

In Vivo Study of Inoculation Approaches and Pathogenicity in African Swine Fever


Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Isolation
2.2. qPCR
2.3. Hemadsorption Assay
2.4. Immunofluorescence Assay
2.5. Electron Microscopy
2.6. Virus Growth Titration
2.7. Animal Experiments
2.8. Histopathological and Immunohistochemical Analysis
3. Results
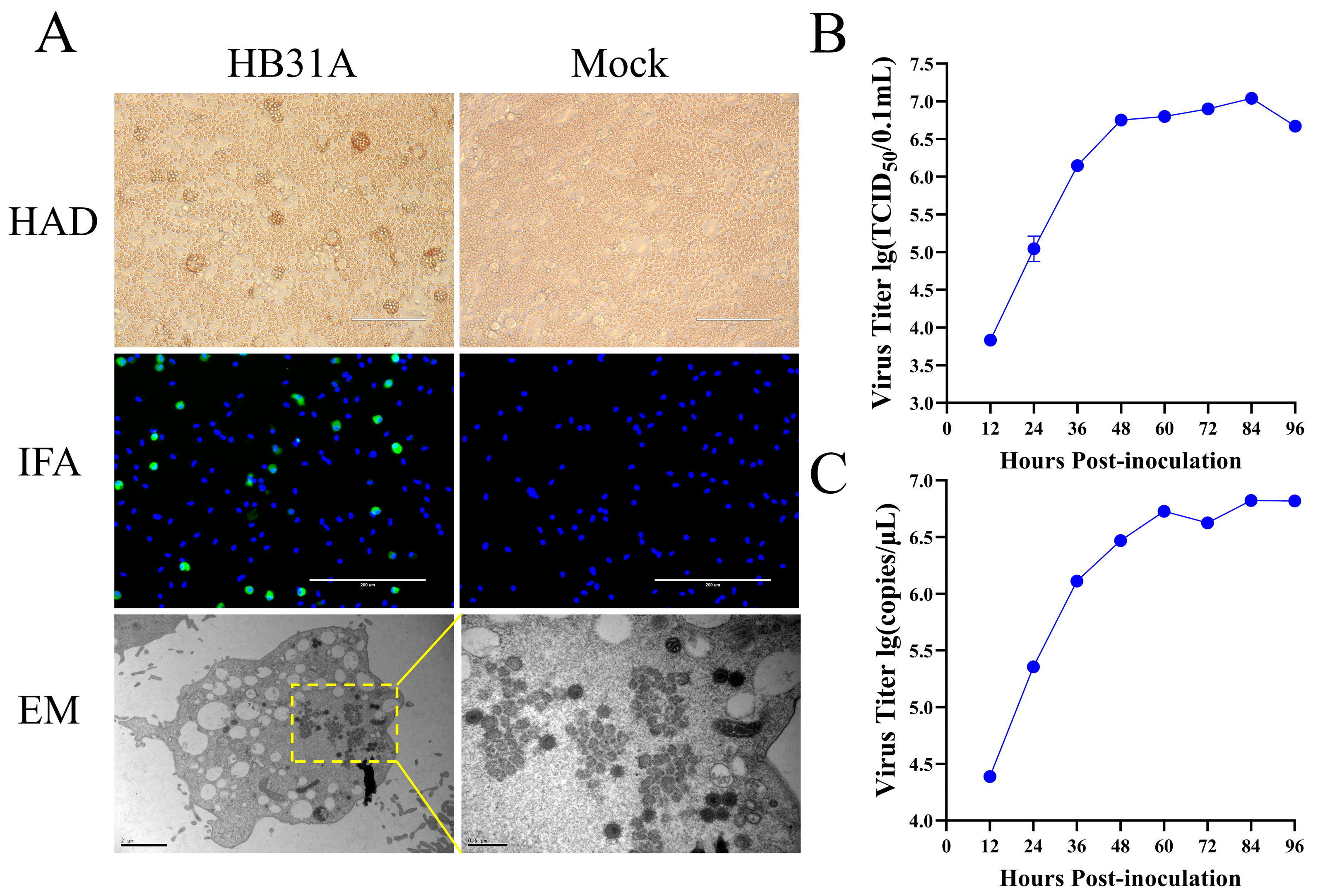
3.1. Isolation and Characterization of HB31A In Vitro
3.2. The Pathogenicity of HB31A in Pigs via Intramuscular and Oronasal Inoculations
3.3. Viral Shedding of HB31A in Inoculated Pigs
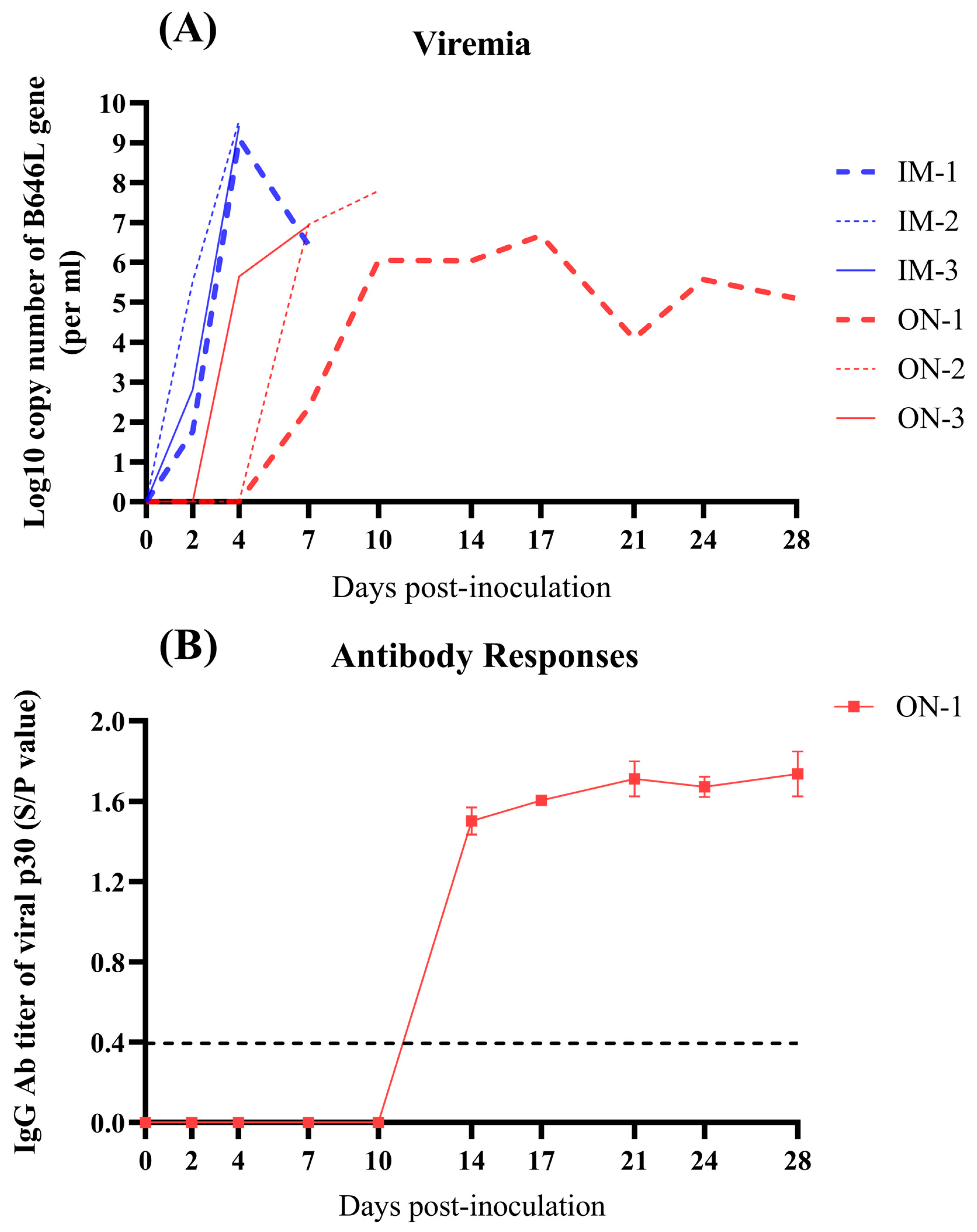
3.4. Viremia and ASFV-Specific Antibody Responses in Inoculated Pigs
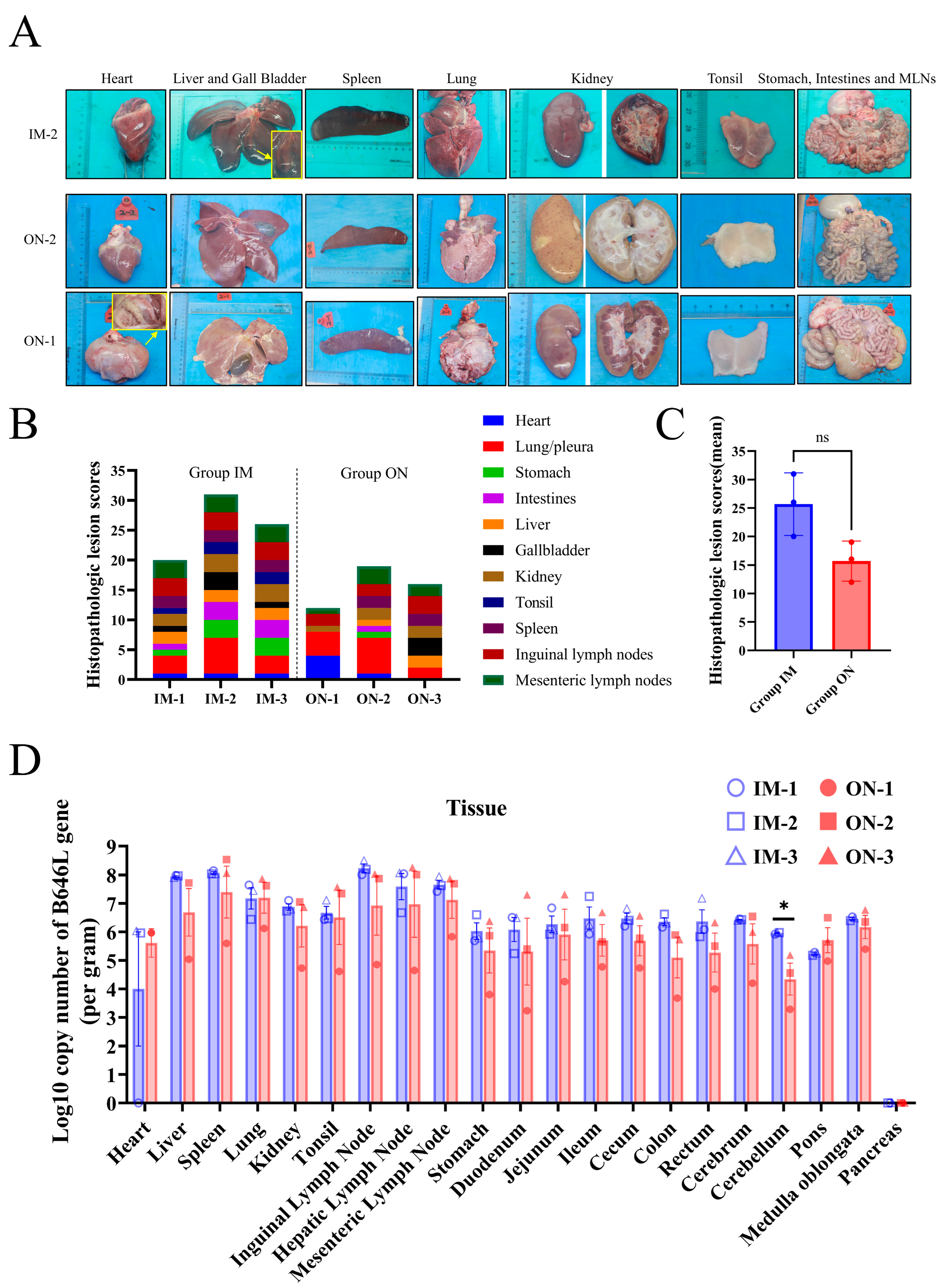
3.5. Lesions and Virus Replication in Tissues
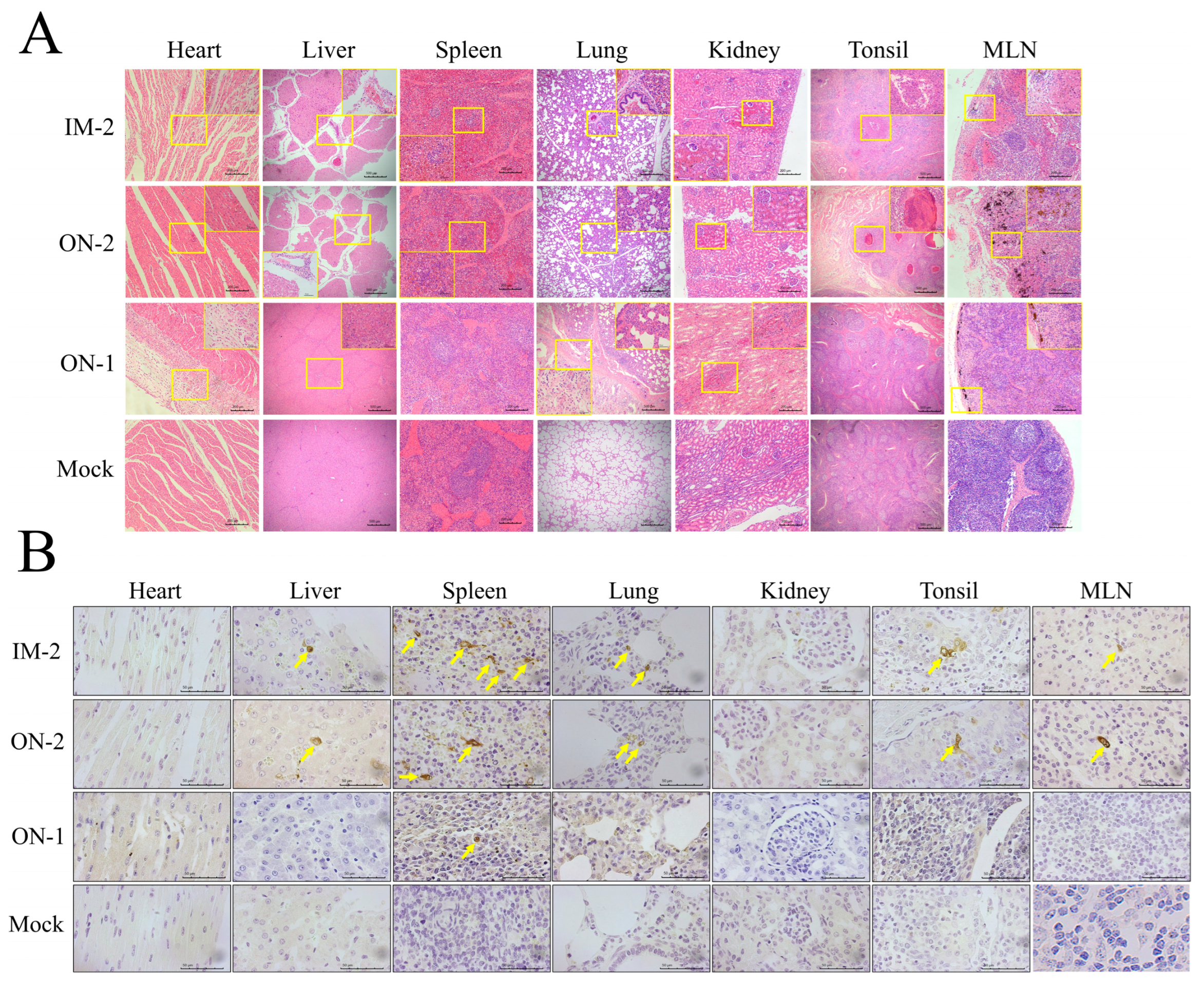
3.6. Histopathological Evaluations and Viral Antigen Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Revilla, Y.; Pérez-Núñez, D.; Richt, J.A. African Swine Fever Virus Biology and Vaccine Approaches. Adv. Virus Res. 2018, 100, 41–74. [Google Scholar] [CrossRef]
- Yáñez, R.J.; Rodríguez, J.M.; Nogal, M.L.; Yuste, L.; Enríquez, C.; Rodriguez, J.F.; Viñuela, E. Analysis of the complete nucleotide sequence of African swine fever virus. Virology 1995, 208, 249–278. [Google Scholar] [CrossRef]
- de Villiers, E.P.; Gallardo, C.; Arias, M.; Da Silva, M.; Upton, C.; Martin, R.; Bishop, R.P. Phylogenomic analysis of 11 complete African swine fever virus genome sequences. Virology 2010, 400, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.A.G.; Tcherepanov, V.; Upton, C.; Dixon, L.K. Comparison of the genome sequences of non-pathogenic and pathogenic African swine fever virus isolates. J. General. Virol. 2008, 89 Pt 2, 397–408. [Google Scholar] [CrossRef]
- Dixon, L.K.; Chapman, D.A.G.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Nurmoja, I.; Cano-Gómez, C.; Cvetkova, S.; Frant, M.; Woźniakowski, G.; Simón, A.; Pérez, C.; Nieto, R.; et al. Dynamics of African swine fever virus (ASFV) infection in domestic pigs infected with virulent, moderate virulent and attenuated genotype II ASFV European isolates. Transbound. Emerg. Dis. 2021, 68, 2826–2841. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Zhang, Z.; Wang, Z.; He, X.; Zhang, X.; Wang, L.; Wang, W.; Huang, L.; Xi, F.; Huangfu, H.; et al. Emergence and prevalence of naturally occurring lower virulent African swine fever viruses in domestic pigs in China in 2020. Sci. China Life Sci. 2021, 64, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Iacolina, L.; Penrith, M.L.; Bellini, S.; Chenais, E.; Gavier-Widén, D. Understanding and Combatting African Swine Fever—A European Perspective; Wageningen Academic Publishers: Wageningen, The Netherlands, 2021; p. 310. [Google Scholar] [CrossRef]
- Arzt, J.; White, W.R.; Thomsen, B.V.; Brown, C.C. Agricultural diseases on the move early in the third millennium. Vet. Pathol. 2010, 47, 15–27. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, R.; Zhang, X.; Li, F.; Wang, J.; Zhang, J.; Liu, X.; Wang, L.; Zhang, J.; Wu, X.; et al. Replication and virulence in pigs of the first African swine fever virus isolated in China. Emerg. Microbes Infect. 2019, 8, 438–447. [Google Scholar] [CrossRef]
- Oura, C.A.; Powell, P.P.; Parkhouse, R.M. Detection of African swine fever virus in infected pig tissues by immunocytochemistry and in sity hybridisation. J. Virol. Methods 1998, 72, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, L.; De Lara, F.C.; Martín De Las Mulas, J.; Gómez-Villamandos, J.C.; Hervás, J.; Wilkinson, P.J.; Sierra, M.A. Virus association with lymphocytes in acute African swine fever. Vet. Res. 1996, 27, 305–312. [Google Scholar] [PubMed]
- Gómez-Villamandos, J.C.; Bautista, M.J.; Sánchez-Cordón, P.J.; Carrasco, L. Pathology of African swine fever: The role of monocyte-macrophage. Virus Res. 2013, 173, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Ballester, M.; Galindo-Cardiel, I.; Gallardo, C.; Argilaguet, J.M.; Segalés, J.; Rodríguez, J.M.; Rodríguez, F. Intranuclear detection of African swine fever virus DNA in several cell types from formalin-fixed and paraffin-embedded tissues using a new in situ hybridisation protocol. J. Virol. Methods 2010, 168, 38–43. [Google Scholar] [CrossRef]
- Lv, C.; Yang, J.; Zhao, L.; Wu, C.; Kang, C.; Zhang, Q.; Sun, X.; Chen, X.; Zou, Z.; Jin, M. Infection Characteristics and Transcriptomics of African Swine Fever Virus in Bama Minipigs. Microbiol. Spectr. 2022, 10, e0383422. [Google Scholar] [CrossRef]
- Sun, E.; Huang, L.; Zhang, X.; Zhang, J.; Shen, D.; Zhang, Z.; Wang, Z.; Huo, H.; Wang, W.; Huangfu, H.; et al. Genotype I African swine fever viruses emerged in domestic pigs in China and caused chronic infection. Emerg. Microbes Infect. 2021, 10, 2183–2193. [Google Scholar] [CrossRef]
- Gómez-Villamandos, J.C.; Hervás, J.; Méndez, A.; Carrasco, L.; Martín De Las Mulas, J.; Villeda, C.J.; Wilkinson, P.J.; Sierra, M.A. Experimental African swine fever: Apoptosis of lymphocytes and virus replication in other cells. J. Gen. Virol. 1995, 76 Pt 9, 2399–2405. [Google Scholar] [CrossRef]
- Fernández De Marco, M.; Salguero, F.J.; Bautista, M.J.; Núñez, A.; Sánchez-Cordón, P.J.; Gómez-Villamandos, J.C. An immunohistochemical study of the tonsils in pigs with acute African swine fever virus infection. Res. Vet. Sci. 2007, 83, 198–203. [Google Scholar] [CrossRef]
- Howey, E.B.; O’donnell, V.; De Carvalho Ferreira, H.C.; Borca, M.V.; Arzt, J. Pathogenesis of highly virulent African swine fever virus in domestic pigs exposed via intraoropharyngeal, intranasopharyngeal, and intramuscular inoculation, and by direct contact with infected pigs. Virus Res. 2013, 178, 328–339. [Google Scholar] [CrossRef]
- Wardley, R.C.; De MAndrade, C.; Black, D.N.; De Castro Portugal, F.L.; Enjuanes, L.; Hess, W.R.; Mebus, C.; Ordas, A.; Rutili, D.; Sanchez Vizcaino, J.; et al. African Swine Fever virus. Brief review. Arch. Virol. 1983, 76, 73–90. [Google Scholar] [CrossRef]
- Sánchez-Vizcaíno, J.M.; Mur, L.; Martínez-López, B. African swine fever: An epidemiological update. Transbound. Emerg. Dis. 2012, 59 (Suppl. S1), 27–35. [Google Scholar] [CrossRef]
- Zhang, C.; Cheng, T.; Li, D.; Yu, X.; Chen, F.; He, Q. Low-host double MDA workflow for uncultured ASFV positive blood and serum sample sequencing. Front. Vet. Sci. 2022, 9, 936781. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, A.L.; Santarén, J.F.; Viñuela, E. Production and titration of African swine fever virus in porcine alveolar macrophages. J. Virol. Methods 1982, 3, 303–310. [Google Scholar] [CrossRef]
- King, D.P.; Reid, S.M.; Hutchings, G.H.; Grierson, S.S.; Wilkinson, P.J.; Dixon, L.K.; Bastos, A.D.S.; Drew, T.W. Development of a TaqMan PCR assay with internal amplification control for the detection of African swine fever virus. J. Virol. Methods 2003, 107, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Malmquist, W.A.; Hay, D. Hemadsorption and cytopathic effect produced by African Swine Fever virus in swine bone marrow and buffy coat cultures. Am. J. Vet. Res. 1960, 21, 104–108. [Google Scholar] [PubMed]
- Yu, X.; Zhu, X.; Chen, X.; Li, D.; Xu, Q.; Yao, L.; Sun, Q.; Ghonaim, A.H.; Ku, X.; Fan, S.; et al. Establishment of a Blocking ELISA Detection Method for Against African Swine Fever Virus p30 Antibody. Front. Vet. Sci. 2021, 8, 781373. [Google Scholar] [CrossRef]
- Carrascosa, A.L.; Bustos, M.J.; De Leon, P. Methods for growing and titrating African swine fever virus: Field and laboratory samples. Curr. Protoc. Cell Biol. 2011, 53, 26.14.1–26.14.25. [Google Scholar] [CrossRef]
- King, K.; Chapman, D.; Argilaguet, J.M.; Fishbourne, E.; Hutet, E.; Cariolet, R.; Hutchings, G.; Oura, C.A.L.; Netherton, C.L.; Moffat, K.; et al. Protection of European Domestic Pigs from Virulent African Isolates of African Swine Fever Virus by Experimental Immunisation. Vaccine 2011, 29, 4593–4600. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Floyd, T.; Hicks, D.; Crooke, H.R.; Mccleary, S.; Mccarthy, R.R.; Strong, R.; Dixon, L.K.; Neimanis, A.; Wikström-Lassa, E.; et al. Evaluation of Lesions and Viral Antigen Distribution in Domestic Pigs Inoculated Intranasally with African Swine Fever Virus Ken05/Tk1 (Genotype X). Pathogens 2021, 10, 768. [Google Scholar] [CrossRef]
- Plowright, W.; Parker, J.; Staple, R.F. The growth of a virulent strain of African swine fever virus in domestic pigs. J. Hyg. 1968, 66, 117–134. [Google Scholar] [CrossRef]
- Greig, A. Pathogenesis of African swine fever in pigs naturally exposed to the disease. J. Comp. Pathol. 1972, 82, 73–79. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Earliest Appearance of Disease Signs (dpi) | Pig No. in Group IM | Pig No. in Group ON | ||||
|---|---|---|---|---|---|---|
| IM-1 | IM-2 | IM-3 | ON-1 | ON-2 | ON-3 | |
| Fever (≥40 °C) | 5 | 4 | 4 | 6 | 7 | 5 |
| Inappetence | 4 | 3 | 3 | 5 | 5 | 4 |
| Lethargy | 5 | 3 | 3 | 4 | 5 | 5 |
| Wheezing/coughing | / a | / | / | 7 | / | / |
| Nosebleed | / | / | / | / | 9 | / |
| Papule | 4 | / | 4 | / | / | / |
| Cutaneous necrosis | / | / | 5 | 8 | / | / |
| Diarrhea | 6 | 4 | 4 | 4 | / | / |
| Archorrhagia | / | / | / | / | / | 8 |
| Death | 7 | 6 | 7 | NA b | 10 | 8 |
| Group | Incubation period (±SD) | Oral Shedding | Nasal Shedding | Rectal Shedding | Viremia | |||
|---|---|---|---|---|---|---|---|---|
| Days to the Onset (±SD) | Maximum Titer (±SD) a | Days to the Onset (±SD) | Maximum Titer (±SD) a | Days to the Onset (±SD) | Maximum Titer (±SD) a | Maximum Titer (±SD) a | ||
| IM | 4.33 (±0.47) b | 3.00 (±0.82) c | 6.36 (±0.20) d | 2.33 (±1.25) b | 7.20 (±0.50) b | 3.33 (±0.47) c | 6.80 (±0.17) b | 9.36 (±0.19) d |
| ON | 6.00 (±0.82) | 5.00 (±0) | 5.19 (±0.25) | 4.33 (±1.25) | 6.75 (±1.46) | 6.33 (±1.25) | 5.84 (±1.20) | 7.13 (±0.48) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Li, D.; Chen, X.; Liu, X.; Cao, H.; Wang, H.; Wu, H.; Cheng, T.; Ren, W.; Xu, F.; et al. In Vivo Study of Inoculation Approaches and Pathogenicity in African Swine Fever. Vet. Sci. 2024, 11, 403. https://doi.org/10.3390/vetsci11090403
Xu Q, Li D, Chen X, Liu X, Cao H, Wang H, Wu H, Cheng T, Ren W, Xu F, et al. In Vivo Study of Inoculation Approaches and Pathogenicity in African Swine Fever. Veterinary Sciences. 2024; 11(9):403. https://doi.org/10.3390/vetsci11090403
Chicago/Turabian StyleXu, Qian, Dongfan Li, Xiaoyu Chen, Xiaoli Liu, Hua Cao, Hui Wang, Haowei Wu, Tangyu Cheng, Wenhui Ren, Fengqin Xu, and et al. 2024. "In Vivo Study of Inoculation Approaches and Pathogenicity in African Swine Fever" Veterinary Sciences 11, no. 9: 403. https://doi.org/10.3390/vetsci11090403
APA StyleXu, Q., Li, D., Chen, X., Liu, X., Cao, H., Wang, H., Wu, H., Cheng, T., Ren, W., Xu, F., He, Q., Yu, X., & Li, W. (2024). In Vivo Study of Inoculation Approaches and Pathogenicity in African Swine Fever. Veterinary Sciences, 11(9), 403. https://doi.org/10.3390/vetsci11090403