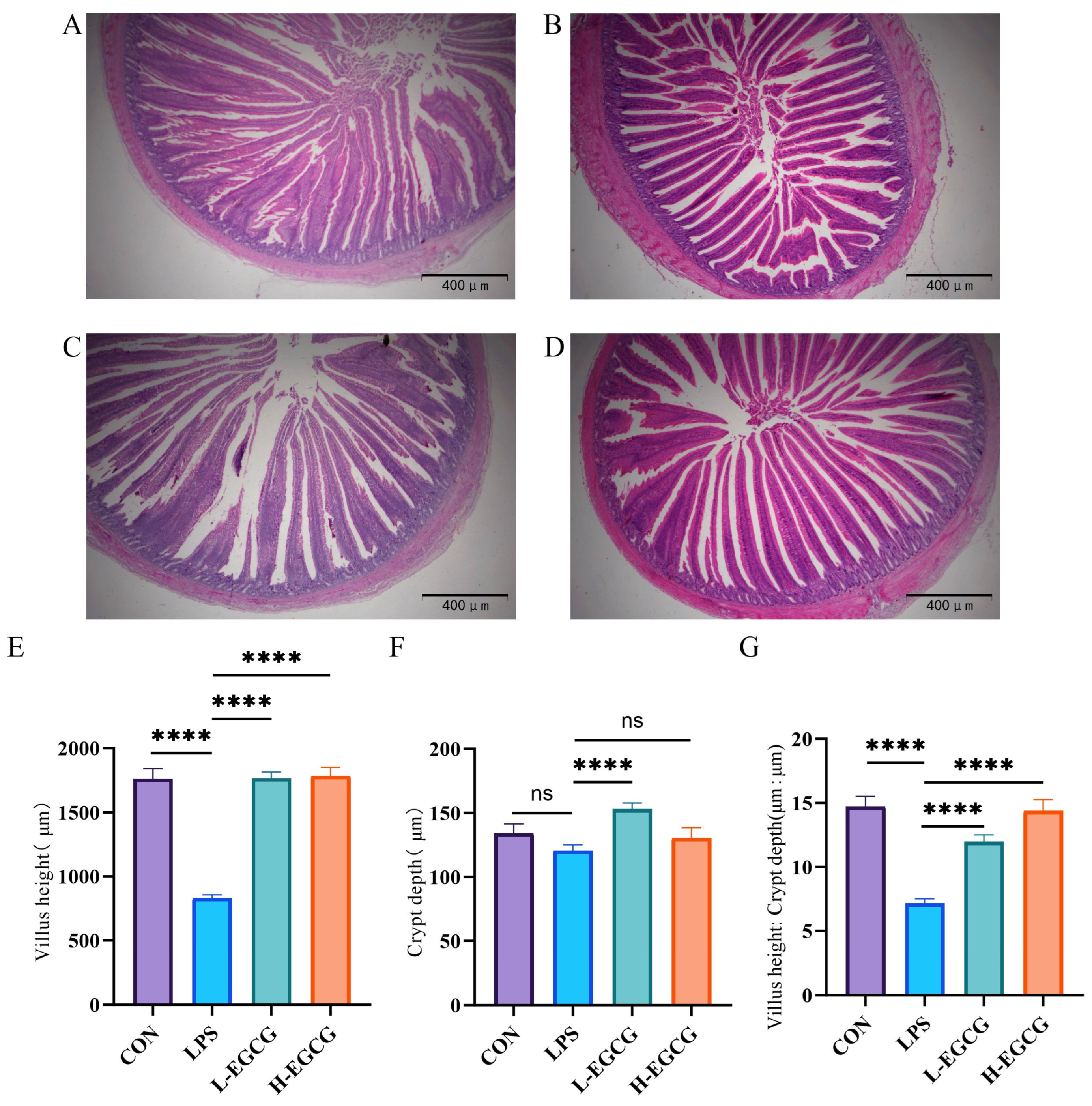
2.4. Intestinal Morphology
2.5. Antioxidant Activity
2.6. Detection of Inflammation and Assessment of Barrier Function
2.7. Total RNA Extraction and Real-Time Quantitative PCR (RT-qPCR) Analysis
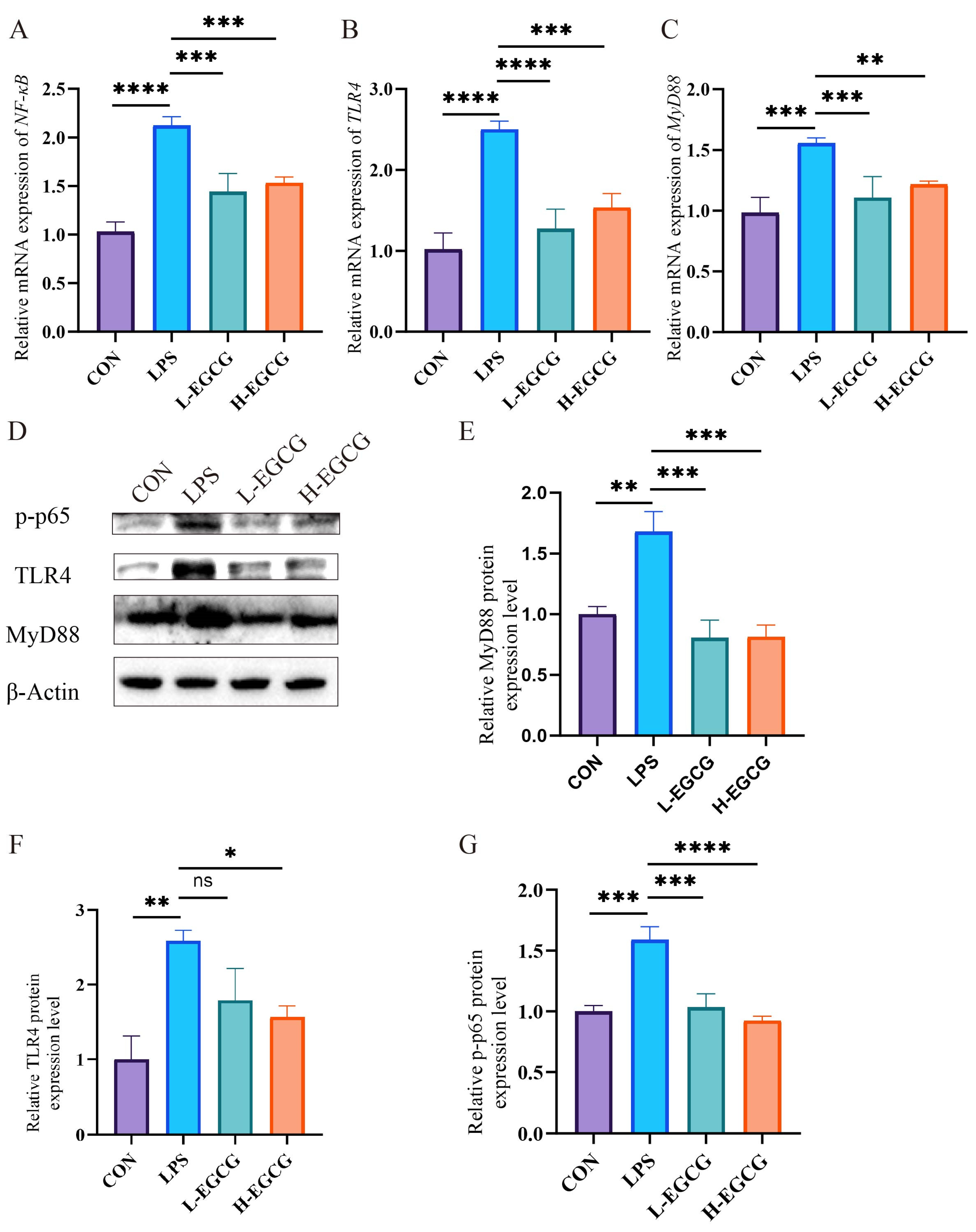
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Effects of EGCG on the Growth Performance of LPS-Challenged Wenchang Chicks
3.2. Effects of EGCG on the Jejunum Morphology of LPS-Challenged Wenchang Chicks
3.3. Effects of EGCG on the Jejunum Permeability of LPS-Challenged Wenchang Chicks
3.4. Antioxidant Status of the Serum in LPS-Challenged Wenchang Chicks
3.5. Effects of EGCG on the Inflammatory Cytokine Levels in Serum and Jejunum of LPS-Challenged Wenchang Chicks
3.6. Effects of EGCG on TLR4/MyD88/NF-κB Pathway Expression in LPS-Challenged Wenchang Chickens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, L.P.; Li, W.H.; Liu, Y.L.; Lun, J.C.; He, Y.M. Heat stress aggravates intestinal inflammation through TLR4-NF-κB signaling pathway in Ma chickens infected with Escherichia coli O157:H7. Poult. Sci. 2021, 100, 101030. [Google Scholar] [CrossRef]
- Cardoso Dal Pont, G.; Lee, A.; Bortoluzzi, C.; Farnell, Y.Z.; Gougoulias, C.; Kogut, M.H. Novel model for chronic intestinal inflammation in chickens: (2) Immunologic mechanism behind the inflammatory response. Dev. Comp. Immunol. 2023, 138, 104524. [Google Scholar] [CrossRef]
- Manjunatha, V.; Nixon, J.E.; Mathis, G.F.; Lumpkins, B.S.; Güzel-Seydim, Z.B.; Seydim, A.C.; Greene, A.K.; Jiang, X. Nigella sativa as an antibiotic alternative to promote growth and enhance health of broilers challenged with Eimeria maxima and Clostridium perfringens. Poult. Sci. 2023, 102, 102831. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 479. [Google Scholar] [CrossRef]
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern Approaches in the Discovery and Development of Plant-Based Natural Products and Their Analogues as Potential Therapeutic Agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, Y.; Chen, D.; Reddy, M.B. The Protection of EGCG Against 6-OHDA-Induced Oxidative Damage by Regulating PPARγ and Nrf2/HO-1 Signaling. Nutr. Metab. Insights 2024, 17, 11786388241253436. [Google Scholar] [CrossRef]
- Shen, M.; You, Y.; Xu, C.; Chen, Z. Epigallocatechin-3-Gallate attenuates lipopolysacharide-induced pneumonia via modification of inflammation, oxidative stress, apoptosis, and autophagy. BMC Complement. Med. Ther. 2024, 24, 147. [Google Scholar] [CrossRef]
- Wang, S.; Wang, R.; Hu, D.; Zhang, C.; Cao, P.; Huang, J.; Wang, B. Epigallocatechin gallate modulates ferroptosis through downregulation of tsRNA-13502 in non-small cell lung cancer. Cancer Cell Int. 2024, 24, 200. [Google Scholar] [CrossRef]
- Chen, B.; Li, Y.F.; Fang, Z.; Cai, W.Y.; Tian, Z.Q.; Li, D.; Wang, Z.M. Epigallocatechin-3-gallate protects sepsis-induced myocardial dysfunction by inhibiting the nuclear factor-κB signaling pathway. Heliyon 2024, 10, e27163. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, S.; Li, T.; Li, N.; Han, D.; Zhang, B.; Xu, Z.Z.; Zhang, S.; Pang, J.; Wang, S.; et al. Gut microbiota from green tea polyphenol-dosed mice improves intestinal epithelial homeostasis and ameliorates experimental colitis. Microbiome 2021, 9, 184. [Google Scholar] [CrossRef]
- Farahat, M.; Abdallah, F.; Abdel-Hamid, T.; Hernandez-Santana, A. Effect of supplementing broiler chicken diets with green tea extract on the growth performance, lipid profile, antioxidant status and immune response. Br. Poult. Sci. 2016, 57, 714–722. [Google Scholar] [CrossRef]
- Xue, B.; Song, J.; Liu, L.; Luo, J.; Tian, G.; Yang, Y. Effect of epigallocatechin gallate on growth performance and antioxidant capacity in heat-stressed broilers. Arch. Anim. Nutr. 2017, 71, 362–372. [Google Scholar] [CrossRef]
- Song, J.; Lei, X.; Luo, J.; Everaert, N.; Zhao, G.; Wen, J.; Yang, Y. The effect of Epigallocatechin-3-gallate on small intestinal morphology, antioxidant capacity and anti-inflammatory effect in heat-stressed broilers. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1030–1038. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Shen, J.; Wang, S.; Yao, J.; Yang, X. Effect of Astragalus polysaccharide and its sulfated derivative on growth performance and immune condition of lipopolysaccharide-treated broilers. Int. J. Biol. Macromol. 2015, 76, 188–194. [Google Scholar] [CrossRef]
- Liu, H.; Meng, H.; Du, M.; Lv, H.; Wang, Y.; Zhang, K. Chlorogenic acid ameliorates intestinal inflammation by inhibiting NF-κB and endoplasmic reticulum stress in lipopolysaccharide-challenged broilers. Poult. Sci. 2024, 103, 103586. [Google Scholar] [CrossRef]
- Zheng, L.; Han, Z.; Zhang, J.; Kang, J.; Li, C.; Pang, Q.; Liu, S. Lactiplantibacillus plantarum and Saccharomyces cerevisiae-Fermented Coconut Water Alleviates Dextran Sodium Sulfate-Induced Enteritis in Wenchang Chicken: A Gut Microbiota and Metabolomic Approach. Animals 2024, 14, 575. [Google Scholar] [CrossRef]
- Fang, X.; Nong, K.; Qin, X.; Liu, Z.; Gao, F.; Jing, Y.; Fan, H.; Wang, Z.; Wang, X.; Zhang, H. Effect of purple sweet potato-derived anthocyanins on heat stress response in Wenchang chickens and preliminary mechanism study. Poult. Sci. 2023, 102, 102861. [Google Scholar] [CrossRef]
- Zhang, X.; Akhtar, M.; Chen, Y.; Ma, Z.; Liang, Y.; Shi, D.; Cheng, R.; Cui, L.; Hu, Y.; Nafady, A.A.; et al. Correction: Chicken jejunal microbiota improves growth performance by mitigating intestinal inflammation. Microbiome 2022, 10, 116. [Google Scholar] [CrossRef]
- Riva, F.; Eckersall, P.D.; Chadwick, C.C.; Chadwick, L.C.; McKeegan, D.E.F.; Peinado-Izaguerri, J.; Bruggeman, G.; Hermans, D.; McLaughlin, M.; Bain, M. Identification of novel biomarkers of acute phase response in chickens challenged with Escherichia coli lipopolysaccharide endotoxin. BMC Vet. Res. 2024, 20, 236. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, S.; Wan, S.; Li, Z.; Li, H.; Tang, S. Anti-inflammatory properties of Bacillus pumilus TS1 in lipopolysaccharide-induced inflammatory damage in broilers. Anim. Biotechnol. 2024, 35, 2418516. [Google Scholar] [CrossRef]
- Jelveh, K.; Mottaghitalab, M.; Mohammadi, M. Effects of green tea phytosome on growth performance and intestinal integrity under coccidiosis infection challenge in broilers. Poult. Sci. 2023, 102, 102627. [Google Scholar] [CrossRef]
- Kiess, A.; Hirai, J.; Triplett, M.; Parker, H.; McDaniel, C.J.P.S. Impact of oral Lactobacillus acidophilus gavage on rooster seminal and cloacal Lactobacilli concentrations. Poult. Sci. 2016, 95, 1934–1938. [Google Scholar] [CrossRef]
- Chi, X.; Zhang, Y.; Ma, X.; Lu, M.; Li, Z.; Xu, W.; Hu, S. Antioxidative stress of oral administration of tea extract granule in chickens. Poult. Sci. 2020, 99, 1956–1966. [Google Scholar] [CrossRef]
- Li, H.L.; Li, Z.J.; Wei, Z.S.; Liu, T.; Zou, X.Z.; Liao, Y.; Luo, Y. Long-term effects of oral tea polyphenols and Lactobacillus brevis M8 on biochemical parameters, digestive enzymes, and cytokines expression in broilers. J. Zhejiang Univ. Sci. B 2015, 16, 1019–1026. [Google Scholar] [CrossRef]
- Fu, G.; Zhang, M.; Huang, Y.; Han, R.; Qi, K.; Yin, L.; Zhao, D.; Huang, Y.; Ma, T.; Wang, L. Effects of different addition levels of CHM-JM113 on growth performance, antioxidant capacity, organ index, and intestinal health of AA broilers. Front. Vet. Sci. 2024, 11, 1388173. [Google Scholar] [CrossRef]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef]
- Shin, W.; Kim, H.J. Intestinal barrier dysfunction orchestrates the onset of inflammatory host-microbiome cross-talk in a human gut inflammation-on-a-chip. Proc. Natl. Acad. Sci. USA 2018, 115, e10539–e10547. [Google Scholar] [CrossRef]
- Zhang, R.; Bai, D.; Zhen, W.; Hu, X.; Zhang, H.; Zhong, J.; Zhang, Y.; Ito, K.; Zhang, B.; Yang, Y.; et al. Aspirin eugenol ester affects ileal barrier function, inflammatory response and microbiota in broilers under lipopolysaccharide-induced immune stress conditions. Front. Vet. Sci. 2024, 11, 1401909. [Google Scholar] [CrossRef]
- Huangfu, W.; Ma, J.; Zhang, Y.; Liu, M.; Liu, B.; Zhao, J.; Wang, Z.; Shi, Y. Dietary Fiber-Derived Butyrate Alleviates Piglet Weaning Stress by Modulating the TLR4/MyD88/NF-κB Pathway. Nutrients 2024, 16, 1714. [Google Scholar] [CrossRef]
- Huang, T.; Che, Q.; Chen, X.; Chen, D.; Yu, B.; He, J.; Chen, H.; Yan, H.; Zheng, P.; Luo, Y.; et al. Apple Polyphenols Improve Intestinal Antioxidant Capacity and Barrier Function by Activating the Nrf2/Keap1 Signaling Pathway in a Pig Model. J. Agric. Food Chem. 2022, 70, 7576–7585. [Google Scholar] [CrossRef]
- Lei, Y.; Sun, W.; Xu, T.; Shan, J.; Gao, M.; Lin, H. Selenomethionine modulates the JAK2/STAT3/A20 pathway through oxidative stress to alleviate LPS-induced pyroptosis and inflammation in chicken hearts. Biochim. Biophys. Acta Gen. Subj. 2024, 1868, 130564. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, D.; Chen, Y. Preparation and Antioxidant Activity of Green Tea Extract Enriched in Epigallocatechin (EGC) and Epigallocatechin Gallate (EGCG). J. Agric. Food Chem. 2009, 57, 1349–1353. [Google Scholar] [CrossRef]
- Xin, H.; Yang, C.; Hua, R.; Xiao-Fen, P. Antioxidation Function of EGCG by Activating Nrf2/HO-1 Pathway in Mice with Coronary Heart Disease. Contrast Media Mol. Imaging 2022, 1, 8639139. [Google Scholar]
- Lee, J.; Heo, J.B.; Cho, S.; Ryu, C.W.; Heo, H.J.; Yun, M.Y.; Nam, G.; Song, G.Y.; Bae, J.S. Inhibitory Effects of Decursin Derivative against Lipopolysaccharide-Induced Inflammation. Pharmaceuticals 2024, 17, 1337. [Google Scholar] [CrossRef]
- Allard, R.L.; Mayfield, J.; Barchiesi, R.; Salem, N.A.; Mayfield, R.D. Toll-like receptor 7: A novel neuroimmune target to reduce excessive alcohol consumption. Neurobiol. Stress 2024, 31, 100639. [Google Scholar] [CrossRef]
- Yu, L.; Gao, F.; Li, Y.; Su, D.; Han, L.; Li, Y.; Zhang, X.; Feng, Z. Role of pattern recognition receptors in the development of MASLD and potential therapeutic applications. Biomed. Pharmacother. 2024, 175, 116724. [Google Scholar] [CrossRef]
- Zhao, W.; Shen, T.; Zhao, B.; Li, M.; Deng, Z.; Huo, Y.; Aernouts, B.; Loor, J.J.; Psifidi, A.; Xu, C. Epigallocatechin-3-gallate protects bovine ruminal epithelial cells against lipopolysaccharide-induced inflammatory damage by activating autophagy. J. Anim. Sci. Biotechnol. 2024, 15, 109. [Google Scholar] [CrossRef]
- Liao, X.; Song, X.; Li, J.; Li, L.; Fan, X.; Qin, Q.; Zhong, C.; Yang, P.; Zhan, J.; Cai, Y. An injectable co-assembled hydrogel blocks reactive oxygen species and inflammation cycle resisting myocardial ischemia-reperfusion injury. Acta Biomater. 2022, 149, 82–95. [Google Scholar] [CrossRef]
- Shao, Y.; Zhang, Y.; Zou, S.; Wang, J.; Li, X.; Qin, M.; Sun, L.; Yin, W.; Chang, X.; Wang, S.; et al. (-)-Epigallocatechin 3-gallate protects pancreatic β-cell against excessive autophagy-induced injury through promoting FTO degradation. Autophagy 2024, 20, 2460–2477. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content (%) |
|---|---|
| Corn | 52.50 |
| Soybean meal | 40.00 |
| Soybean oil | 3.00 |
| Dicalcium phosphate | 1.90 |
| Limestone powder | 1.08 |
| Salt | 0.37 |
| Lysine | 0.05 |
| Methionine | 0.19 |
| Premix (1) | 0.80 |
| Choline chloride | 0.11 |
| Total | 100.00 |
| Nutritional levels (2) | |
| Metabolizable energy (Kcal/kg) | 2966.47 |
| Crude protein | 21.77 |
| Calcium | 1.0 |
| Available phosphorus | 0.44 |
| Lysine | 1.34 |
| Methionine | 0.55 |
| Cystine | 0.4 |
| Gene (1) | Primer Sequences (5′ to 3′) | Length | Accession No. |
|---|---|---|---|
| β-actin | F-CAGCCAGCCATGGATGATGA | 150 | NM_205518.2 |
| R-CATACCAACCATCACACCCTGA | |||
| ZO-1 | F-CACTAGAGGATGAGGAGGAAGAAGAC | 80 | XM_040706827.2 |
| R-TACGCCACCATTGCTGTTGAATAC | |||
| Occludin | F-CTTCGCCTGCGTCGCTTCC | 104 | NM_205128.1 |
| R-TGCCGTAGTAGTTGGAGCCGTAG | |||
| Claudin-1 | F-GGTATGGCAACAGAGTGGCT | 91 | NM_001013611.2 |
| R-CAGCCAATGAAGAGGGCTGA | |||
| NF-κB | F-ACTTCTGGTGAAACACGGGG | 80 | NM_001001472.3 |
| R-CTCGTCCCCATGTAAGCTGG | |||
| MyD88 | F-AGCGTGCCAAAGACTTCAGA | 201 | NM_001030962.5 |
| R-ACACGTTCCTGGCAAGACAT | |||
| TLR4 | F-TGACCTACCCATCGGACACT | 111 | NM_001030693.2 |
| R-TGCCTGAGAGAGGTCAGGTT |
| Item (1) | Treatments | SEM | p-Value (2) | |||
|---|---|---|---|---|---|---|
| z | CON | LPS | L-EGCG | H-EGCG | ||
| T-AOC (U/mL) | 3.8 bc | 2.9 c | 9.9 a | 5.8 b | 0.67 | <0.001 |
| GSH-Px (U/mL) | 1755 | 1677 | 1746 | 1834 | 25.10 | 0.172 |
| T-SOD (U/mL) | 388 | 370 | 412 | 421 | 8.42 | 0.124 |
| CAT (U/mL) | 32.7 a | 7.6 c | 15.3 b | 14.8 b | 1.98 | <0.001 |
| MDA (nmol/mL) | 3.6 ab | 5.4 a | 2.2 b | 2.6 b | 0.40 | 0.027 |
| Item (1) | Treatments | SEM | p-Value (2) | |||
|---|---|---|---|---|---|---|
| z | CON | LPS | L-EGCG | H-EGCG | ||
| Serum | ||||||
| IL-1β (pg/mL) | 68.7 c | 95.8 a | 79.7 b | 83.0 b | 2.31 | <0.001 |
| TNF-α (pg/mL) | 10.9 c | 14.3 a | 12.0 bc | 12.7 b | 0.30 | <0.001 |
| IL-10 (pg/mL) | 9.3 a | 6.2 c | 8.2 ab | 7.5 b | 0.27 | <0.001 |
| Jejunum | ||||||
| IL-1β (pg/mg) | 53.1 c | 90.0 a | 66.4 b | 76.2 b | 3.03 | <0.001 |
| TNF-α (pg/mg) | 6.1 c | 11.0 a | 6.9 c | 8.8 b | 0.38 | <0.001 |
| IL-10 (pg/mg) | 13.1 a | 8.4 c | 12.4 a | 9.9 b | 0.40 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Li, K.; Liu, Q.; Zhang, H.; Li, L. Epigallocatechin Gallate Alleviates Lipopolysaccharide-Induced Intestinal Inflammation in Wenchang Chicken by Inhibiting the TLR4/MyD88/NF-κB Signaling Pathway. Vet. Sci. 2025, 12, 225. https://doi.org/10.3390/vetsci12030225
Xu X, Li K, Liu Q, Zhang H, Li L. Epigallocatechin Gallate Alleviates Lipopolysaccharide-Induced Intestinal Inflammation in Wenchang Chicken by Inhibiting the TLR4/MyD88/NF-κB Signaling Pathway. Veterinary Sciences. 2025; 12(3):225. https://doi.org/10.3390/vetsci12030225
Chicago/Turabian StyleXu, Xin, Kunpeng Li, Qian Liu, Haiwen Zhang, and Lianbin Li. 2025. "Epigallocatechin Gallate Alleviates Lipopolysaccharide-Induced Intestinal Inflammation in Wenchang Chicken by Inhibiting the TLR4/MyD88/NF-κB Signaling Pathway" Veterinary Sciences 12, no. 3: 225. https://doi.org/10.3390/vetsci12030225
APA StyleXu, X., Li, K., Liu, Q., Zhang, H., & Li, L. (2025). Epigallocatechin Gallate Alleviates Lipopolysaccharide-Induced Intestinal Inflammation in Wenchang Chicken by Inhibiting the TLR4/MyD88/NF-κB Signaling Pathway. Veterinary Sciences, 12(3), 225. https://doi.org/10.3390/vetsci12030225