
The Adaptation Time to the Extender as a Crucial Step for an Accurate Evaluation of Ram Sperm Quality during the Liquid Storage

 ,
,  , ,
, ,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Design
2.2.1. Experiment 1: Establishment of a Stabilization Period for Ram Sperm
2.2.2. Experiment 2: Identification of the Factor Altering Ram Sperm Quality
2.3. Sperm Quality Evaluation
2.3.1. Sperm Motility and Kinetic Parameter Analysis with the CASA System
2.3.2. Sperm Functionality Analysis with Flow Cytometry
2.4. Statistical Analysis
3. Results
3.1. Experiment 1: Establishment of a Stabilization Period for Ram Sperm
3.1.1. Sperm Motility and Kinetic Parameters
3.1.2. Sperm Functionality
3.2. Experiment 2: Identification of the Factor Altering Ram Sperm Quality
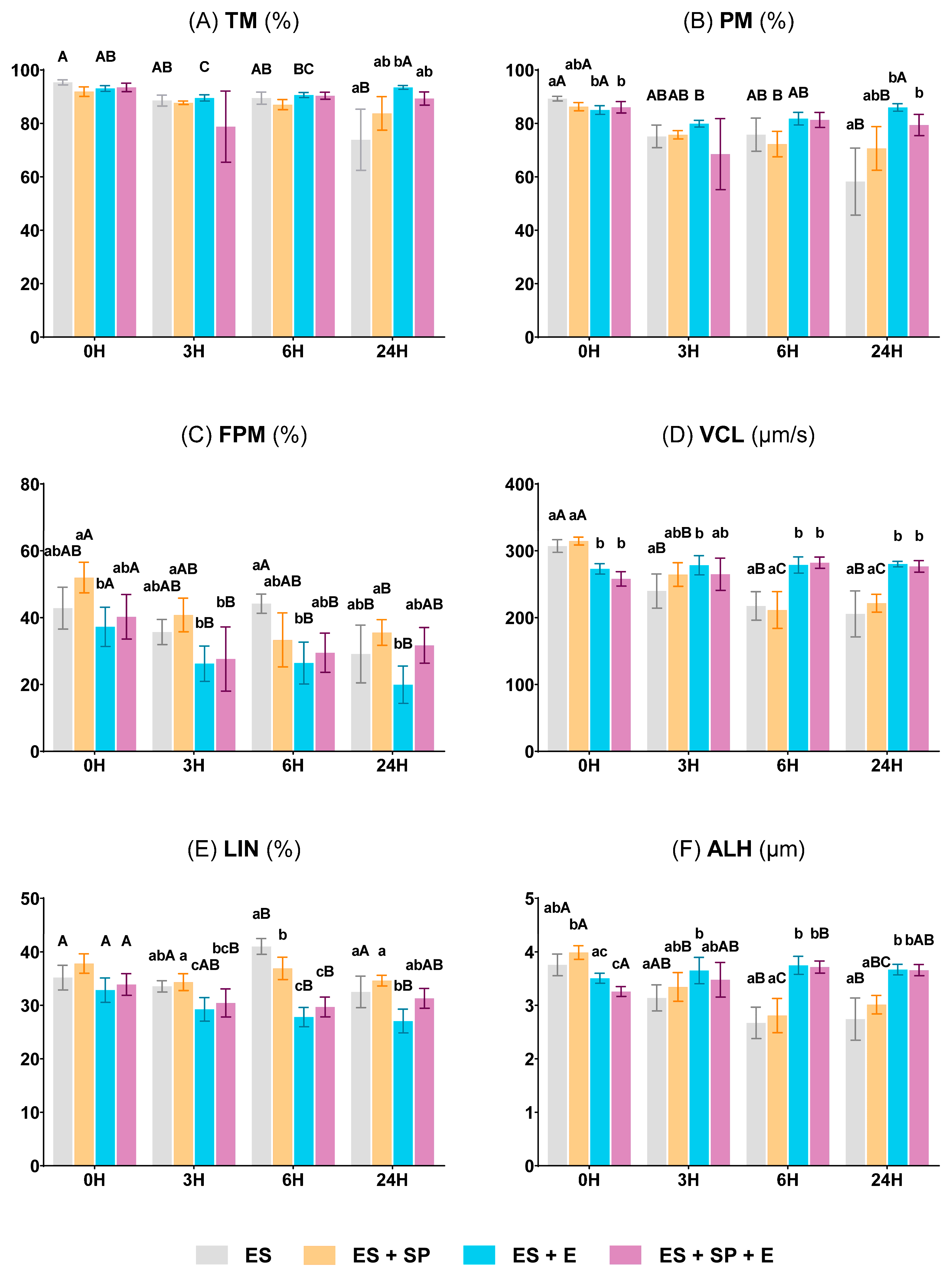
3.2.1. Sperm Motility and Kinetic Parameters
3.2.2. Sperm Functionality
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naitana, S.; Ledda, S. Reproductive Technologies in Sheep; INC: New York, NY, USA, 2020; ISBN 9780128171073. [Google Scholar]
- Amiridis, G.S.; Cseh, S. Assisted Reproductive Technologies in the Reproductive Management of Small Ruminants. Anim. Reprod. Sci. 2012, 130, 152–161. [Google Scholar] [CrossRef]
- Kameni, S.L.; Meutchieye, F.; Ngoula, F. Liquid Storage of Ram Semen: Associated Damages and Improvement. Open J. Anim. Sci. 2021, 11, 473–500. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, H. Laboratory Semen Assessment and Prediction of Fertility: Still Utopia? In Reproduction in Domestic Animals; Academic Press: Cambridge, MA, USA, 2003; Volume 38, pp. 312–318. [Google Scholar]
- Vicente-Fiel, S.; Palacín, I.; Santolaria, P.; Fantova, E.; Quintín-Casorrán, F.J.; Sevilla-Mur, E.; Yániz, J.L. In Vitro Assessment of Sperm Quality from Rams of High and Low Field Fertility. Anim. Reprod. Sci. 2014, 146, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Gillan, L.; Maxwell, W.M.C.; Evans, G. Preservation and Evaluation of Semen for Artificial Insemination. Reprod. Fertil. Dev. 2004, 16, 447–454. [Google Scholar] [CrossRef]
- Kasimanickam, R.; Kasimanickam, V.; Tibary, A.; Pelzer, K. Effect of Semen Extenders on Sperm Parameters of Ram Semen during Liquid Storage at 4 °C. Small Rumin. Res. 2011, 99, 208–213. [Google Scholar] [CrossRef]
- Quan, G.B.; Wu, G.Q.; Wang, Y.J.; Li, D.J.; Ma, Y.; Hong, Q.H. Effects of the Tris, Tes, or Skim Milk Based Extender on in vitro Parameters of Ram Spermatozoa during Liquid Storage. Small Rumin. Res. 2016, 134, 14–21. [Google Scholar] [CrossRef]
- López-Sáez, A.; Ortiz, N.; Gallego, L.; Garde, J.J. Liquid Storage (5 °C) of Ram Semen in Different Diluents. Arch. Androl. 2000, 44, 155–164. [Google Scholar] [CrossRef]
- Paulenz, H.; Söderquist, L.; Pérez-Pé, R.; Andersen Berg, K. Effect of Different Extenders and Storage Temperatures on Sperm Viability of Liquid Ram Semen. Theriogenology 2002, 57, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Anel, L.; De Paz, P.; Álvarez, M.; Chamorro, C.A.; Boixo, J.C.; Manso, A.; González, M.; Kaabi, M.; Anel, E. Field and in vitro Assay of Three Methods for Freezing Ram Semen. Theriogenology 2003, 60, 1293–1308. [Google Scholar] [CrossRef]
- Gil, J.; Lundeheim, N.; Söderquist, L.; Rodríguez-Martínez, H. Influence of Extender, Temperature, and Addition of Glycerol on Post-Thaw Sperm Parameters in Ram Semen. Theriogenology 2003, 59, 1241–1255. [Google Scholar] [CrossRef]
- D’Alessandro, A.G.; Martemucci, G.; Colonna, M.A.; Bellitti, A. Post-Thaw Survival of Ram Spermatozoa and Fertility after Insemination as Affected by Prefreezing Sperm Concentration and Extender Composition. Theriogenology 2001, 55, 1159–1170. [Google Scholar] [CrossRef]
- Gundogan, M.; Yeni, D.; Avdatek, F.; Fidan, A.F. Influence of Sperm Concentration on the Motility, Morphology, Membrane and DNA Integrity along with Oxidative Stress Parameters of Ram Sperm during Liquid Storage. Anim. Reprod. Sci. 2010, 122, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Tamayo-Canul, J.; Anel, E.; Boixo, J.C.; Mata-Campuzano, M.; Martinez-Pastor, F.; Anel, L.; de Paz, P. Sperm Concentration at Freezing Affects Post-Thaw Quality and Fertility of Ram Semen. Theriogenology 2012, 77, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Gibb, Z.; Lambourne, S.R.; Quadrelli, J.; Smith, N.D.; Aitken, R.J. L-Carnitine and Pyruvate Are Prosurvival Factors during the Storage of Stallion Spermatozoa at Room Temperature. Biol. Reprod. 2015, 93, 104. [Google Scholar] [CrossRef] [PubMed]
- Pini, T.; Leahy, T.; de Graaf, S.P. Sublethal Sperm Freezing Damage: Manifestations and Solutions. Theriogenology 2018, 118, 172–181. [Google Scholar] [CrossRef]
- Rizkallah, N.; Chambers, C.G.; de Graaf, S.P.; Rickard, J.P. Factors Affecting the Survival of Ram Spermatozoa during Liquid Storage and Options for Improvement. Animals 2022, 12, 244. [Google Scholar] [CrossRef]
- Holt, W.V.; Medrano, A.; Thurston, L.M.; Watson, P.F. The Significance of Cooling Rates and Animal Variability for Boar Sperm Cryopreservation: Insights from the Cryomicroscope. In Theriogenology; Elsevier: Amsterdam, The Netherlands, 2005; Volume 63, pp. 370–382. [Google Scholar]
- Okano, T.; Murase, T.; Asano, M.; Tsubota, T. Effects of Final Dilution Rate, Sperm Concentration and Times for Cooling and Glycerol Equilibration on Post-Thaw Characteristics of Canine Spermatozoa. J. Vet. Med. Sci. 2004, 66, 1359–1364. [Google Scholar] [CrossRef]
- Belala, R.; Briand-Amirat, L.; Vinciguerra, L.; Tainturier, D.; Kaidi, R.; Thorin, C.; Michaud, S.; Anton, M.; Bencharif, D. Effect of Equilibration Time on the Motility and Functional Integrity of Canine Spermatozoa Frozen in Three Different Extenders. Res. Vet. Sci. 2016, 106, 66–73. [Google Scholar] [CrossRef]
- Schäfer, J.; Waberski, D.; Jung, M.; Schulze, M. Impact of Holding and Equilibration Time on Post-Thaw Quality of Shipped Boar Semen. Anim. Reprod. Sci. 2017, 187, 109–115. [Google Scholar] [CrossRef]
- Passarelli, M.d.S.; Pinoti Pavaneli, A.P.; Mouro Ravagnani, G.; Pasini Martins, M.; Pedrosa, A.C.; Maria Massami Kitamura Martins, S.; Rocha, N.R.d.A.; Bittar Rigo, V.H.; Yasui, G.; Yeste, M.; et al. Effects of Different Equilibration Times at 5 °C on Boar Sperm Cryotolerance. Anim. Reprod. Sci. 2020, 219, 106547. [Google Scholar] [CrossRef]
- Yi, Y.J.; Cheon, Y.M.; Park, C.S. Effect of N-Acetyl-d-Glucosamine, and Glycerol Concentration and Equilibration Time on Acrosome Morphology and Motility of Frozen-Thawed Boar Sperm. Anim. Reprod. Sci. 2002, 69, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.M.; Eivers, B.; O’Meara, C.M.; Lonergan, P.; Fair, S. Effect of Increasing Equilibration Time of Diluted Bull Semen up to 72 h Prior to Freezing on Sperm Quality Parameters and Calving Rate Following Artificial Insemination. Theriogenology 2018, 108, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Doležalová, M.; Stádník, L.; Biniová, Z.; Ducháček, J.; Stupka, R. Equilibration and Freezing Interactions Affecting Bull Sperm Characteristics after Thawing. Czech J. Anim. Sci. 2016, 61, 515–525. [Google Scholar] [CrossRef]
- Leite, T.G.; do Vale Filho, V.R.; de Arruda, R.P.; de Andrade, A.F.C.; Emerick, L.L.; Zaffalon, F.G.; Martins, J.A.M.; Andrade, V.J.d. Effects of Extender and Equilibration Time on Post-Thaw Motility and Membrane Integrity of Cryopreserved Gyr Bull Semen Evaluated by CASA and Flow Cytometry. Anim. Reprod. Sci. 2010, 120, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Melean, M.; Ibanescu, I.; Pieper, L.; Goebel, H.; Leiding, C.; Bollwein, H. Impact of a Prolonged Equilibration Period Prior to Freezing on Sperm Quality and Fertility of AI Bulls. Anim. Reprod. Sci. 2022, 247, 107133. [Google Scholar] [CrossRef]
- Ahmad, M.; Nasrullah, R.; Ahmad, N. Effect of Cooling Rate and Equilibration Time on Pre-Freeze and Post-Thaw Survival of Buck Sperm. Cryobiology 2015, 70, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Vozaf, J.; Makarevich, A.V.; Balazi, A.; Vasicek, J.; Svoradova, A.; Olexikova, L.; Chrenek, P. Cryopreservation of Ram Semen: Manual versus Programmable Freezing and Different Lengths of Equilibration. Anim. Sci. J. 2021, 92, e13670. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.K.; Balaganur, K.; Kumar, D.; Singh, R. Pre-Freezing Equilibration for 22 h Improves Post-Thaw Sperm Functions in Cryopreserved Ram Semen by Reducing Cholesterol Efflux. Cryobiology 2020, 96, 76–84. [Google Scholar] [CrossRef]
- Câmara, D.R.; Silva, S.V.; Almeida, F.C.; Nunes, J.F.; Guerra, M.M.P. Effects of Antioxidants and Duration of Pre-Freezing Equilibration on Frozen-Thawed Ram Semen. Theriogenology 2011, 76, 342–350. [Google Scholar] [CrossRef]
- Purdy, P.H. The Post-Thaw Quality of Ram Sperm Held for 0 to 48 h at 5 °C Prior to Cryopreservation. Anim. Reprod. Sci. 2006, 93, 114–123. [Google Scholar] [CrossRef]
- Purdy, P.H.; Mocé, E.; Stobart, R.; Murdoch, W.J.; Moss, G.E.; Larson, B.; Ramsey, S.; Graham, J.K.; Blackburn, H.D. The Fertility of Ram Sperm Held for 24 h at 5 °C Prior to Cryopreservation. Anim. Reprod. Sci. 2010, 118, 231–235. [Google Scholar] [CrossRef]
- Tibary, A.; Manar, S. Cryo-Preservation of Sperm and Embryos in Small Ruminants. Rev. Marocaine Des Sci. Agron. Vétérinaires 2018, 6, 195–210. [Google Scholar]
- Lv, C.; Wu, G.; Hong, Q.; Quan, G. Spermatozoa Cryopreservation: State of Art and Future in Small Ruminants. Biopreserv. Biobank. 2019, 17, 171–182. [Google Scholar] [CrossRef]
- Batellier, F.; Vidament, M.; Fauquant, J.; Duchamp, G.; Arnaud, G.; Yvon, J.M.; Magistrini, M. Advances in Cooled Semen Technology. Anim. Reprod. Sci. 2001, 68, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Barrier-Battut, I.; Bonnet, C.; Giraudo, A.; Dubois, C.; Caillaud, M.; Vidament, M. Removal of Seminal Plasma Enhances Membrane Stability on Fresh and Cooled Stallion Spermatozoa. Reprod. Domest. Anim. 2013, 48, 64–71. [Google Scholar] [CrossRef]
- Love, C.C.; Brinsko, S.P.; Rigby, S.L.; Thompson, J.A.; Blanchard, T.L.; Varner, D.D. Relationship of Seminal Plasma Level and Extender Type to Sperm Motility and DNA Integrity. Theriogenology 2005, 63, 1584–1591. [Google Scholar] [CrossRef]
- Brinsko, S.P.; Crockett, E.C.; Squires, E.L. Effect of Centrifugation and Partial Removal of Seminal Plasma on Equine Spermatozoal Motility after Cooling and Storage. Theriogenology 2000, 54, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Bedford, S.J.; Graham, J.K.; Amann, R.P.; Squires, E.L.; Pickett, B.W. Use of Two Freezing Extenders to Cool Stallion Spermatozoa to 5 °C with and without Seminal Plasma. Theriogenology 1995, 43, 939–953. [Google Scholar] [CrossRef]
- Pavaneli, A.P.P.; Passarelli, M.d.S.; de Freitas, F.V.; Ravagnani, G.M.; Torres, M.A.; Martins, S.M.M.K.; Yeste, M.; de Andrade, A.F.C. Removal of Seminal Plasma Prior to Liquid Storage of Boar Spermatozoa: A Practice That Can Improve Their Fertilizing Ability. Theriogenology 2019, 125, 79–86. [Google Scholar] [CrossRef]
- Rota, A.; Magelli, C.; Panzani, D.; Camillo, F. Effect of Extender, Centrifugation and Removal of Seminal Plasma on Cooled-Preserved Amiata Donkey Spermatozoa. Theriogenology 2008, 69, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, M.; Lehimcioğlu, N.C.; Akman, O. Effect of Seminal Plasma on Functional Integrity of Rabbit Sperm Membranes during Storage at 4 °C or Freezing. World Rabbit Sci. 2008, 16, 01–06. [Google Scholar] [CrossRef][Green Version]
- Zhu, J.; Xu, X.; Cosgrove, J.R.; Foxcroft, G.R. Effects of Semen Plasma from Different Fractions of Individual Ejaculates on IVF in Pigs. Theriogenology 2000, 54, 1443–1452. [Google Scholar] [CrossRef]
- Caballero, I.; Vazquez, J.M.; Centurión, F.; Rodríguez-Martinez, H.; Parrilla, I.; Roca, J.; Cuello, C.; Martinez, E.A. Comparative Effects of Autologous and Homologous Seminal Plasma on the Viability of Largely Extended Boar Spermatozoa. Reprod. Domest. Anim. 2004, 39, 370–375. [Google Scholar] [CrossRef]
- Maxwell, W.M.; Graaf, S.; Ghaoui, E.-H.; Evans, G.; de Graaf, S.P.S.; Ghaoui, R.E.-H.H.; Evans, G. Seminal Plasma Effects on Sperm Handling and Female Fertility. Soc. Reprod. Fertil. Suppl. 2007, 64, 13–38. [Google Scholar] [CrossRef] [PubMed]
- Killian, G.J.; Chapman, D.A.; Rogowski, L.A. Fertility-Associated Proteins in Holstein Bull Seminal Plasma. Biol. Reprod. 1993, 49, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Aurich, J.E.; Kühne, A.; Hoppe, H.; Aurich, C. Seminal Plasma Affects Membrane Integrity and Motility of Equine Spermatozoa after Cryopreservation. Theriogenology 1996, 46, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.F.; Anel-Lopez, L.; Neila-Montero, M.; Palacin-Martinez, C.; Montes-Garrido, R.; Alvarez, M.; de Paz, P.; Anel, L. ProAKAP4 as Novel Molecular Marker of Sperm Quality in Ram: An Integrative Study in Fresh, Cooled and Cryopreserved Sperm. Biomolecules 2020, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Brinsko, S.P. Insemination Doses: How Low Can We Go? Theriogenology 2006, 66, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Salamon, S.; Maxwell, W.M.C. Storage of Ram Semen. Anim. Reprod. Sci. 2000, 62, 77–111. [Google Scholar] [CrossRef] [PubMed]
- Dziekońska, A.; Partyka, A. Current Status and Advances in Semen Preservation. Animals 2023, 13, 123. [Google Scholar] [CrossRef] [PubMed]
- Gillan, L.; Evans, G.; Maxwell, W.M.C. Flow Cytometric Evaluation of Sperm Parameters in Relation to Fertility Potential. Theriogenology 2005, 63, 445–457. [Google Scholar] [CrossRef]
- Petrunkina, A.M.; Waberski, D.; Günzel-Apel, A.R.; Töpfer-Petersen, E. Determinants of Sperm Quality and Fertility in Domestic Species. Reproduction 2007, 134, 3–17. [Google Scholar] [CrossRef]
- Martínez-Pastor, F.; Mata-Campuzano, M.; Álvarez-Rodríguez, M.; Álvarez, M.; Anel, L.; de Paz, P. Probes and Techniques for Sperm Evaluation by Flow Cytometry. Reprod. Domest. Anim. 2010, 45, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Gündoǧan, M. Short Term Preservation of Ram Semen with Different Extenders. Kafkas Univ. Vet. Fak. Derg. 2009, 15, 429–435. [Google Scholar] [CrossRef]
- Maxwell, W.M.C.; Salomon, S. Liquid Storage of Ram Semen: A Review. Reprod. Fertil. Dev. 1993, 5, 613–638. [Google Scholar] [CrossRef] [PubMed]
- Roostaei-Ali Mehr, M.; Chambary, B.; Ghavi Hossein-Zadeh, N. Effect of Different Diluents and Storage Time on Field Fertility of Cooled Ram Semen after Vaginal Insemination. Small Rumin. Res. 2013, 115, 82–85. [Google Scholar] [CrossRef]
- Pellicer-Rubio, M.T.; Combarnous, Y. Deterioration of Goat Spermatozoa in Skimmed Milk-Based Extenders as a Result of Oleic Acid Released by the Bulbourethral Lipase BUSgp60. J. Reprod. Fertil. 1998, 112, 95–105. [Google Scholar] [CrossRef]
- Manjunath, P. New Insights into the Understanding of the Mechanism of Sperm Protection by Extender Components. Anim. Reprod. Sci. 2012, 9, 809–815. [Google Scholar]
- Watson, P.F. The Causes of Reduced Fertility with Cryopreserved Semen. Anim. Reprod. Sci. 2000, 60–61, 481–492. [Google Scholar] [CrossRef]
- Pursel, V.G.; Johnson, L.A.; Rampacek, G.B. Acrosome Morphology of Boar Spermatozoa Incubated before Cold Shock. J. Anim. Sci. 1972, 34, 278–283. [Google Scholar] [CrossRef]
- Falchi, L.; Galleri, G.; Zedda, M.T.; Pau, S.; Bogliolo, L.; Ariu, F.; Ledda, S. Liquid Storage of Ram Semen for 96 h: Effects on Kinematic Parameters, Membranes and DNA Integrity, and ROS Production. Livest. Sci. 2018, 207, 1–6. [Google Scholar] [CrossRef]
- Shi, L.; Jin, T.; Hu, Y.; Ma, Z.; Niu, H.; Ren, Y. Effects of Reduced Glutathione on Ram Sperm Parameters, Antioxidant Status, Mitochondrial Activity and the Abundance of Hexose Transporters during Liquid Storage at 5 °C. Small Rumin. Res. 2020, 189, 106139. [Google Scholar] [CrossRef]
- Albiaty, N.M.; Alobaidi, H.J.; Kareem, A.F.; Al-Hakim, A.M.; Alnaeb, A.Y. Effect of Extenders and Preservation Periods in Some Semen Characteristics of Awassi Rams. World J. Pharm. Res. 2016, 5, 234–243. [Google Scholar]
- Yániz, J.L.; Santolaria, P.; Marco-Aguado, M.A.; López-Gatius, F. Use of Image Analysis to Assess the Plasma Membrane Integrity of Ram Spermatozoa in Different Diluents. Theriogenology 2008, 70, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.A.P.; Dott, H.M.; Foster, G.C. Bovine Serum Albumin, Sperm Motility, and the “Dilution Effect”. J. Exp. Zool. 1982, 222, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, P.J.C.; Harrison, R.A.P.; Miller, N.G.A.; Plummer, J.M.; Watson, P.F. Survival of Ram Spermatozoa at High Dilution: Protective Effect of Simple Constituents of Culture Media as Compared with Seminal Plasma. Reprod. Fertil. Dev. 1994, 6, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Castellini, C.; Lattaioli, P.; Moroni, M.; Minelli, A. Effect of Seminal Plasma on the Characteristics and Fertility of Rabbit Spermatozoa. Anim. Reprod. Sci. 2000, 63, 275–282. [Google Scholar] [CrossRef]
- Caballero, I.; Vázquez, J.M.; García, E.M.; Roca, J.; Martínez, E.A.; Calvete, J.J.; Sanz, L.; Ekwall, H.; Rodríguez-Martínez, H. Immunolocalization and Possible Functional Role of PSP-I/PSP-II Heterodimer in Highly Extended Boar Spermatozoa. J. Androl. 2006, 27, 766–773. [Google Scholar] [CrossRef]
- Garner, D.L.; Thomas, C.A.; Gravance, C.G.; Marshall, C.E.; DeJarnette, J.M.; Allen, C.H. Seminal Plasma Addition Attenuates the Dilution Effect in Bovine Sperm. Theriogenology 2001, 56, 31–40. [Google Scholar] [CrossRef]
- Catt, S.L.; O’Brien, J.K.; Maxwell, W.M.C.; Evans, G. Assessment of Ram and Boar Spermatozoa during Cell-Sorting by Flow Cytometry. Reprod. Domest. Anim. 1997, 32, 251–258. [Google Scholar] [CrossRef]
- Paul, R.K.; Balaganur, K.; Bahire, S.V.; Kumar, D.; Singh, R. Supplementation of Cauda Epididymal Plasma Improves Sperm Characteristics Following Liquid Preservation of Ram Semen at 3–5 °C. Reprod. Fertil. Dev. 2018, 30, 1389–1401. [Google Scholar] [CrossRef]
- Herold, F.C.; de Haas, K.; Colenbrander, B.; Gerber, D. Comparison of Equilibration Times When Freezing Epididymal Sperm from African Buffalo (Syncerus Caffer) Using TriladylTM or AndroMed®. Theriogenology 2006, 66, 1123–1130. [Google Scholar] [CrossRef]
- Manjunath, P.; Thérien, I. Role of Seminal Plasma Phospholipid-Binding Proteins in Sperm Membrane Lipid Modification That Occurs during Capacitation. J. Reprod. Immunol. 2002, 53, 109–119. [Google Scholar] [CrossRef]
- Lusignan, M.F.; Bergeron, A.; Crête, M.H.; Lazure, C.; Manjunath, P. Induction of Epididymal Boar Sperm Capacitation by PB1 and BSP-A1/-A2 Proteins, Members of the BSP Protein Family. Biol. Reprod. 2007, 76, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Manjunath, P.; Bergeron, A.; Lefebvre, J.; Fan, J. Seminal Plasma Proteins: Functions and Interaction with Protective Agents during Semen Preservation. Soc. Reprod. Fertil. Suppl. 2007, 65, 217–228. [Google Scholar] [PubMed]
- Plante, G.; Lusignan, M.F.; Lafleur, M.; Manjunath, P. Interaction of Milk Proteins and Binder of Sperm (BSP) Proteins from Boar, Stallion and Ram Semen. Reprod. Biol. Endocrinol. 2015, 13, 92. [Google Scholar] [CrossRef] [PubMed]
- Desnoyers, L.; Manjunath, P. Major Proteins of Bovine Seminal Plasma Exhibit Novel Interactions with Phospholipid. J. Biol. Chem. 1992, 267, 10149–10155. [Google Scholar] [CrossRef] [PubMed]
- Anzar, M.; Kroetsch, T.; Boswall, L. Cryopreservation of Bull Semen Shipped Overnight and Its Effect on Post-Thaw Sperm Motility, Plasma Membrane Integrity, Mitochondrial Membrane Potential and Normal Acrosomes. Anim. Reprod. Sci. 2011, 126, 23–31. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neila-Montero, M.; Alvarez, M.; Riesco, M.F.; Soriano-Úbeda, C.; Montes-Garrido, R.; Palacin-Martinez, C.; de Paz, P.; Anel, L.; Anel-Lopez, L. The Adaptation Time to the Extender as a Crucial Step for an Accurate Evaluation of Ram Sperm Quality during the Liquid Storage. Vet. Sci. 2024, 11, 132. https://doi.org/10.3390/vetsci11030132
Neila-Montero M, Alvarez M, Riesco MF, Soriano-Úbeda C, Montes-Garrido R, Palacin-Martinez C, de Paz P, Anel L, Anel-Lopez L. The Adaptation Time to the Extender as a Crucial Step for an Accurate Evaluation of Ram Sperm Quality during the Liquid Storage. Veterinary Sciences. 2024; 11(3):132. https://doi.org/10.3390/vetsci11030132
Chicago/Turabian StyleNeila-Montero, Marta, Mercedes Alvarez, Marta F. Riesco, Cristina Soriano-Úbeda, Rafael Montes-Garrido, Cristina Palacin-Martinez, Paulino de Paz, Luis Anel, and Luis Anel-Lopez. 2024. "The Adaptation Time to the Extender as a Crucial Step for an Accurate Evaluation of Ram Sperm Quality during the Liquid Storage" Veterinary Sciences 11, no. 3: 132. https://doi.org/10.3390/vetsci11030132
APA StyleNeila-Montero, M., Alvarez, M., Riesco, M. F., Soriano-Úbeda, C., Montes-Garrido, R., Palacin-Martinez, C., de Paz, P., Anel, L., & Anel-Lopez, L. (2024). The Adaptation Time to the Extender as a Crucial Step for an Accurate Evaluation of Ram Sperm Quality during the Liquid Storage. Veterinary Sciences, 11(3), 132. https://doi.org/10.3390/vetsci11030132