

Tick Diversity and Pathogen Transmission in Daejeon, Korea: Implications from Companion Animals and Walking Trails

 and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Identification of Tick Species and Developmental Stages
2.3. DNA Extraction for Polymerase Chain Reaction Amplification
2.4. Real-Time Polymerase Chain Reaction
2.5. SFTSV Conventional (Nested) Polymerase Chain Reaction
2.6. Anaplasma/Ehrlichia Detection Using Conventional (Nested) Polymerase Chain Reaction
3. Results
3.1. Tick Species and Developmental Stages of Collected Ticks
3.2. Distribution of Tick and Pathogen Detection on Urban Walking Trails
3.3. Distribution of Ticks and Pathogen Detection on Pet Trails
3.4. Distribution of Ticks and Pathogen Detection from Companion Animals
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef]
- Mrzljak, A.; Novak, R.; Pandak, N.; Tabain, I.; Franusic, L.; Barbic, L.; Bogdanic, M.; Savic, V.; Mikulic, D.; Pavicic-Saric, J.; et al. Emerging and neglected zoonoses in transplant population. World J. Transplant. 2020, 10, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Khoo, J.J.; Chen, F.; Kho, K.L.; Shanizza, A.I.A.; Lim, F.S.; Tan, K.K.; Chang, L.Y.; AbuBakar, S. Bacterial community in Haemaphysalis ticks of domesticated animals from the Orang Asli communities in Malaysia. Ticks Tick Borne Dis. 2016, 7, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Campbell-Lendrum, D.; Holloway, T.; Foley, J.A. Impact of regional climate change on human health. Nature 2005, 438, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Süss, J.; Klaus, C.; Gerstengarbe, F.W.; Werner, P.C. What makes ticks tick? Climate change, ticks, and tick-borne diseases. J. Travel Med. 2008, 15, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, P.A. Climate change impacts on ticks and tick-borne infections. Biologia 2022, 77, 1503–1512. [Google Scholar] [CrossRef]
- Bouchard, C.; Dibernardo, A.; Koffi, J.; Wood, H.; Leighton, P.A.; Lindsay, L.R. Climate change and infectious diseases: The challenges: N increased risk of tick-borne diseases with climate and environmental changes. Can. Commun. Dis. Rep. CCDR 2019, 45, 83. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, S.; Bouchard, C.; Ogden, N.H.; Adam-Poupart, A.; Pelcat, Y.; Rees, E.E.; Milord, F.; Leighton, P.A.; Lindsay, R.L.; Koffi, J.K.; et al. Evidence for increasing densities and geographic ranges of tick species of public health significance other than Ixodes scapularis in Québec, Canada. PLoS ONE 2018, 13, e0201924. [Google Scholar] [CrossRef] [PubMed]
- Bush, E.; Lemmen, D.S. Canada’s Changing Climate Report; Government of Canada: Ottawa, ON, Canada, 2019; 444p. [Google Scholar]
- Jaenson, T.G.; Jaenson, D.G.; Eisen, L.; Petersson, E.; Lindgren, E. Changes in the geographical distribution and abundance of the tick Ixodes ricinus during the past 30 years in Sweden. Parasites Vectors 2012, 5, 8. [Google Scholar] [CrossRef]
- Hvidsten, D.; Frafjord, K.; Gray, J.S.; Henningsson, A.J.; Jenkins, A.; Kristiansen, B.E.; Lager, M.; Rognerud, B.; Slatsve, A.M.; Stordal, F.; et al. The distribution limit of the common tick, Ixodes ricinus, and some associated pathogens in north-western Europe. Ticks Tick Borne Dis. 2020, 11, 101388. [Google Scholar] [CrossRef]
- Colwell, D.D.; Dantas-Torres, F.; Otranto, D. Vector-borne parasitic zoonoses: Emerging scenarios and new perspectives. Vet. Parasitol. 2011, 182, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.H.; Hinckley, A.F.; Hook, S.A.; Meek, J.I.; Backenson, B.; Kugeler, K.J.; Feldman, K.A. Pet ownership increases human risk of encountering ticks. Zoonoses Public Health 2018, 65, 74–79. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Meteorological Research. Climate Change of Korea: Present State and Future; National Institute of Meteorological Research: Seogwipo, Republic of Korea, 2009. [Google Scholar]
- Lee, H.; Kim, G.; Park, C.; Cha, D. A Study of Future Changes of Climate Classification and Extreme Temperature Events over South Korea in Multi Regional Climate Model Simulations. J. Clim. Res. 2017, 12, 149–164. [Google Scholar] [CrossRef]
- Park, C.Y.; Choi, Y.E.; Kwon, Y.A.; Kwon, J.I.; Lee, H.S. Studies on Changes and Future Projections of Subtropical Climate Zones and Extreme Temperature Events over South Korea Using High Resolution Climate Change Scenario Based on PRIDE Model. J. Korean Assoc. Reg. Geogr. 2013, 19, 600–614. [Google Scholar]
- Im, J.H.; Baek, J.; Durey, A.; Kwon, H.Y.; Chung, M.H.; Lee, J.S. Current status of tick-borne diseases in South Korea. Vector Borne Zoonotic Dis. 2019, 19, 225–233. [Google Scholar] [CrossRef]
- Liu, S.; Chai, C.; Wang, C.; Amer, S.; Lv, H.; He, H.; Sun, J.; Lin, J. Systematic review of severe fever with thrombocytopenia syndrome: Virology, epidemiology, and clinical characteristics. Rev. Med. Virol. 2014, 24, 90–102. [Google Scholar] [CrossRef]
- Yamaguti, N.; Tipton, V.J.; Keegan, H.L.; Toshioka, S. Ticks of Japan, Korea, and the Ryukyu islands. Brigh. Young Univ. Sci. Bull. Biol. Ser. 1971, 15, 1. [Google Scholar]
- Intirach, J.; Lv, X.; Han, Q.; Lv, Z.Y.; Chen, T. Morphological and Molecular Identification of Hard Ticks in Hainan Island, China. Genes 2023, 14, 1592. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Fu, Y.; Li, S.; Zhang, Z.; Man, S.; Li, X.; Zhang, W.; Zhang, C.; Cheng, X. Phylogeographic analysis of severe fever with thrombocytopenia syndrome virus from Zhoushan Islands, China: Implication for transmission across the ocean. Sci. Rep. 2016, 6, 19563. [Google Scholar] [CrossRef]
- Kim, C.M.; Yi, Y.H.; Yu, D.H.; Lee, M.J.; Cho, M.R.; Desai, A.R.; Shringi, S.; Klein, T.A.; Kim, H.C.; Song, J.W.; et al. Tick-borne rickettsial pathogens in ticks and small mammals in Korea. Appl. Environ. Microbiol. 2006, 72, 5766–5776. [Google Scholar] [CrossRef] [PubMed]
- Noda, A.A.; Rodríguez, I.; Mondeja, B.; Fernándeze, C. Design, optimization and evaluation of a polymerase chain reaction for detection of Borrelia spp. Adv. Clin. Exp. Med. 2013, 22, 639–653. [Google Scholar] [PubMed]
- Corrigan, J.; Marion, B.; English, J.; Eneku, W.; Weng, J.L.; Rugg, M.; Dotrang, T.; Dunford, J.; Byaruhanga, A.M.; Ramalho-Ortigao, M.; et al. Minimal rickettsial infection rates and distribution of ticks in Uganda: An assessment of the seasonal effects and relevance to tick-borne disease risk in East Africa. J. Med. Entomol. 2023, 60, 185–192. [Google Scholar] [CrossRef] [PubMed]
- John, H.K.S.; Masuoka, P.; Jiang, J.; Takhampunya, R.; Klein, T.A.; Kim, H.C.; Chong, S.T.; Song, J.W.; Kim, Y.J.; Farris, C.M.; et al. Geographic distribution and modeling of ticks in the Republic of Korea and the application of tick models towards understanding the distribution of associated pathogenic agents. Ticks Tick Borne Dis. 2021, 12, 101686. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, J.; Cui, X.; Jia, N.; Wei, J.; Xia, L.; Wang, H.; Zhou, Y.; Wang, Q.; Liu, X.; et al. Distribution of Haemaphysalis longicornis and associated pathogens: Analysis of pooled data from a China field survey and global published data. Lancet Planet. Health 2020, 4, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, S.H.; Shin, S.; Kwak, D. Molecular identification of Borrelia spp. from ticks in pastures nearby livestock farms in Korea. Insects 2021, 12, 1011. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Meng, C.; Li, Y.; Zhou, G.; Wang, L.; Xu, X.; Li, N.; Ji, Y.; Tian, J.; Wang, W.; et al. Rickettsia sp. and Anaplasma spp. in Haemaphysalis longicornis from Shandong province of China, with evidence of a novel species “Candidatus Anaplasma Shandongensis”. Ticks Tick Borne Dis. 2023, 14, 102082. [Google Scholar] [CrossRef]
- Suh, J.H.; Lim, J.W.; Kang, D.H.; Kim, H.C. Taxonomy and Occurences of Disease Mediating Ricks. National Institute of Biological Resources (NIBR. 201501110). National R&D Report. 2015. Available online: https://scienceon.kisti.re.kr/commons/util/originalView.do?cn=TRKO201600001544&dbt=TRKO&rn= (accessed on 29 November 2023).
- Seo, M.G.; Noh, B.E.; Lee, H.S.; Kim, T.K.; Song, B.G.; Lee, H.I. Nationwide temporal and geographical distribution of tick populations and phylogenetic analysis of severe fever with thrombocytopenia syndrome virus in ticks in Korea, 2020. Microorganisms 2021, 9, 1630. [Google Scholar] [CrossRef]
- Oh, S.S.; Chae, J.B.; Kang, J.G.; Kim, H.C.; Chong, S.T.; Shin, J.H.; Hur, M.S.; Suh, J.H.; Oh, M.D.; Jeong, S.M.; et al. Detection of severe fever with thrombocytopenia syndrome virus from wild animals and Ixodidae ticks in the Republic of Korea. Vector Borne Zoonotic Dis. 2016, 16, 408–414. [Google Scholar] [CrossRef]
- Seo, M.G.; Lee, H.; Alkathiri, B.; Ahn, K.; Lee, S.H.; Shin, S.; Bae, S.; Kim, K.; Jang, M.; Lee, S.; et al. Tick Populations and Molecular Analysis of Anaplasma Species in Ticks from the Republic of Korea. Microorganisms 2023, 11, 820. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, C.; Lynn, G.E.; Pedra, J.H.; Pal, U.; Narasimhan, S.; Fikrig, E. Interactions between Borrelia burgdorferi and ticks. Nat. Rev. Microbiol. 2020, 18, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Kida, K.; Matsuoka, Y.; Shimoda, T.; Matsuoka, H.; Yamada, H.; Saito, T.; Imataki, O.; Kadowaki, N.; Noguchi, K.; Maeda, K.; et al. A case of cat-to-human transmission of severe fever with thrombocytopenia syndrome virus. Jpn. J. Infect. Dis. 2019, 72, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Crump, A.; Tanimoto, T. Severe fever with thrombocytopenia syndrome: Japan under threat from life-threatening emerging tick-borne disease. Jpn. Med. Assoc. J. 2020, 3, 295–302. [Google Scholar]
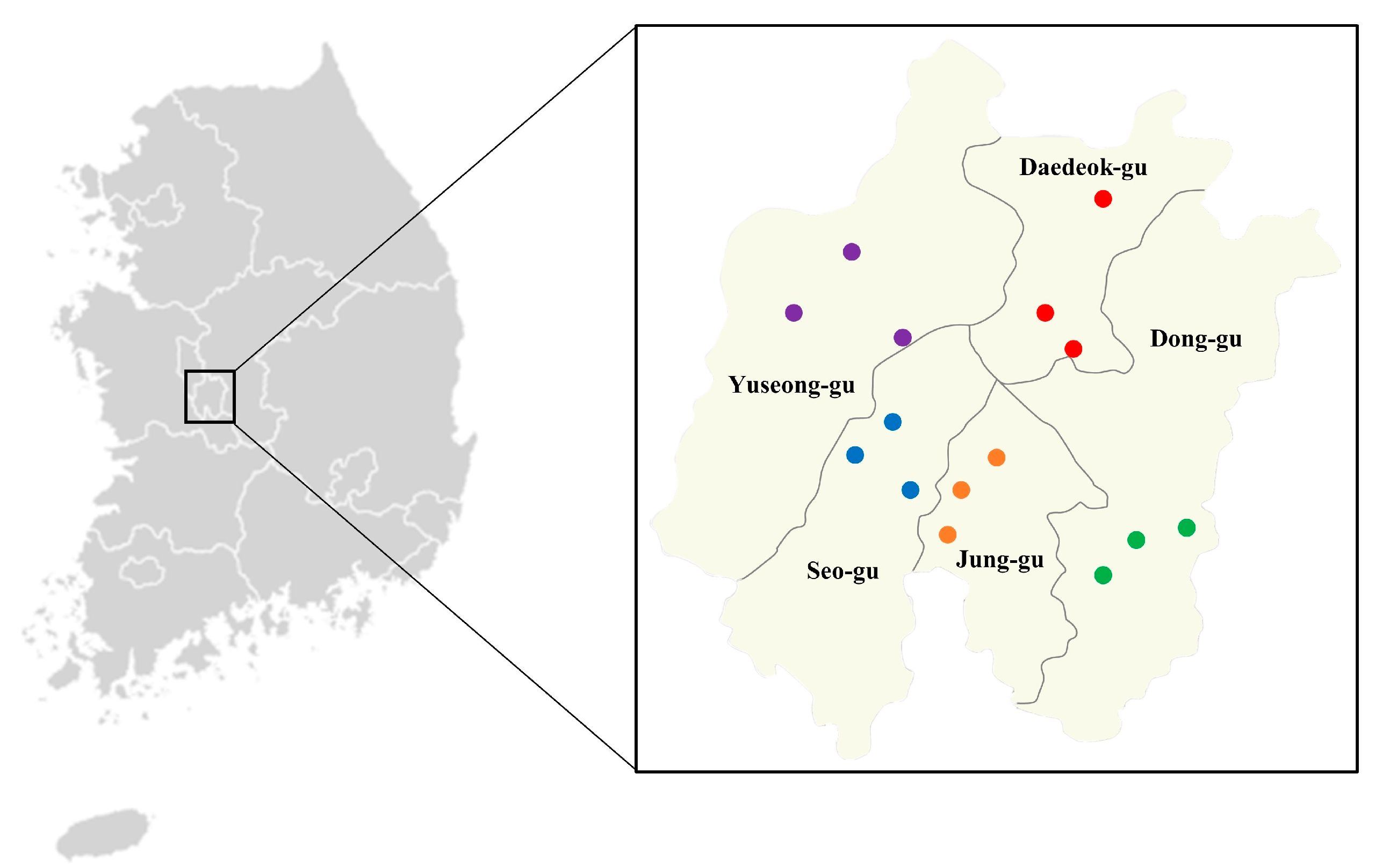

{kind=link}
| District | Administrative Location | GPS Location |
|---|---|---|
| Dong-gu | Gao Neighborhood park | 36°30′63″ N 127°45′87″ E |
| Choji park | 36°27′85″ N 127°46′22″ E | |
| Sanseo-ro | 36°28′35″ N 127°46′8″ E | |
| Jung-gu | Sajeong park | 36°29′92″ N 127°40′69″ E |
| Daedunsan-ro | 36°28′90″ N 127°37′23″ E | |
| Baekcheon Buddhist temple | 36°29′92″ N 127°39′56″ E | |
| Seo-gu | Sotae Neighborhood park | 36°31′62″ N 127°34′37″ E |
| Wolpyeong park | 36°32′26″ N 127°36′02″ E | |
| Whaum Buddhist temple | 36°29′90″ N 127°36′64″ E | |
| Yuseong-gu | Sundusan Neighborhood park | 36°37′25″ N 127°37′39″ E |
| Sinsung Neighborhood park | 36°38′5″ N 127°35′35″ E | |
| Bokyong Urban natural park | 36°34′29″ N 127°53′18″ E | |
| Daedeok-gu | Gilchi-Neighborhood park | 36°36′19″ N 127°45′82″ E |
| Birae Buddhist temple | 36°37′9″ N 127°44′88″ E | |
| Jangdong Forest park | 36°40′65″ N 127°43′88″ E |
| Target Pathogen | Primer | Nucleotide Sequences (5′ → 3′) | Target Gene (Product Size) | Reference |
|---|---|---|---|---|
| SFTSV | MF3 MR2 | GATGAGATGGTCCATGCTGATTCT CTCATGGGGTGGAATGTCCTCAC | M segment (560 bp) | [22] |
| MMF3 MMF2 | TAAACTTGTGTCGTGCAGGC CCCAGCGACATCTCCTTACA | M segment (245 bp) | ||
| Anaplasma spp. | AE1-F1 AE1-R1 | AAGCTTAACACATGCAAGTCGAA AGTCACTGACCCAACCTTAAATG | 16S rRNA (1406 bp) | [23] |
| AP-F AP-R | GTCGAACGGATTATTCTTTATAGCTTGC CCCTTCCGTTAAGAAGGATCTAATCTCC | 16S rRNA (926 bp) | ||
| Ehrlichia spp. | AE1-F1 AE1-R1 | AAGCTTAACACATGCAAGTCGAA AGTCACTGACCCAACCTTAAATG | 16S rRNA (1406 bp) | [23] |
| EC-F2 EC-R2 | CAATTGCTTATAACCTTTTGGTTATAAAT TATAGGTACCGTCATTATCTTCCCTAT | 16S rRNA (390 bp) | ||
| Borrelia spp. | B1 B2 | TAGATGAGTCTGCGTCTTATTA CTTACACCAGGAATTCTAACTT | 16S rRNA (465 bp) | [24] |
| Species of Ticks | Stages | No. of Ticks | No. of Pools | No. of Detected Pathogens | Total | |||
|---|---|---|---|---|---|---|---|---|
| SFTSV | Anaplasma spp. | Ehrlichia spp. | Borrelia spp. | |||||
| H. longicornis | Adult Nymph Larva Subtotal | 1074 3952 9923 14,949 | 279 318 251 848 | 2 0 0 2 | 0 0 0 0 | 0 0 0 0 | 6 6 1 13 | 8 6 1 15 |
| H. flava | Adult Nymph Larva Subtotal | 76 224 711 1011 | 51 65 35 151 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 1 0 1 | 0 1 0 1 |
| I. nipponensis | Adult Nymph Larva Subtotal | 51 16 775 842 | 58 11 27 86 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 14 4 0 18 | 14 4 0 18 |
| A. testudinarium | Adult Nymph Larva Subtotal | 0 1 0 1 | 0 1 0 1 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 |
| Total | 16,803 | 1086 | 2 | 0 | 0 | 32 | 34 | |
| MIR (%) | - | - | 0.01 | 0 | 0 | 0.19 | - | |
| Species of Ticks | Stages | No. of Ticks | No. of Pools | No. of Detected Pathogens | Total | |||
|---|---|---|---|---|---|---|---|---|
| SFTSV | Anaplasma spp. | Ehrlichia spp. | Borrelia spp. | |||||
| H. longicornis | Adult Nymph Larva Subtotal | 1591 7232 1090 9913 | 186 212 23 421 | 1 0 0 1 | 0 0 0 0 | 3 2 0 5 | 0 0 0 0 | 4 2 0 6 |
| H. flava | Adult Nymph Larva Subtotal | 123 164 772 1059 | 51 26 21 98 | 0 0 0 0 | 0 0 0 0 | 1 1 0 2 | 0 0 0 0 | 1 1 0 2 |
| I. nipponensis | Adult Nymph Larva Subtotal | 33 10 0 43 | 25 4 0 29 | 0 0 0 0 | 0 0 0 0 | 1 0 0 1 | 3 0 0 3 | 4 0 0 4 |
| A. testudinarium | Adult Nymph Larva Subtotal | 0 1 0 1 | 0 1 0 1 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 |
| Total | 11,016 | 549 | 1 | 0 | 8 | 3 | 12 | |
| MIR (%) | - | - | 0.01 | 0 | 0.07 | 0.03 | - | |
| Species of Ticks | Stages | No. of Ticks | No. of Pools | No. of Detected Pathogens | Total | |||
|---|---|---|---|---|---|---|---|---|
| SFTSV | Anaplasma spp. | Ehrlichia spp. | Borrelia spp. | |||||
| H. longicornis | Adult Nymph Larva Subtotal | 275 35 0 310 | 93 11 0 104 | 1 0 0 1 | 1 0 0 1 | 2 0 0 2 | 1 0 0 1 | 5 0 0 5 |
| H. flava | Adult Nymph Larva Subtotal | 1 0 0 1 | 1 0 0 1 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 |
| I. nipponensis | Adult Nymph Larva Subtotal | 34 1 0 35 | 24 1 0 25 | 0 0 0 0 | 2 0 0 2 | 1 0 0 1 | 0 0 0 0 | 3 0 0 3 |
| Total | 346 | 130 | 1 | 3 | 3 | 1 | 8 | |
| MIR (%) | - | - | 0.29 | 0.87 | 0.87 | 0.29 | - | |
| Species of Ticks | Stages | No. of Ticks | No. of Pools | No. of Detected Pathogens | Total | |||
|---|---|---|---|---|---|---|---|---|
| SFTSV | Anaplasma spp. | Ehrlichia spp. | Borrelia spp. | |||||
| H. longicornis | Adult Nymph Larva Subtotal | 225 514 25 764 | 32 25 3 60 | 0 0 0 0 | 1 0 0 1 | 0 0 0 0 | 1 0 0 1 | 2 0 0 2 |
| H. flava | Adult Nymph Larva Subtotal | 66 74 0 140 | 10 5 0 15 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 | 0 0 0 0 |
| I. nipponensis | Adult Nymph Larva Subtotal | 107 0 0 107 | 18 0 0 18 | 0 0 0 0 | 1 0 0 1 | 0 0 0 0 | 0 0 0 0 | 1 0 0 1 |
| Total | 1011 | 93 | 0 | 2 | 0 | 1 | 3 | |
| MIR (%) | - | - | 0 | 0.2 | 0 | 0.1 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, J.; Kim, G.; Lim, J.-a.; Song, S.; Yoo, D.-S.; Cho, H.-S.; Oh, Y. Tick Diversity and Pathogen Transmission in Daejeon, Korea: Implications from Companion Animals and Walking Trails. Vet. Sci. 2024, 11, 90. https://doi.org/10.3390/vetsci11020090
Seo J, Kim G, Lim J-a, Song S, Yoo D-S, Cho H-S, Oh Y. Tick Diversity and Pathogen Transmission in Daejeon, Korea: Implications from Companion Animals and Walking Trails. Veterinary Sciences. 2024; 11(2):90. https://doi.org/10.3390/vetsci11020090
Chicago/Turabian StyleSeo, Jinwoo, Gyurae Kim, Jeong-ah Lim, Seungho Song, Dae-Sung Yoo, Ho-Seong Cho, and Yeonsu Oh. 2024. "Tick Diversity and Pathogen Transmission in Daejeon, Korea: Implications from Companion Animals and Walking Trails" Veterinary Sciences 11, no. 2: 90. https://doi.org/10.3390/vetsci11020090
APA StyleSeo, J., Kim, G., Lim, J.-a., Song, S., Yoo, D.-S., Cho, H.-S., & Oh, Y. (2024). Tick Diversity and Pathogen Transmission in Daejeon, Korea: Implications from Companion Animals and Walking Trails. Veterinary Sciences, 11(2), 90. https://doi.org/10.3390/vetsci11020090