
Influence of Maternal Nutrition and One-Carbon Metabolites Supplementation during Early Pregnancy on Bovine Fetal Small Intestine Vascularity and Cell Proliferation

 ,
,  ,
,  , , , , and
, , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Animals, Diet, and Treatment
2.3. Sample Collection and Preparation
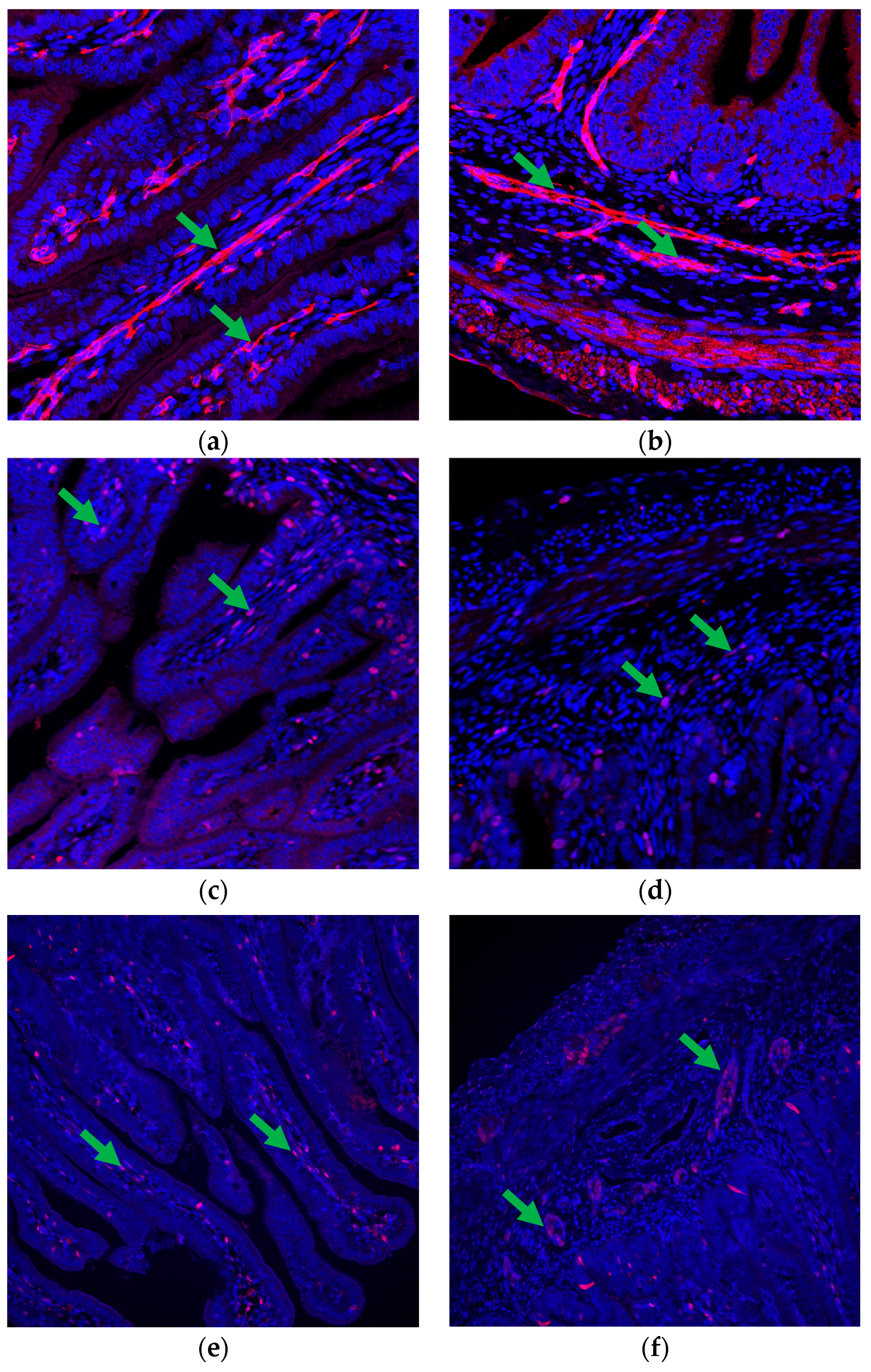
2.4. Immunohistochemistry and Immunofluorescence
2.5. Image Processing
2.6. Statistical Analysis
3. Results
3.1. Small Intestine Phenotype
3.2. Capillary Development of the Fetal Small Intestine
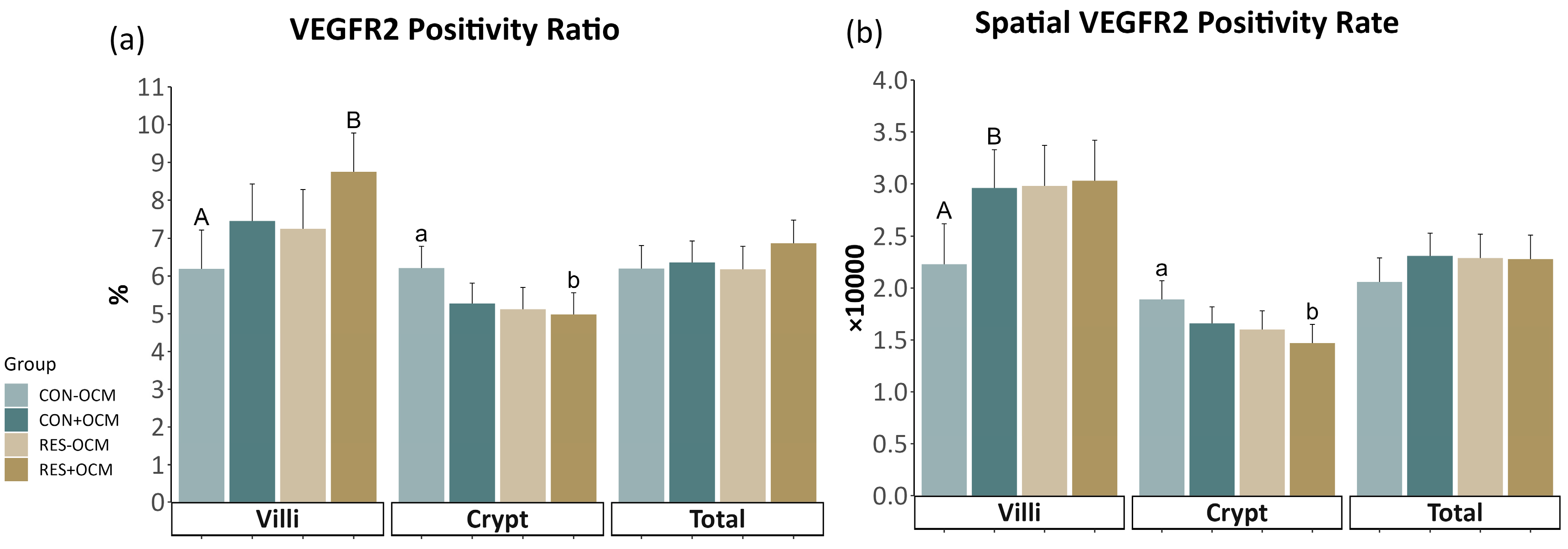
3.3. VEGFR2 Expression in the Fetal Small Intestine
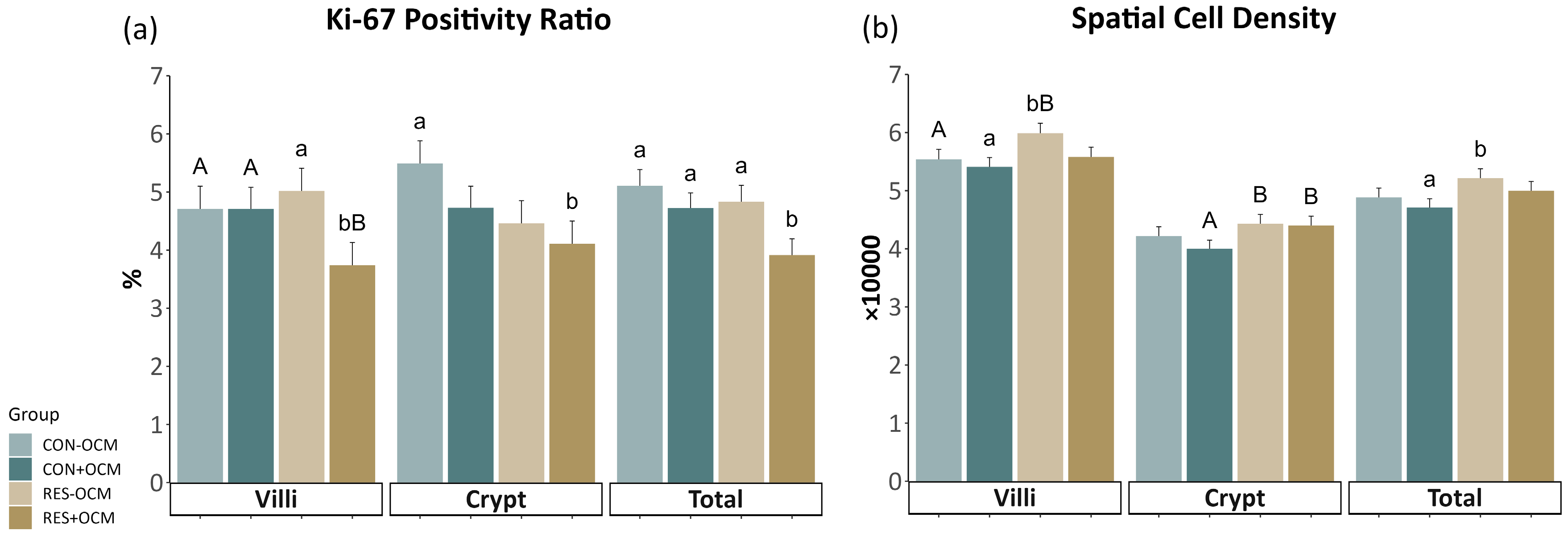
3.4. Cell Proliferation of the Fetal Small Intestine
3.5. Correlations between Variables
4. Discussion
4.1. Fetal Small Intestine Weight
4.2. Vascular Development in the Fetal Intestine
4.3. Cell Proliferation in the Fetal Intestine
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caton, J.S.; Crouse, M.S.; McLean, K.J.; Dahlen, C.R.; Ward, A.K.; Cushman, R.A.; Grazul-Bilska, A.T.; Neville, B.W.; Borowicz, P.P.; Reynolds, L.P. Maternal periconceptual nutrition, early pregnancy, and developmental outcomes in beef cattle. J. Anim. Sci. 2020, 98, skaa358. [Google Scholar] [CrossRef]
- Hammer, C.J.; Caton, J.S.; Dahlen, C.R.; Ward, A.K.; Borowicz, P.P.; Reynolds, L.P. DOHaD: A MENAGERIE OF ADAPTATIONS AND PERSPECTIVES: Large animal models of developmental programming: Sustenance, stress, and sex matter. Reproduction 2023, 165, F1–F13. [Google Scholar] [CrossRef] [PubMed]
- Funston, R.N.; Larson, D.M.; Vonnahme, K.A. Effects of maternal nutrition on conceptus growth and offspring performance: Implications for beef cattle production. J. Anim. Sci. 2010, 88, E205–E215. [Google Scholar] [CrossRef] [PubMed]
- Anas, M.; Diniz, W.J.S.; Menezes, A.C.B.; Reynolds, L.P.; Caton, J.S.; Dahlen, C.R.; Ward, A.K. Maternal Mineral Nutrition Regulates Fetal Genomic Programming in Cattle: A Review. Metabolites 2023, 13, 593. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.M.; Reed, J.J.; Vonnahme, K.A.; Soto-Navarro, S.A.; Reynolds, L.P.; Ford, S.P.; Hess, B.W.; Caton, J.S. Effects of stage of gestation and nutrient restriction during early to mid-gestation on maternal and fetal visceral organ mass and indices of jejunal growth and vascularity in beef cows. J. Anim. Sci. 2010, 88, 2410–2424. [Google Scholar] [CrossRef] [PubMed]
- Prezotto, L.D.; Lemley, C.O.; Camacho, L.E.; Doscher, F.E.; Meyer, A.M.; Caton, J.S.; Awda, B.J.; Vonnahme, K.A.; Swanson, K.C. Effects of nutrient restriction and melatonin supplementation on maternal and foetal hepatic and small intestinal energy utilization. J. Anim. Physiol. Anim. Nutr. 2014, 98, 797–807. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.L.; AC, B.M.; Kassetas, C.J.; Baumgaertner, F.; Kirsch, J.D.; Dorsam, S.T.; Neville, T.L.; Ward, A.K.; Borowicz, P.P.; Reynolds, L.P.; et al. Vitamin and Mineral Supplementation and Rate of Gain in Beef Heifers II: Effects on Concentration of Trace Minerals in Maternal Liver and Fetal Liver, Muscle, Allantoic, and Amniotic Fluids at Day 83 of Gestation. Animals 2022, 12, 1925. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.M.; Hess, B.W.; Paisley, S.I.; Du, M.; Caton, J.S. Small intestinal growth measures are correlated with feed efficiency in market weight cattle, despite minimal effects of maternal nutrition during early to midgestation. J. Anim. Sci. 2014, 92, 3855–3867. [Google Scholar] [CrossRef]
- Yunusova, R.D.; Neville, T.L.; Vonnahme, K.A.; Hammer, C.J.; Reed, J.J.; Taylor, J.B.; Redmer, D.A.; Reynolds, L.P.; Caton, J.S. Impacts of maternal selenium supply and nutritional plane on visceral tissues and intestinal biology in 180-day-old offspring in sheep. J. Anim. Sci. 2013, 91, 2229–2242. [Google Scholar] [CrossRef]
- Trahair, J.F.; DeBarro, T.M.; Robinson, J.S.; Owens, J.A. Restriction of nutrition in utero selectively inhibits gastrointestinal growth in fetal sheep. J. Nutr. 1997, 127, 637–641. [Google Scholar] [CrossRef]
- Reed, J.J.; Ward, M.A.; Vonnahme, K.A.; Neville, T.L.; Julius, S.L.; Borowicz, P.P.; Taylor, J.B.; Redmer, D.A.; Grazul-Bilska, A.T.; Reynolds, L.P.; et al. Effects of selenium supply and dietary restriction on maternal and fetal body weight, visceral organ mass and cellularity estimates, and jejunal vascularity in pregnant ewe lambs. J. Anim. Sci. 2007, 85, 2721–2733. [Google Scholar] [CrossRef]
- Neville, T.L.; Redmer, D.A.; Borowicz, P.P.; Reed, J.J.; Ward, M.A.; Johnson, M.L.; Taylor, J.B.; Soto-Navarro, S.A.; Vonnahme, K.A.; Reynolds, L.P.; et al. Maternal dietary restriction and selenium supply alters messenger ribonucleic acid expression of angiogenic factors in maternal intestine, mammary gland, and fetal jejunal tissues during late gestation in pregnant ewe lambs. J. Anim. Sci. 2010, 88, 2692–2702. [Google Scholar] [CrossRef]
- Meyer, A.M.; Neville, T.L.; Reed, J.J.; Taylor, J.B.; Reynolds, L.P.; Redmer, D.A.; Hammer, C.J.; Vonnahme, K.A.; Caton, J.S. Maternal nutritional plane and selenium supply during gestation impact visceral organ mass and intestinal growth and vascularity of neonatal lamb offspring. J. Anim. Sci. 2013, 91, 2628–2639. [Google Scholar] [CrossRef]
- Meyer, A.M.; Caton, J.S. Role of the Small Intestine in Developmental Programming: Impact of Maternal Nutrition on the Dam and Offspring. Adv. Nutr. 2016, 7, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Caton, J.S.; Crouse, M.S.; Reynolds, L.P.; Neville, T.L.; Dahlen, C.R.; Ward, A.K.; Swanson, K.C. Maternal nutrition and programming of offspring energy requirements. Transl. Anim. Sci. 2019, 3, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Borowicz, P.P.; Caton, J.S.; Crouse, M.S.; Dahlen, C.R.; Ward, A.K. Developmental Programming of Fetal Growth and Development. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 229–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lan, X.; Radunz, A.; Khatib, H. Maternal nutrition during pregnancy is associated with differential expression of imprinted genes and DNA methyltranfereases in muscle of beef cattle offspring. J. Anim. Sci. 2015, 93, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Chmurzynska, A. Fetal programming: Link between early nutrition, DNA methylation, and complex diseases. Nutr. Rev. 2010, 68, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. The supply of choline is important for fetal progenitor cells. Semin. Cell Dev. Biol. 2011, 22, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Clare, C.E.; Brassington, A.H.; Kwong, W.Y.; Sinclair, K.D. One-Carbon Metabolism: Linking Nutritional Biochemistry to Epigenetic Programming of Long-Term Development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef]
- Sinclair, K.D.; Allegrucci, C.; Singh, R.; Gardner, D.S.; Sebastian, S.; Bispham, J.; Thurston, A.; Huntley, J.F.; Rees, W.D.; Maloney, C.A.; et al. DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc. Natl. Acad. Sci. USA 2007, 104, 19351–19356. [Google Scholar] [CrossRef]
- Kasture, V.V.; Sundrani, D.P.; Joshi, S.R. Maternal one carbon metabolism through increased oxidative stress and disturbed angiogenesis can influence placental apoptosis in preeclampsia. Life Sci. 2018, 206, 61–69. [Google Scholar] [CrossRef]
- Crouse, M.S.; Greseth, N.P.; McLean, K.J.; Crosswhite, M.R.; Pereira, N.N.; Ward, A.K.; Reynolds, L.P.; Dahlen, C.R.; Neville, B.W.; Borowicz, P.P.; et al. Maternal nutrition and stage of early pregnancy in beef heifers: Impacts on hexose and AA concentrations in maternal and fetal fluids1. J. Anim. Sci. 2019, 97, 1296–1316. [Google Scholar] [CrossRef]
- Crouse, M.S.; Caton, J.S.; Cushman, R.A.; McLean, K.J.; Dahlen, C.R.; Borowicz, P.P.; Reynolds, L.P.; Ward, A.K. Moderate nutrient restriction of beef heifers alters expression of genes associated with tissue metabolism, accretion, and function in fetal liver, muscle, and cerebrum by day 50 of gestation. Transl. Anim. Sci. 2019, 3, 855–866. [Google Scholar] [CrossRef]
- Lamb, G.C.; Dahlen, C.R.; Larson, J.E.; Marquezini, G.; Stevenson, J.S. Control of the estrous cycle to improve fertility for fixed-time artificial insemination in beef cattle: A review. J. Anim. Sci. 2010, 88, E181–E192. [Google Scholar] [CrossRef] [PubMed]
- Gionbelli, T.R.S.; Rotta, P.P.; Veloso, C.M.; Valadares Filho, S.C.; Carvalho, B.C.; Marcondes, M.I.; Ferreira, M.F.L.; Souza, J.V.F.; Santos, J.; Lacerda, L.C.; et al. Intestinal development of bovine foetuses during gestation is affected by foetal sex and maternal nutrition. J. Anim. Physiol. Anim. Nutr. 2017, 101, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.E.; Brozek, J.M.; Keen-Rhinehart, E. Our stolen figures: The interface of sexual differentiation, endocrine disruptors, maternal programming, and energy balance. Horm. Behav. 2014, 66, 104–119. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine; Committee on Nutrient Requirements of Beef Cattle. Nutrient Requirements of Beef Cattle, 8th ed.; National Academies Press: Washington, DC, USA, 2016; Chapter xvii; p. 475. [Google Scholar]
- Crouse, M.S.; Freetly, H.C.; Lindholm-Perry, A.K.; Neville, B.W.; Oliver, W.T.; Lee, R.T.; Syring, J.G.; King, L.E.; Reynolds, L.P.; Dahlen, C.R.; et al. One-carbon metabolite supplementation to heifers for the first 14 d of the estrous cycle alters the plasma and hepatic one-carbon metabolite pool and methionine-folate cycle enzyme transcript abundance in a dose-dependent manner. J. Anim. Sci. 2023, 101, skac419. [Google Scholar] [CrossRef] [PubMed]
- Jacometo, C.B.; Zhou, Z.; Luchini, D.; Correa, M.N.; Loor, J.J. Maternal supplementation with rumen-protected methionine increases prepartal plasma methionine concentration and alters hepatic mRNA abundance of 1-carbon, methionine, and transsulfuration pathways in neonatal Holstein calves. J. Dairy. Sci. 2017, 100, 3209–3219. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, A.; Khan, D.R.; Sirard, M.A.; Girard, C.L.; Laforest, J.P.; Richard, F.J. Effects of intramuscular administration of folic acid and vitamin B12 on granulosa cells gene expression in postpartum dairy cows. J. Dairy. Sci. 2015, 98, 7797–7809. [Google Scholar] [CrossRef] [PubMed]
- Lamb, G.; Dahlen, C.; Brown, D. Reproductive ultrasonography for monitoring ovarian structure development, fetal development, embryo survival, and twins in beef cows. Prof. Anim. Sci. 2003, 19, 135–143. [Google Scholar] [CrossRef]
- Trotta, R.J.; Vasquez-Hidalgo, M.A.; Vonnahme, K.A.; Swanson, K.C. Effects of Nutrient Restriction During Midgestation to Late Gestation on Maternal and Fetal Postruminal Carbohydrase Activities in Sheep. J. Anim. Sci. 2020, 98, skz393. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.; Gionbelli, M.; Paulino, P.; Serão, N.; Martins, T.; Tótaro, P.; Neves, C.; Valadares Filho, S.; Dodson, M.; Zhu, M. Effects of maternal nutrition on development of gastrointestinal tract of bovine fetus at different stages of gestation. Livest. Sci. 2013, 153, 60–65. [Google Scholar] [CrossRef]
- Zhang, Y.; Otomaru, K.; Oshima, K.; Goto, Y.; Oshima, I.; Muroya, S.; Sano, M.; Saneshima, R.; Nagao, Y.; Kinoshita, A.; et al. Effects of low and high levels of maternal nutrition consumed for the entirety of gestation on the development of muscle, adipose tissue, bone, and the organs of Wagyu cattle fetuses. Anim. Sci. J. 2021, 92, e13600. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.L.; Willard, D.E. Natural selection of parental ability to vary the sex ratio of offspring. Science 1973, 179, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Crouse, M.S.; Caton, J.S.; Menezes, A.C.B.; Reynolds, L.P.; Dahlen, C.R.; Borowicz, P.P.; Ward, A.K. One-carbon metabolite supplementation improves growth of bovine embryonic fibroblasts cultured in divergent glucose media. Transl. Anim. Sci. 2019, 3, 1705–1709. [Google Scholar] [CrossRef] [PubMed]
- Crouse, M.; Caton, J.; Claycombe-Larson, K.; Dahlen, C.; Reynolds, L.; Borowicz, P.; Ward, A. Effects of glucose and one-carbon metabolites on mitochondrial respiration in bovine embryonic cells. In EAAP Scientific Series; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 237–238. [Google Scholar]
- Syring, J.G.; Crouse, M.S.; Neville, T.L.; Ward, A.K.; Dahlen, C.R.; Reynolds, L.P.; Borowicz, P.P.; McLean, K.J.; Neville, B.W.; Caton, J.S. Concentrations of vitamin B12 and folate in maternal serum and fetal fluids, metabolite interrelationships, and hepatic transcript abundance of key folate and methionine cycle genes: The impacts of maternal nutrition during the first 50 d of gestation. J. Anim. Sci. 2023, 101, skad139. [Google Scholar] [CrossRef] [PubMed]
- Oosterbaan, A.M.; Steegers, E.A.; Ursem, N.T. The effects of homocysteine and folic acid on angiogenesis and VEGF expression during chicken vascular development. Microvasc. Res. 2012, 83, 98–104. [Google Scholar] [CrossRef]
- Wang, X.J.; Tian, D.C.; Wang, F.W.; Zhang, M.H.; Fan, C.D.; Chen, W.; Wang, M.H.; Fu, X.Y.; Ma, J.K. Astaxanthin inhibits homocysteine-induced endothelial cell dysfunction via the regulation of the reactive oxygen species-dependent VEGF-VEGFR2-FAK signaling pathway. Mol. Med. Rep. 2019, 19, 4753–4760. [Google Scholar] [CrossRef]
- Xu, X.; Yang, X.Y.; He, B.W.; Yang, W.J.; Cheng, W.W. Placental NRP1 and VEGF expression in pre-eclamptic women and in a homocysteine-treated mouse model of pre-eclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 196, 69–75. [Google Scholar] [CrossRef]
- Omorphos, N.P.; Gao, C.; Tan, S.S.; Sangha, M.S. Understanding angiogenesis and the role of angiogenic growth factors in the vascularisation of engineered tissues. Mol. Biol. Rep. 2021, 48, 941–950. [Google Scholar] [CrossRef]
- Jiang, Q.; Sherlock, D.N.; Zhang, H.; Guyader, J.; Pan, Y.X.; Loor, J.J. One-carbon metabolism and related pathways in ruminal and small intestinal epithelium of lactating dairy cows. J. Anim. Sci. 2023, 101, skad062. [Google Scholar] [CrossRef]
- Holmes, K.; Charnock Jones, S.D.; Forhead, A.J.; Giussani, D.A.; Fowden, A.L.; Licence, D.; Kempster, S.; Smith, G.C. Localization and control of expression of VEGF-A and the VEGFR-2 receptor in fetal sheep intestines. Pediatr. Res. 2008, 63, 143–148. [Google Scholar] [CrossRef]
- Neville, T.; Reed, J.; Vonnahme, K.; Borowicz, P.; Taylor, J.; Redmer, D.; Luther, J.; Hammer, C.; Reynolds, L.; Caton, J. Effects of maternal nutrition and selenium supply on jejunal characteristics and mRNA expression of angiogenic factors and receptors in offspring at harvest. In Proceedings of the Western Section, American Society of Animal Science, Laramie, WY, USA, 24–28 June 2008; pp. 318–321. [Google Scholar]
- Seyyedin, S.; Nazem, M.N. Histomorphometric study of the effect of methionine on small intestine parameters in rat: An applied histologic study. Folia Morphol. 2017, 76, 620–629. [Google Scholar] [CrossRef]
- Miousse, I.R.; Pathak, R.; Garg, S.; Skinner, C.M.; Melnyk, S.; Pavliv, O.; Hendrickson, H.; Landes, R.D.; Lumen, A.; Tackett, A.J.; et al. Short-term dietary methionine supplementation affects one-carbon metabolism and DNA methylation in the mouse gut and leads to altered microbiome profiles, barrier function, gene expression and histomorphology. Genes. Nutr. 2017, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Bressenot, A.; Pooya, S.; Bossenmeyer-Pourie, C.; Gauchotte, G.; Germain, A.; Chevaux, J.B.; Coste, F.; Vignaud, J.M.; Gueant, J.L.; Peyrin-Biroulet, L. Methyl donor deficiency affects small-intestinal differentiation and barrier function in rats. Br. J. Nutr. 2013, 109, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Van Nevel, C.J.; Decuypere, J.A.; Dierick, N.A.; Molly, K. Incorporation of galactomannans in the diet of newly weaned piglets: Effect on bacteriological and some morphological characteristics of the small intestine. Arch. Anim. Nutr. 2005, 59, 123–138. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments 2 | p-Value 3 | |||||||
|---|---|---|---|---|---|---|---|---|
| Traits | CON − OCM | CON + OCM | RES − OCM | RES + OCM | SEM 4 | MFI | OCM | MFI × OCM |
| SIW (g) 5 | 74.14 | 65.0 | 68.01 | 63.11 | 2.29 | 0.17 | 0.02 | 0.47 |
| Relative SIW (g/kg of body weight) | 1.89 | 1.77 | 1.77 | 1.59 | 0.08 | 0.08 | 0.08 | 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daneshi, M.; Borowicz, P.P.; Entzie, Y.L.; Syring, J.G.; King, L.E.; Safain, K.S.; Anas, M.; Reynolds, L.P.; Ward, A.K.; Dahlen, C.R.; et al. Influence of Maternal Nutrition and One-Carbon Metabolites Supplementation during Early Pregnancy on Bovine Fetal Small Intestine Vascularity and Cell Proliferation. Vet. Sci. 2024, 11, 146. https://doi.org/10.3390/vetsci11040146
Daneshi M, Borowicz PP, Entzie YL, Syring JG, King LE, Safain KS, Anas M, Reynolds LP, Ward AK, Dahlen CR, et al. Influence of Maternal Nutrition and One-Carbon Metabolites Supplementation during Early Pregnancy on Bovine Fetal Small Intestine Vascularity and Cell Proliferation. Veterinary Sciences. 2024; 11(4):146. https://doi.org/10.3390/vetsci11040146
Chicago/Turabian StyleDaneshi, Mojtaba, Pawel P. Borowicz, Yssi L. Entzie, Jessica G. Syring, Layla E. King, Kazi Sarjana Safain, Muhammad Anas, Lawrence P. Reynolds, Alison K. Ward, Carl R. Dahlen, and et al. 2024. "Influence of Maternal Nutrition and One-Carbon Metabolites Supplementation during Early Pregnancy on Bovine Fetal Small Intestine Vascularity and Cell Proliferation" Veterinary Sciences 11, no. 4: 146. https://doi.org/10.3390/vetsci11040146
APA StyleDaneshi, M., Borowicz, P. P., Entzie, Y. L., Syring, J. G., King, L. E., Safain, K. S., Anas, M., Reynolds, L. P., Ward, A. K., Dahlen, C. R., Crouse, M. S., & Caton, J. S. (2024). Influence of Maternal Nutrition and One-Carbon Metabolites Supplementation during Early Pregnancy on Bovine Fetal Small Intestine Vascularity and Cell Proliferation. Veterinary Sciences, 11(4), 146. https://doi.org/10.3390/vetsci11040146