4. Discussion
In order to examine the vasculature of the feet in birds of prey and owls, species were chosen for investigation because they can be taxonomically assigned to the three orders of interest:
Accipitriformes—northern goshawk and common buzzard;
Falconiformes—peregrine falcon, gyr–saker falcon, and common kestrel;
Strigiformes—Eurasian eagle–owl, long-eared owl, and barn owl [
28]. Out of these, the northern goshawk, peregrine falcon, and gyr–saker falcon are popular birds in falconry, while some others, e.g., the Eurasian eagle–owl, are commonly kept in zoological facilities. In addition, except for the gyr–saker falcon, all species are native to Germany and injured or ill birds are regularly admitted to clinics or wildlife rescue centers. Thus, we were able to collect at least six carcasses from each species for subsequent examination. Like several other researchers, e.g., [
4,
24,
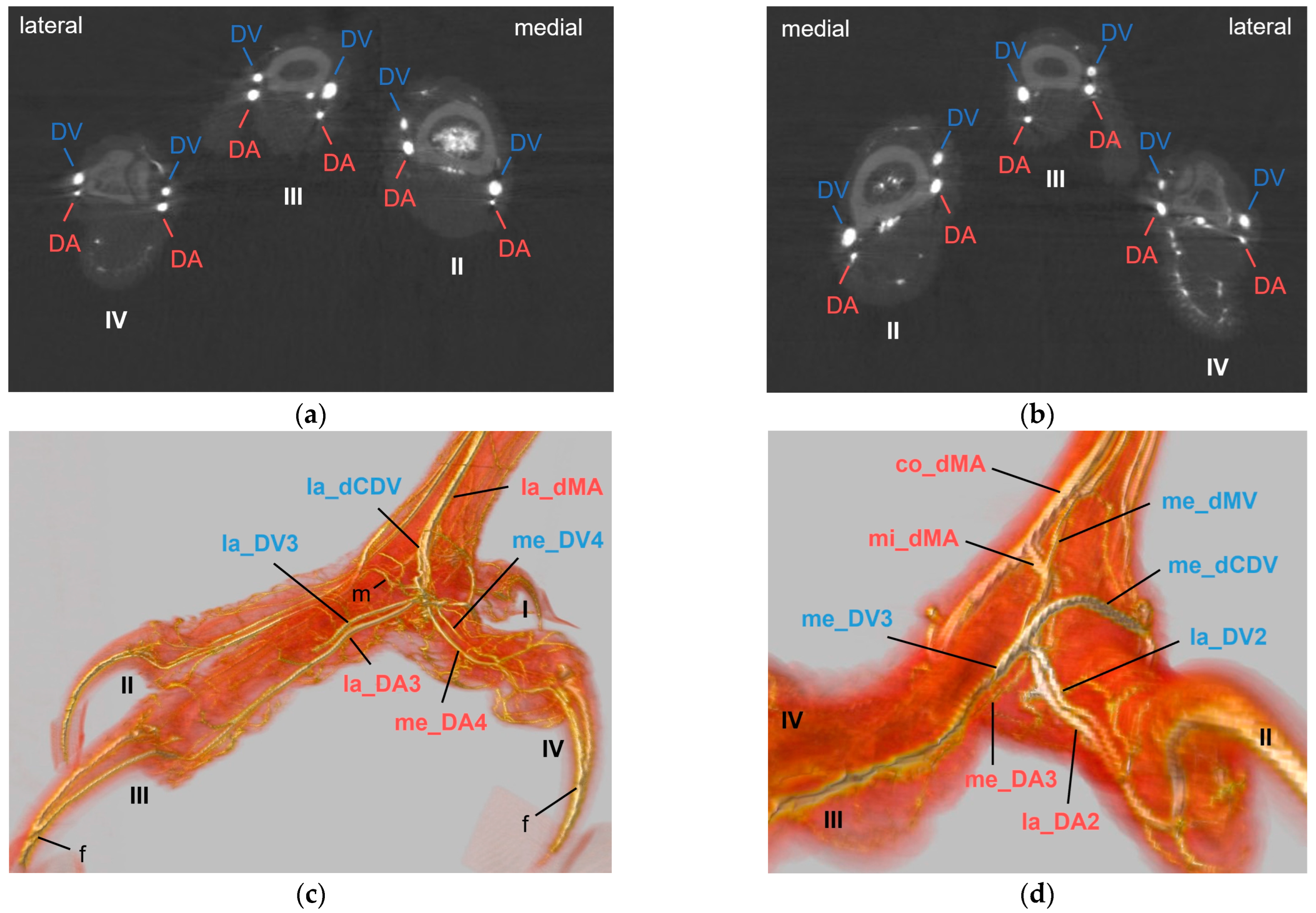
29], we used colored latex to fill either arteries or veins, with the advantage that latex does not leak after hardening and remains flexible, reducing the risk of breaking during dissection. In addition, we performed micro-computed tomography scans after the injection of contrast medium, allowing a 3D visualization of the blood vessel topography, even of the intraosseous courses of the vessels. The position of the arteries and veins in relation to each other became particularly clear, but in contrast to macroscopic dissection, it was more difficult to distinguish between arteries and veins due to the overlapping of the vessels. In conclusion, both methods used in this study complemented each other and were analyzed together to completely describe the topography of arteries and veins of the foot in the examined avian species.
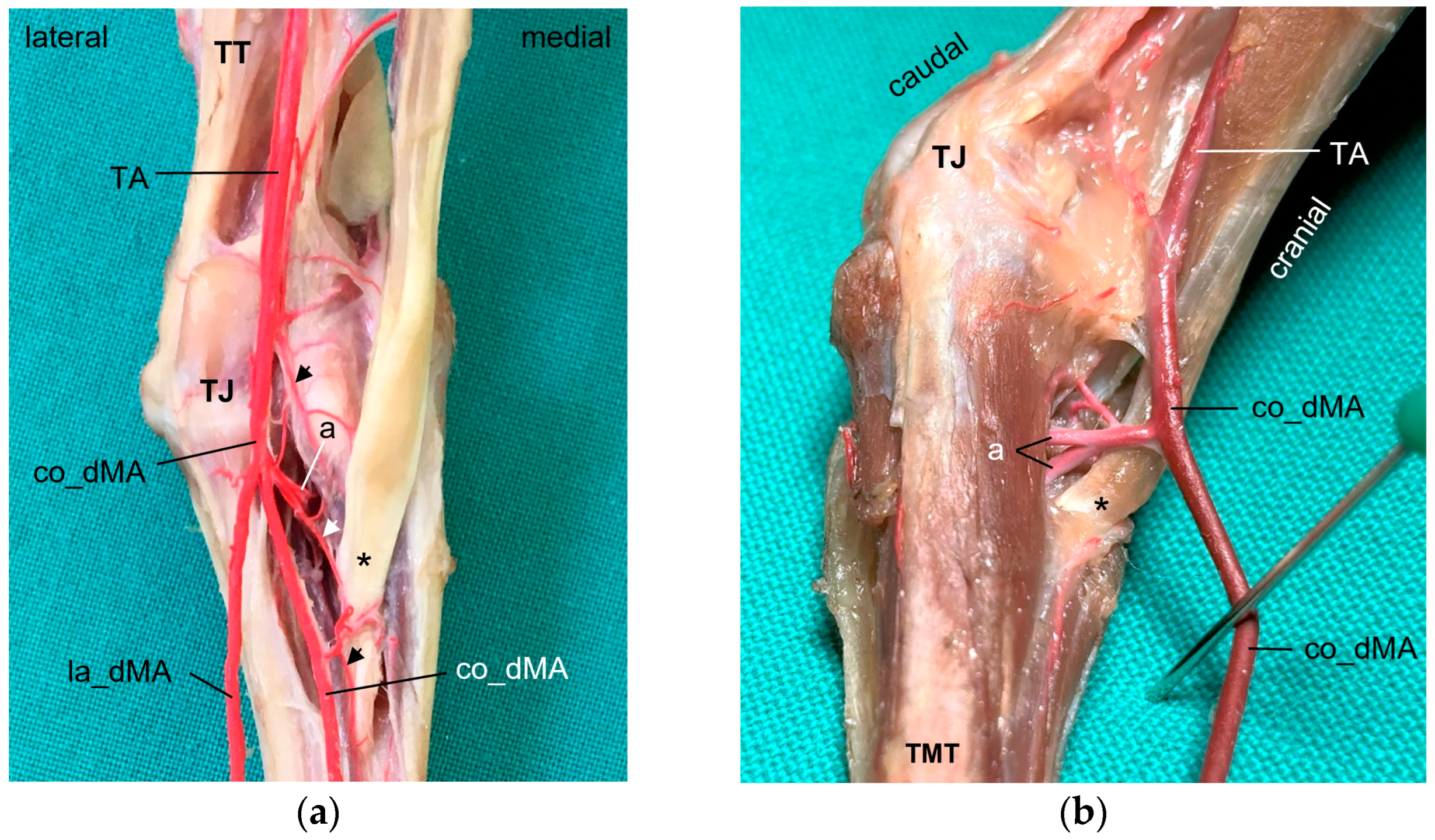
In the distal region of the tibiotarsus and at the intertarsal joint, we found a network of collateral branches formed by the cranial tibial artery. In the literature, this arterial network is called tibiotarsal rete [
22,
30] or tarsal rete [
29,
31]. It can be derived not only from the cranial tibial artery but also from the peroneal, also named fibular, artery, and “differs species specifically in its complexity” according to Midtgård [
22]. As the structures distal to the intertarsal joint were in our focus of examination, this rete was not further examined.
In all eight avian species examined, the cranial tibial artery continued distally at the tarsometatarsus as
common dorsal metatarsal artery, which was by far the largest artery proximal at the tarsometatarsus and therefore represented the main arterial supply of the foot. This is in accordance with previous studies on some species of birds of prey, i.e., northern goshawk [
4,
23,
25], southern caracara (
Caracara plancus) [
24], and peregrine falcon [
4,
23], and is consistently described in textbooks about avian anatomy for the domestic chicken (
Gallus gallus domesticus) [
30,
31,
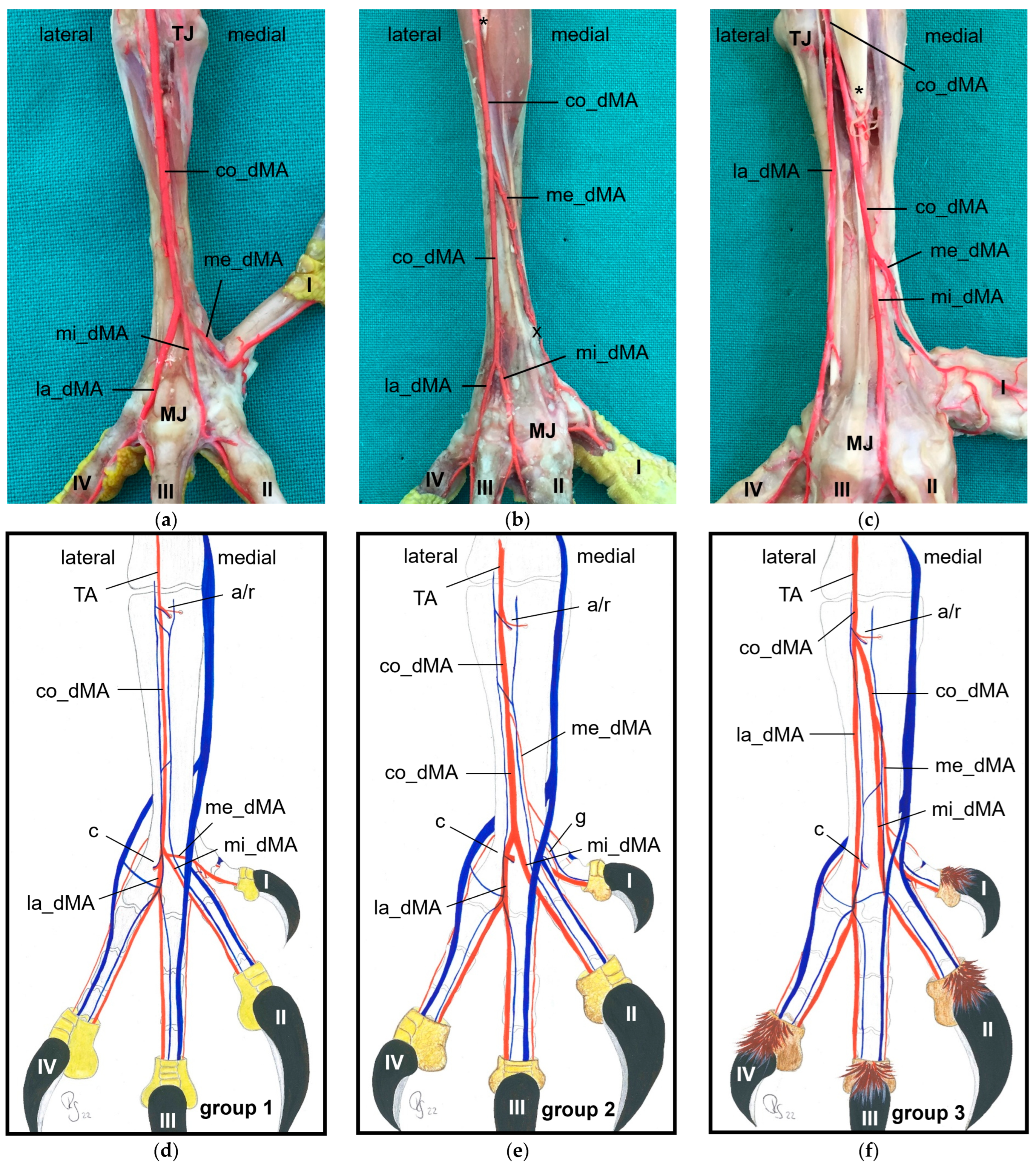
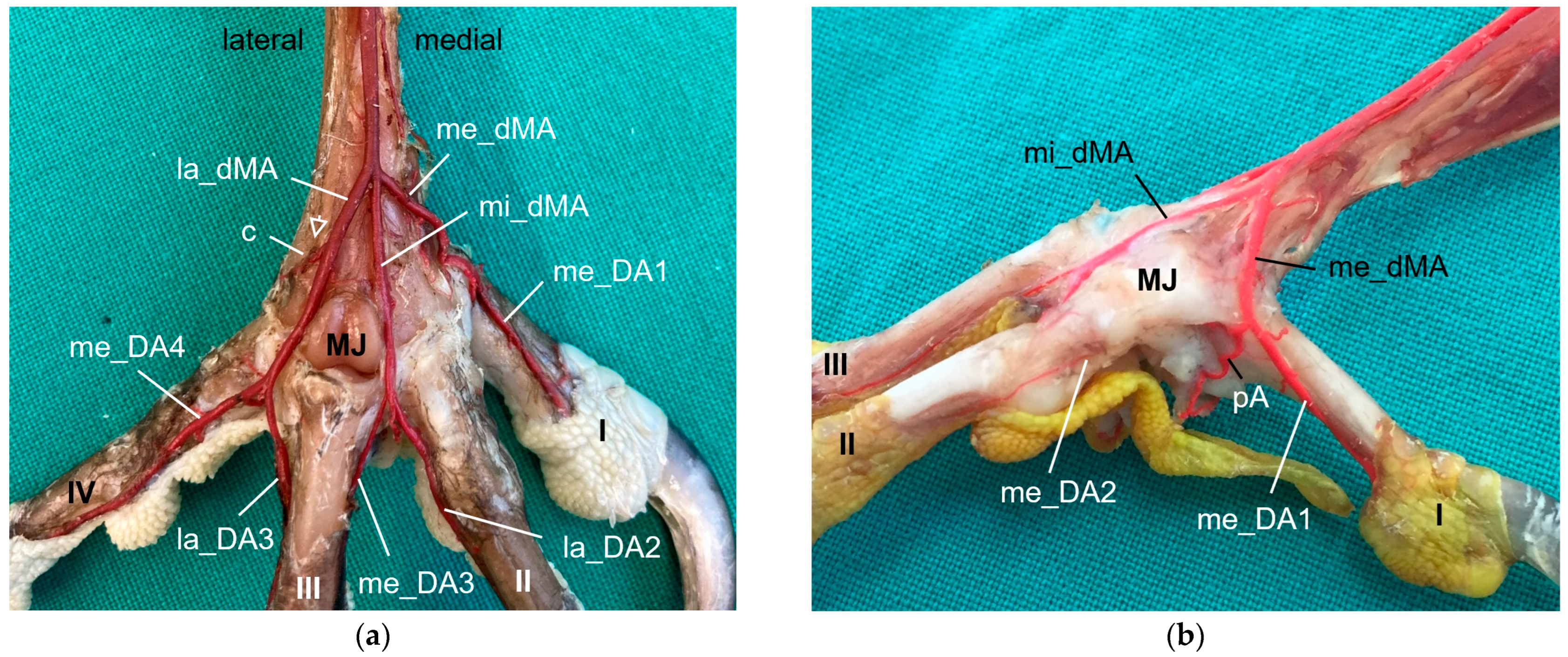
32]. The present study showed that the common dorsal metatarsal artery divided into three metatarsal arteries, the
lateral, middle, and medial metatarsal arteries, in all eight examined avian species. However, the point of division was species-specific, which allowed us to divide the species studied into three groups.
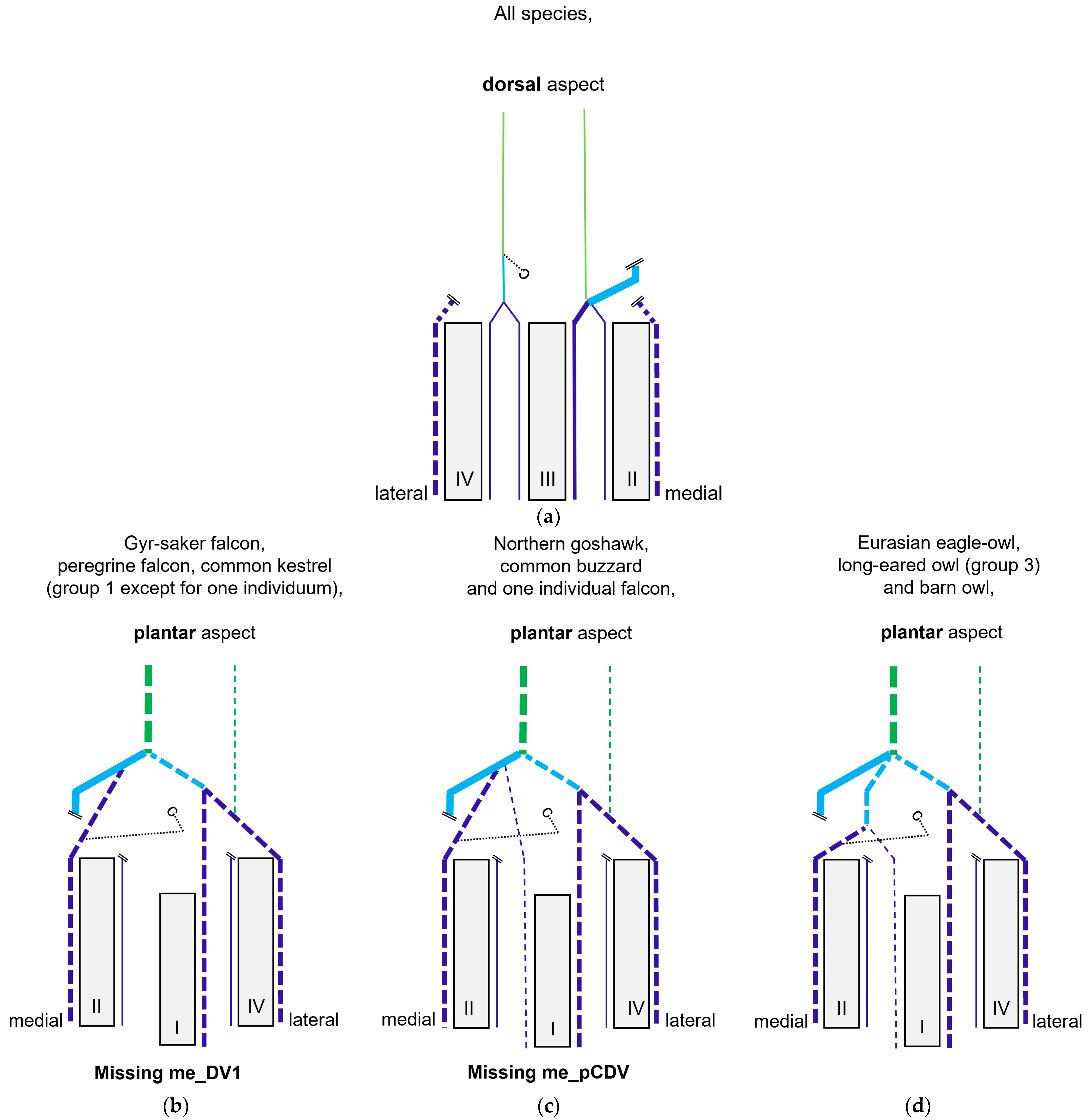
In
group 1, which included common kestrel, peregrine falcon, and gyr–saker falcon, the common dorsal metatarsal artery split far distally just proximal to the metatarsophalangeal joints. Thus, only one dorsal metatarsal artery, i.e., the common dorsal metatarsal artery, coursed along the tarsometatarsus. A similar course and splitting of the metatarsal arteries is shown in drawings of the pedal arteries of the common kestrel by Midtgård [
22] and of the peregrine falcon by Harcourt-Brown [
4,
23] as well as in a photograph of the southern caracara by Oliveira et al. [
24]. The caracara also belongs to the family of
Falconidae within the order
Falconiformes [
28]. This course was also described for the common kestrel in a previous study by Manno [
21], who simply called the common metatarsal artery the “arteria metatarsalis dorsalis” and the resulting arteries the “common digital arteries”, due to the distal splitting and their further ramification into proper digital arteries. However, the simultaneous use of the terms “metatarsal arteries” and “common digital arteries” compared to the nomenclature in mammals [
33] might suggest that there is a superficial and deep arterial system at the tarsometatarsus. However, this is not the case in birds, so we decided to use the term “metatarsal arteries” for both the common artery as well as the three arteries resulting from its splitting, which is consistent with the current avian nomenclature also used in the Nomina Anatomica Avium, e.g., [
22,
30,
32]. In his comparative study investigating 43 different avian species, Midtgård [
22] stated that the number of dorsal metatarsal arteries varied from one to three between species, and that in most bird species—including the common kestrel—just one single artery was present. However, he considered only large arteries in this assessment. Midtgård [
22] labeled the three dorsal metatarsal arteries
Aa. metatarsales dorsales I,
II, and
III from medial to lateral. He based this nomenclature on their original relation to the metatarsal bones before the second to fourth metatarsal bones fused into one tarsometatarsal bone during evolution [
34,
35]. According to Midtgård’s hypothetical pattern, it is the
A. metatarsalis dorsalis III that remained in avian species with only one dorsal metatarsal artery [
22]. However, in the domestic chicken [
29,
30] the terms lateral and medial dorsal metatarsal artery (
A. metatarsalis dorsalis lateralis and
medialis) were used for the two vessels arising from the splitting of the common dorsal metatarsal artery. The same terms were used in different species of waterfowl [
36,
37,
38]. As this nomenclature does not require any knowledge about evolutionary theories, but reflects merely the position of the vessel, it may be easier for veterinary practitioners to handle. Therefore, we favor these terms, and we adopted the terms medial and lateral dorsal metatarsal artery and introduced the term “middle dorsal metatarsal artery”, for we had three metatarsal arteries to label. In Latin, this middle artery would be the
A. metatarsalis dorsalis media.
Only in two species, in the domestic pigeon (
Columba livia domestica) and the short-eared owl (
Asio flammeus), did Midtgård [
22] find three parallel arteries running distally along the tarsometatarsus. The same vessel course was seen in Eurasian eagle–owls and long-eared owls (
group 3), which corresponds with Midtgård’s observations; these two owl species belong to the same family of
Strigidae within the order
Strigiformes as the short-eared owl [
28]. In the little owl (
Athena noctua), a proximal splitting in a medial and lateral metatarsal artery was reported by Manno [
21]. Only further distally, he described a third artery, which split off the medial metatarsal artery, and ran to the first toe. We assume that this artery can be equated with the medial metatarsal artery of the present work.
In the representatives of
group 2, i.e., northern goshawk, common buzzard, and barn owl, the common dorsal metatarsal artery split into two parallel arteries running along the tarsometatarsus, of which the medial one was much smaller than the lateral one. In a study on the northern goshawk, Wendt [
25] described the same arterial course, but called the medial dorsal artery
A. metatarsalis plantaris, a term which is normally used for plantar arteries beginning at the proximal vascular foramen. In contrast to this, studies on northern goshawk and peregrine falcon by Harcourt Brown [
4,
23] concluded that the blood vessel course of the goshawk was similar to that of the falcon, having only one dorsal metatarsal artery, the
A. metatarsalis III. This corresponds to the description presented by Midtgård [
22], who, however, only considered prominent arteries in his assessment. Although Midtgård [
22] basically mentioned a second slender dorsal metatarsal artery in some species showing only one main metatarsal artery, unfortunately he did not describe in which species it occurred. Thus, the only species in which Midtgård [
22] found two—equally thick—dorsal metatarsal arteries was the Humboldt penguin (
Spheniscus humboldti). After the splitting, the lateral artery descended distally and, in our opinion, it remained the common dorsal metatarsal artery in the representatives of group 2, because it split again into two arteries at the distal end of the tarsometatarsus. This branching pattern is in accordance with that one found in the common buzzard by Manno [
21], although the author used the term “common digital artery” instead of “metatarsal artery”. In the Eurasian sparrowhawk (
Accipiter nisus), however, the division into a medial and a lateral artery was far distal, after the distal perforating branch had already left [
21]. Similar to the findings in the birds of prey of group 2, the division of the common metatarsal artery into two metatarsal arteries was observed in both domestic chicken [
29,
30] and waterfowl [
36,
37,
38], with the medial one being much thinner than the lateral one.
The origin of the medial and lateral
plantar metatarsal arteries is stated to be the common dorsal metatarsal artery in most previous studies [
4,
29,
30,
31,
32,
36,
37,
38,
39]. Only a few authors [
22,
23], who did not describe a common metatarsal artery, named the cranial tibial artery as origin. As the origin of the arteries for the plantar side was located a short distance distal to the proximal articular surface of the tarsometatarsus, we consider the common dorsal metatarsal artery to be the origin. According to veterinary anatomy textbooks [
30,
31], the cranial tibial artery changes its name to common dorsal metatarsal artery as soon as it is located on the tarsometatarsus. We observed a common trunk for the two proximal intermetatarsal arteries passing to the plantar side in more than half of the feet examined. In the other feet two separate arteries arose, or only one artery was visible. These different manners seemed to be individually, not necessarily dependent on the species. In the fowl, both a common trunk for two arteries [
29,
30,
36] and the originating of two separate arteries [
31,
32,
37,
38] were described. In birds of prey, the origin of only one single artery [
21,
24,
25], two separate arteries [
4,
21,
23], and two arteries with a common trunk [
21] were mentioned.
In order to reach the plantar side, the intermetatarsal arteries ran through the two proximal vascular foramina of the tarsometatarsus in all eight avian species examined. The term “proximal vascular foramina” was also used by the majority of authors [
22,
23,
25,
30,
32,
36,
37,
38]. Some others referred to them as “interosseal foramina” [
4], or “intermetatarsal foramina” [
24,
29,
31], as the position of the proximal foramina may correspond to the interosseous spaces between the formerly separated metatarsal bones [
34,
35]. Similarly, the nomenclature of the two arteries passing through the vascular foramina varied. They were labeled as “plantar tarsal arteries” [
4,
23,
25,
31,
39], “plantar metatarsal arteries” [
29,
30], or “plantar intermetatarsal arteries” [
32,
36,
37,
38]. We consider a separate term for the arteries from their dorsal origin to their plantar proximal splitting, including the intraosseous part, would be useful for a clear description. However, both the terms “plantar tarsal arteries” and “plantar metatarsal arteries” are already used for the arteries at the plantar side of the tarsometatarsus [
22,
32]. “Plantar tarsal artery” is an appropriate name for a proximally running artery, because it supplies the plantar side of the tarsus. The same is true for the term “plantar metatarsal artery” for a distally running artery supplying the plantar side of the tarsometatarsus. The use of the term “plantar intermetatarsal arteries” (
Aa. intermetatarsales plantares) for the transversing arteries, listed among others in the Nomina Anatomica Avium [
32], is also confusing in our opinion, as we consider the term “plantar” inappropriate for a vessel that begins on the dorsal side of the foot. Additionally, there were not only the two proximal arteries that passed through the proximal vascular foramina, but also a distal artery that ran through the distal vascular foramen. Thus, in our opinion the terms “proximal intermetatarsal arteries” (
Aa. intermetatarsales proximales) and “distal intermetatarsal artery” (
A. intermetatarsalis distalis) would be appropriate as they clearly describe the course of the arteries and distinguish them from others.
Both plantar metatarsal arteries descending distally at the plantar side of the tarsometatarsus were present in seven of eight species examined; in one species, the common kestrel, only the lateral plantar metatarsal artery was demonstrated. With this observation, the question arises whether the common kestrel actually had no medial plantar metatarsal artery or whether this could not be depicted with our chosen methods due to its small size or was even due to individual variations. Manno [
21] reported for the common kestrel and the little owl two plantar metatarsal arteries each, which, however, terminated approximately on half of the tarsometatarsus. In studies on the southern caracara [
24], the northern goshawk [
4,
23,
25], and the peregrine falcon [
4,
23], no information about the further course of the plantar arteries along the tarsometatarsus were given at all. For the domestic chicken, two arteries were described, of which only the lateral one continued distally while the medial one stayed in the region of the tarsus [
29,
30]. However, in two comparative references [
22,
32], as well as in a study about the common buzzard and the sparrowhawk [
21] and in a study about the ostrich (
Struthio camelus) [
39], two metatarsal arteries were followed distally, the lateral artery being much more prominent than the medial one. Depending on the author, only the lateral [
22,
29,
30] or both [
21,
31,
36,
37,
38] descending metatarsal arteries finally joined the arterial plantar arch distally at the tarsometatarsus. We found that the lateral plantar metatarsal artery in (nearly) all specimens of all species examined connected with the plantar arch; this was true for the medial artery only for some specimens. Otherwise, the distal termination of the medial plantar metatarsal artery remained unclear in a couple of cases including all examined specimens of the long-eared owl.
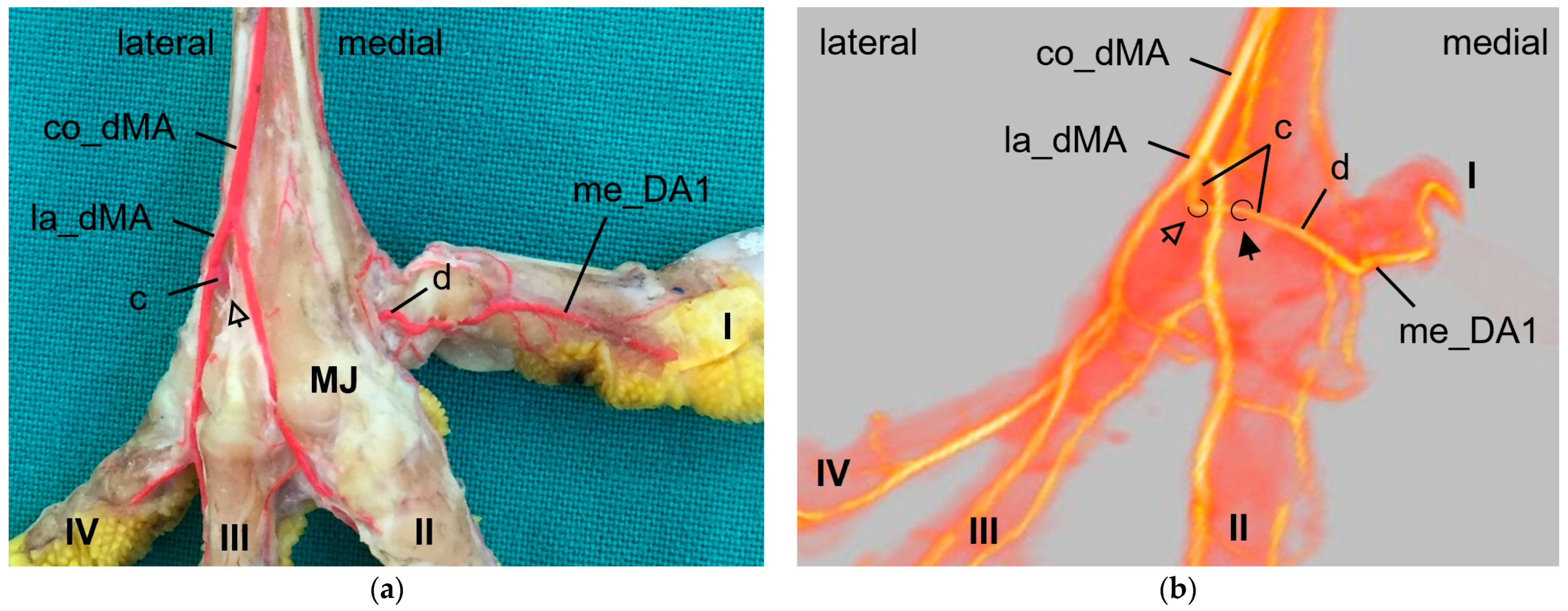
The
arterial plantar arch lay underneath the flexor tendons and was present distally at the tarsometatarsus in all species of birds of prey and owls examined, except for the long-eared owl. In the long-eared owl, we only discovered some vessels, which, however, did not form a clear arch. Here again the question arises whether the arch had not been filled due to its small size or was actually missing in the long-eared owl. When present, the arterial plantar arch originated from the artery that passed through the distal vascular foramen and that we therefore named “distal intermetatarsal artery” similar to the proximal transversing arteries, as mentioned above. The dorsal origin of the distal intermetatarsal artery was the lateral dorsal metatarsal artery in all eight species studied. This origin was also observed in some species of birds of prey and owls [
4,
22], in the domestic chicken [
29,
30], in different species of waterfowl [
36,
37,
38], and several other bird species [
22,
32], even though the artery was called the third dorsal metatarsal artery in the latter studies. However, in former studies on birds of prey, i.e., northern goshawk [
4,
23,
25] and peregrine falcon [
4,
23], the connection between the distal intermetatarsal artery and the arterial plantar arch was not described. In the present study, species-specific differences in the strength of the plantar arch were seen. In common buzzards, northern goshawks, and barn owls (group 2), the arch was much more prominent than in the falcon species (group 1) and in the Eurasian eagle–owl (group 3). These differences have not yet been described in species of birds of prey and owls, but have a decisive influence on the origin of the digital arteries of the first and partially of the second toe and might therefore be associated with circulatory-related pathological conditions of the feet.
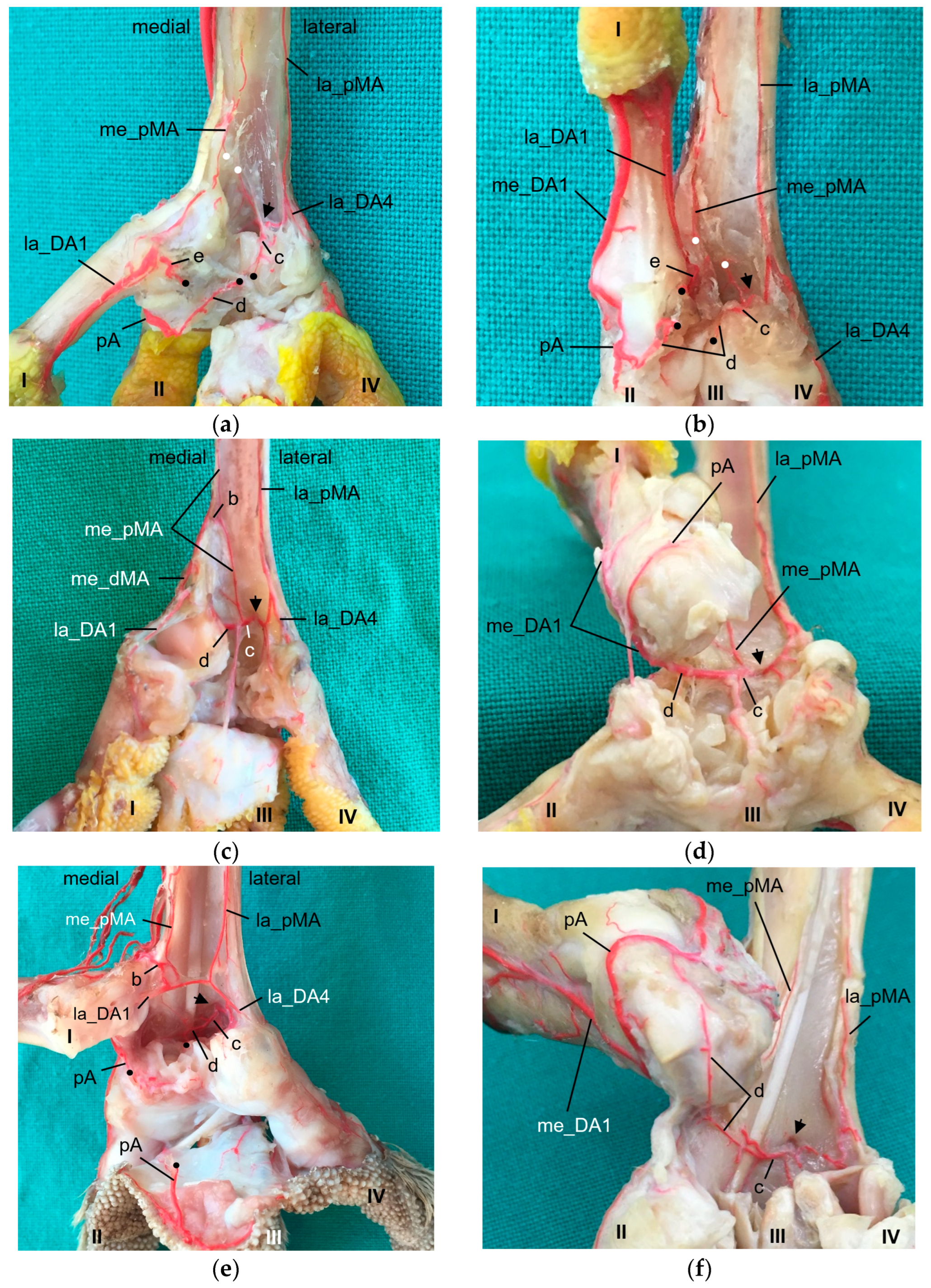
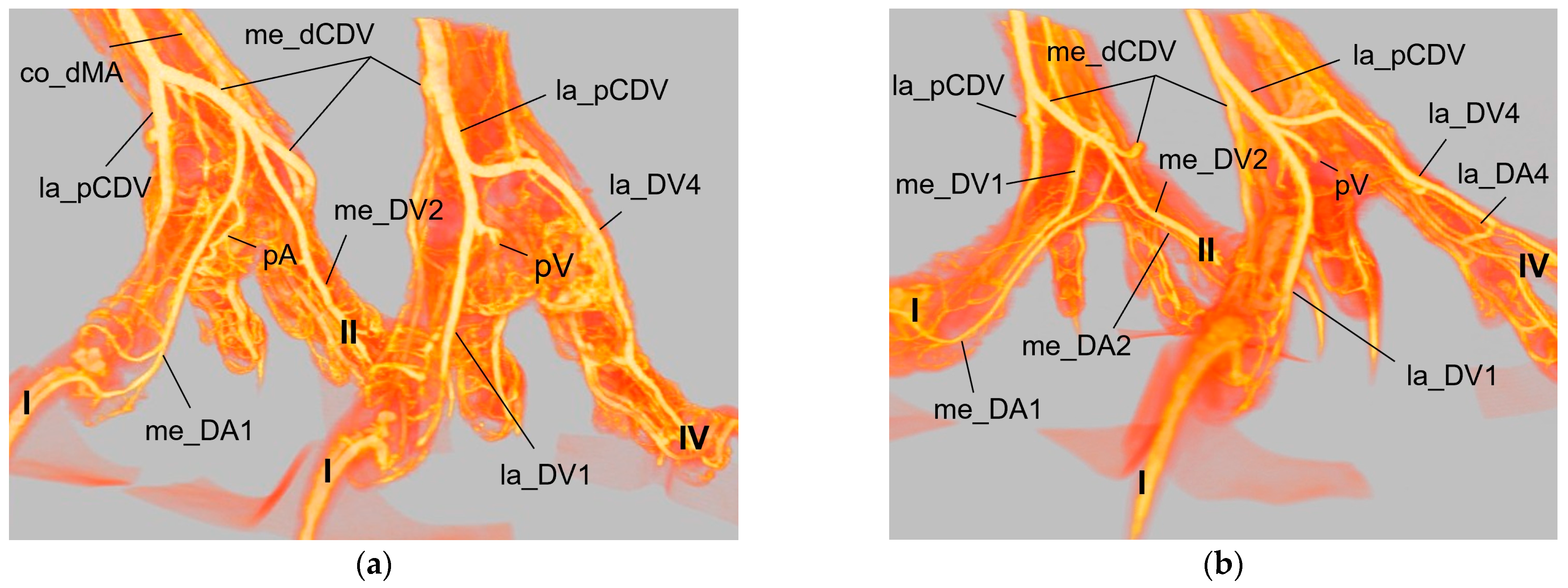
In general, in the present study, we discovered two proper
digital arteries per toe, one on the medial side and one on the lateral side. Exceptions were the gyr–saker falcon, which was missing the medial artery of toe II and the long-eared owl, which was missing the lateral artery of toe IV. Additionally, we noticed that in all toes the artery on one side (me_DA1, la_DA2, la_DA3, me_DA4) was more prominent than the artery on the other side, again in all species examined. This pattern was also shown in a comparative study in 43 species by Midtgård [
22] and adopted in the Nomina Anatomica Avium by Baumel [
32]. In northern goshawks and peregrine falcons, two arteries per toe were described but no differences in their size were stated by Harcourt-Brown [
4]. This is in accordance with findings in the domestic duck (
Anas platyrhynchos domesticus) [
36] and the domestic goose (
Anser anser domesticus) [
37,
38]; however, only one digital artery supplying the first toe was found in these species. In the southern caracara, belonging to the family
Falconidae, digital arteries were found medially on the first toe, and “axially” between the second and third as well as between the third and fourth toes [
24]. This is similar to the distribution pattern found in the domestic chicken, in which one artery per toe was present medially at toe I and IV as well as laterally at toe II and III, and, in addition, a small opposite artery on the medial side of toe III [
29,
30]. This resembles the location of the main digital arteries found in the present study, plus the small medial artery of toe III. Surprisingly, in the study by Vollmerhaus and Hegner [
29], an opposite artery was seen only on the third toe; on all other toes, small opposite arteries were found merely along the distal phalanges.
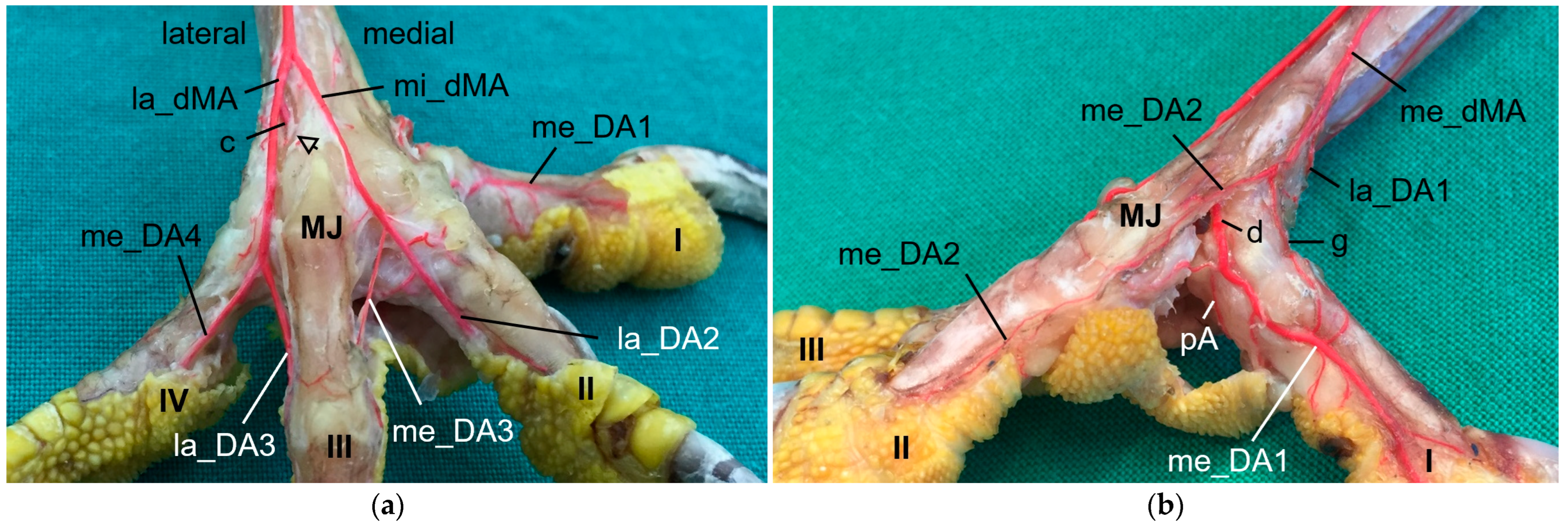
As mentioned above, the
medial artery of toe II and both arteries of the first toe (la_DA1, me_DA1) showed a species-specific origin. In particular, the origin of the medial artery of the first toe depended on the size of the plantar arterial arch. In common kestrels, peregrine falcons, gyr–saker falcons, Eurasian eagle–owls, and long-eared owls (groups 1 and 3), the strong medial artery of toe I was the direct prolongation of the medial dorsal metatarsal artery, which gave also rise to the small medial artery of the second toe. Thus, in these species the medial metatarsal artery was strong while the arterial plantar arch just continued as a very small branch that only anastomosed with the medial digital artery of the first toe via the arterial pulvinar branch. The small lateral artery of toe I branched off from the medial dorsal metatarsal artery in both owl species, whereas it originated from the medial plantar metatarsal artery in peregrine falcons and gyr–saker falcons. Its origin in the common kestrel remained unclear. In this context must be noted that both methods (latex and µCT) applied in the present study reached their limits especially with blood vessels of a very small diameter in smaller avian species, i.e., common kestrel, long-eared owl, and barn owl. In some cases, vessels were not completely filled with injection mass and thus, their origin or destination could not be identified clearly. This may be the reason why in several previous studies, the smaller arteries, i.e., the lateral artery of toe I and the medial artery of toe II, were not found [
21,
22,
24]. Although Manno [
21] and Midtgård [
22] used a different nomenclature, their representations depicting the large arteries showed the same origin for the me_DA1, la_DA1, and me_DA2 in falcon and owl species, as described by us. A drawing of the dorsal aspect of the pedal arteries of a peregrine falcon [
4] is in total accordance with our findings in the falcon species examined. Unfortunately, the legend contradicted the drawing, which stated that the medial side of the first—and also of the fourth—toe were supplied through the distal vascular foramen. This and the authors statement that no significant differences between northern goshawks and peregrine falcons were found [
4,
23] did not agree with our results. Oliveira et al. [
24] reported only that the common dorsal metatarsal artery gave off digital arteries, of which one ran medially along the first toe in the southern caracara. In contrast to species of groups 1 and 3, we were able to demonstrate a strong arterial plantar arch in northern goshawks, common buzzards, and barn owls (group 2), the largest part of which continued as the strong medial artery of toe I, while the medial dorsal metatarsal artery was quite small in these species. Our findings for species of group 2 were in accordance with the description for common buzzard and sparrowhawk [
21], European honey buzzard (
Pernis apivorus) [
20], and northern goshawk [
25]. In the study on the northern goshawk [
25], a photograph suggests that the strong medial artery of toe I also was joined by the small medial dorsal metatarsal artery, which is similar to that found in the present study.
On the lateral side of the foot, a small
lateral artery of toe IV was detected in all species examined, except for the long-eared owl. This digital artery was derived from the lateral plantar metatarsal artery. The same was also observed in the common buzzard and the sparrowhawk [
21], as well as in different species of waterfowl [
36,
37,
38], and was illustrated in a drawing showing the hypothetical original state of all metatarsal and digital arteries in birds [
22]. In peregrine falcons and northern goshawks, the origin was stated to be an arterial branch from the dorsal metatarsal artery that ran through the distal vascular foramen [
23], in this way, possibly describing the connection of the distal intermetatarsal artery via the plantar arch to the lateral artery of toe IV. Only in the little owl was a dorsal origin of the lateral artery of toe IV from the lateral metatarsal artery reported [
21].
In all species examined, the
medial artery of toe IV and the lateral one of toe III originated from the splitting of the lateral dorsal metatarsal artery, and the
small medial artery of toe III and large lateral one of toe II arose from the splitting of the middle dorsal metatarsal artery. The same splitting pattern was described using old nomenclature in the European honey buzzard [
20] and in a more recent study on the southern caracara [
24]. A drawing of the dorsal aspect of the pedal arteries of a peregrine falcon [
4] is also in accordance with our finding in the falcon species examined with respect to the splitting. The same is true for drawings of the main dorsal metatarsal arteries and their distribution pattern in the common kestrel and the short-eared owl [
22]. In the northern goshawk, a photograph suggests that the pattern is similar to that; unfortunately, Wendt [
25] did not explain the ramifications, but only mentioned the presence of digital arteries at the toes II to IV. Thus, the main arteries of toes IV, III, and II originated from dorsal metatarsal arteries in all birds of prey and owls examined in agreement to the available literature [
4,
20,
22,
24].
In early comparative studies on the blood supply of the avian pelvic limb, Barkow [
20], Manno [
21], and Hafferl [
40] already defined the origin of the digital arteries from the dorsal metatarsal artery or arteries to be a
dorsal supply of the toes. This was taken up again in a more recent study by Midtgård [
22]. Although digital arteries were described as prolongations of metatarsal arteries in handbooks [
30,
32], in the domestic chicken, only the lateral artery of toe III (la_DA3) and the medial artery of toe IV (med_DA4) were direct continuations of the lateral dorsal metatarsal artery; the large lateral artery of toe II (la_DA2) and the large medial artery of toe I (me_DA1) had their main origin from the arterial plantar arch [
22,
29,
30]. Only a small medial dorsal metatarsal artery also joined the lateral artery of toe II [
29,
30]. This pattern was defined to be a partly plantar supply [
22,
40]. In six different species of wild ducks, Midtgård [
22] showed that the lateral artery of toe III (la_DA3) and the medial artery of toe IV (med_DA4) had their origin also from the plantar arch. According to Midtgård [
22], an entirely
plantar supply was present when all toes were provided with arteries that originated from branches passing through the proximal or distal vascular foramina of the tarsometatarsus. However, studies on the domestic goose [
37,
38] and domestic duck [
36] observed that the lateral artery of toe II and the medial one of toe III branched off from the medial dorsal metatarsal artery. The latter one was not described in different species of the orders
Anseriformes by Midtgård [
22], as he only focused on the prominent metatarsal and digital arteries and used them for his classification into “entirely dorsal”, “entirely plantar”, and “partly plantar” supply. When all—small and prominent—metatarsal and digital arteries are taken into account, there is no entirely plantar or dorsal type of supply. Therefore, in our opinion, it would be more appropriate to speak of a mainly plantar or mainly dorsal supply. Summarizing, the present study revealed a mainly dorsal supply for common kestrel, peregrine falcon and gyr–saker falcon, Eurasian eagle–owl, and long-eared owl, and a predominantly or partly dorsal supply for northern goshawk, common buzzard, and barn owl.
Considering the results of the present study, it seems that taxonomically closely related species of birds of prey and owls show a similar course of blood vessels, as demonstrated for the course and branching of metatarsal and digital arteries in different species of the orders
Falconiformes and
Accipitriformes as well as the family
Strigidae. The barn owl occupied a special position as its arterial branching pattern resembled that of the
Accipitriformes more than that of the other owls, i.e., Eurasian eagle–owl and long-eared owl, belonging to the family of
Strigidae. This might be explained by the fact that the barn owl belongs to another family within the order Strigiformes, the
Tytonidae, which shows some morphological specialties in comparison to the
Strigidae [
28]. However, some even closely related species also showed specific differences in the pedal arterial supply, i.e., within the order
Charadriiformes or
Anseriiformes [
22]. Therefore, Midtgård [
22] suggested that the species-specific flexion or extension posture of the intertarsal joint in physiological movement makes either the dorsal or plantar course more convenient for the vessels, and thus different patterns have evolved in different species.
In the veins of the avian foot, a large number of venous valves occur [
29,
41]; unfortunately, these valves are a known problem in retrograde vein imaging, and are a possible cause of the incomplete retrograde filling of veins [
22]. Reasons for this might also be the clotting of blood in the vessels or unknown previous injuries. In alignment to Vollmerhaus and Hegner [
29], we observed that an increased (compared to arteries) and continuous manual pressure was necessary to overcome the venous valves and to adequately fill the veins retrogradely, especially with latex milk. Like Harcourt-Brown [
4] and Vollmerhaus and Hegner [
29], we found that, during the filling of the arteries, the injection material was able to pass orthogradely into veins via arteriovenous anastomoses in the distal phalanges of the toes. This effect was particularly evident when using the much less viscous X-ray contrast medium. The anastomoses in the distal phalanges were already described by Clara [
42], Vollmerhaus and Hegner [
29], and Schumacher [
41].
Similar to the arteries, every toe had two proper
digital veins—a lateral and a medial, one always stronger than the other one. The main digital vein lay always opposite the main digital artery in all species examined, which was seen especially well in the micro-computed tomography scans. This reciprocal asymmetry of digital arteries and veins was also observed by Midtgård [
22]. In contrast to their description of arteries, Vollmerhaus and Hegner [
29] found two veins per toe, with the vein not accompanied by an artery always stronger than the opposite vein. Midtgård [
22] suggested that hemodynamic effects during angiogenesis could be a possible reason for the development of the reciprocal asymmetry. Similar to the development of a more dorsal or more plantar arterial supply of the toes, hemodynamic effects might cause one pathway to become prominent while the other one recedes [
22].
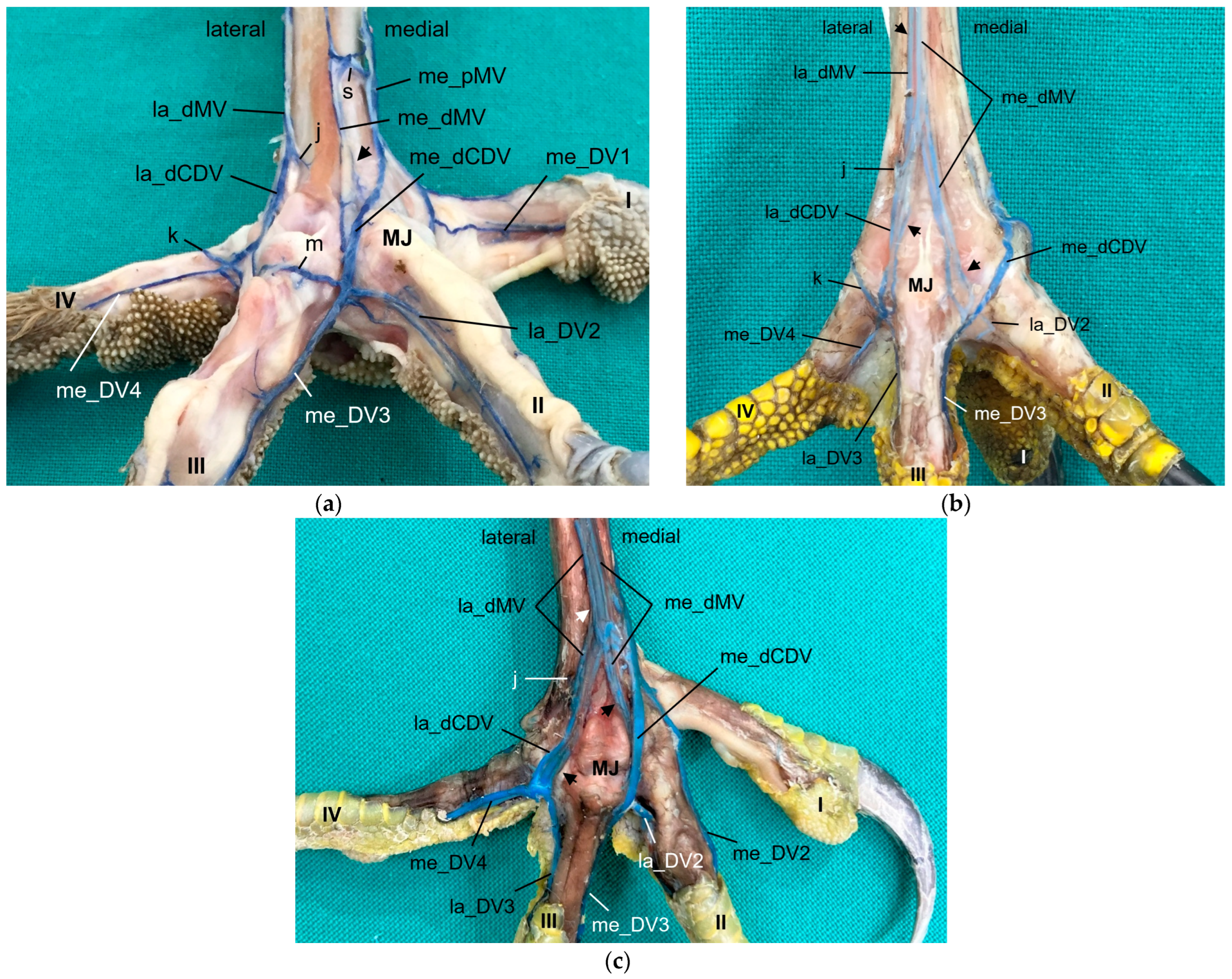
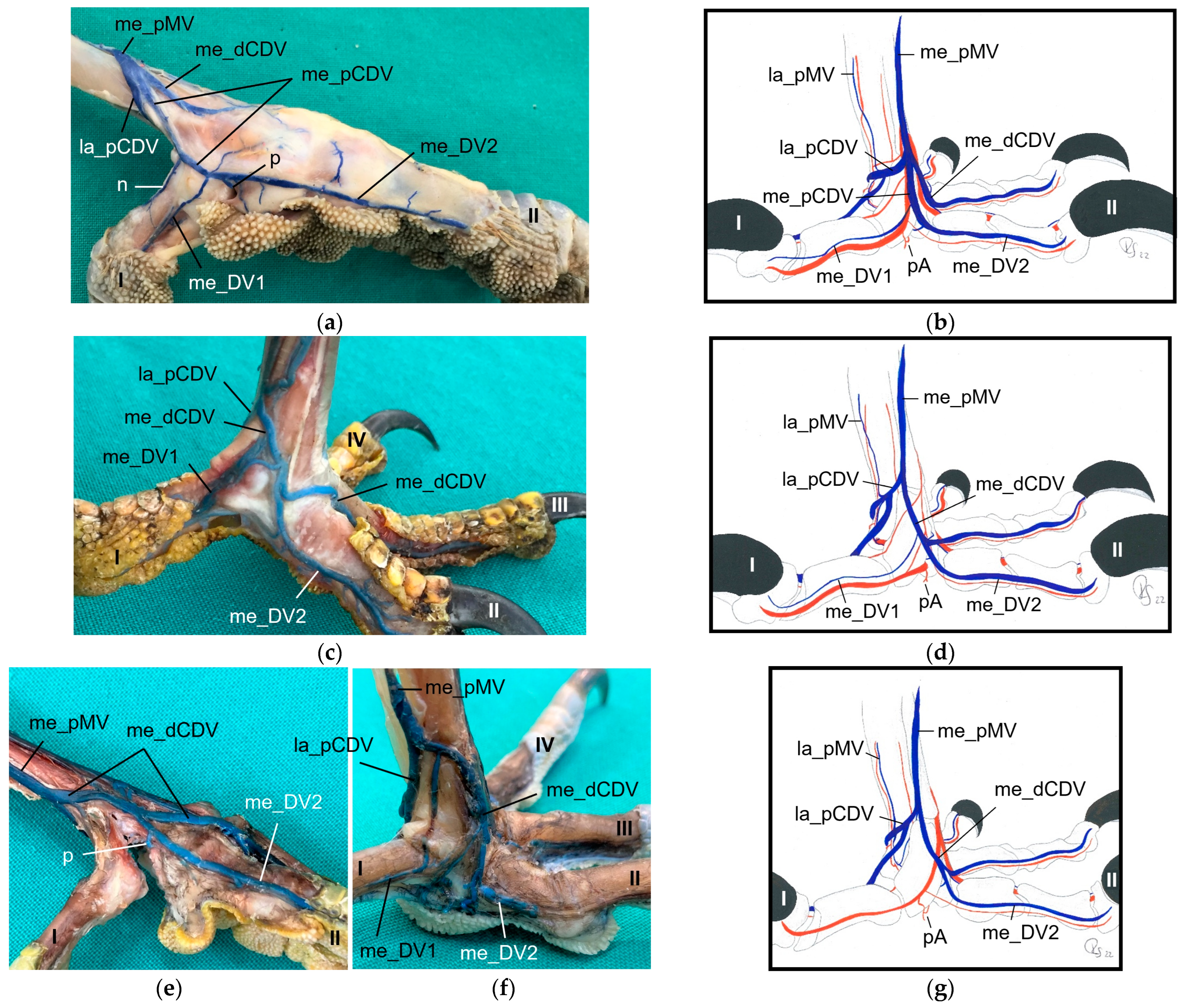
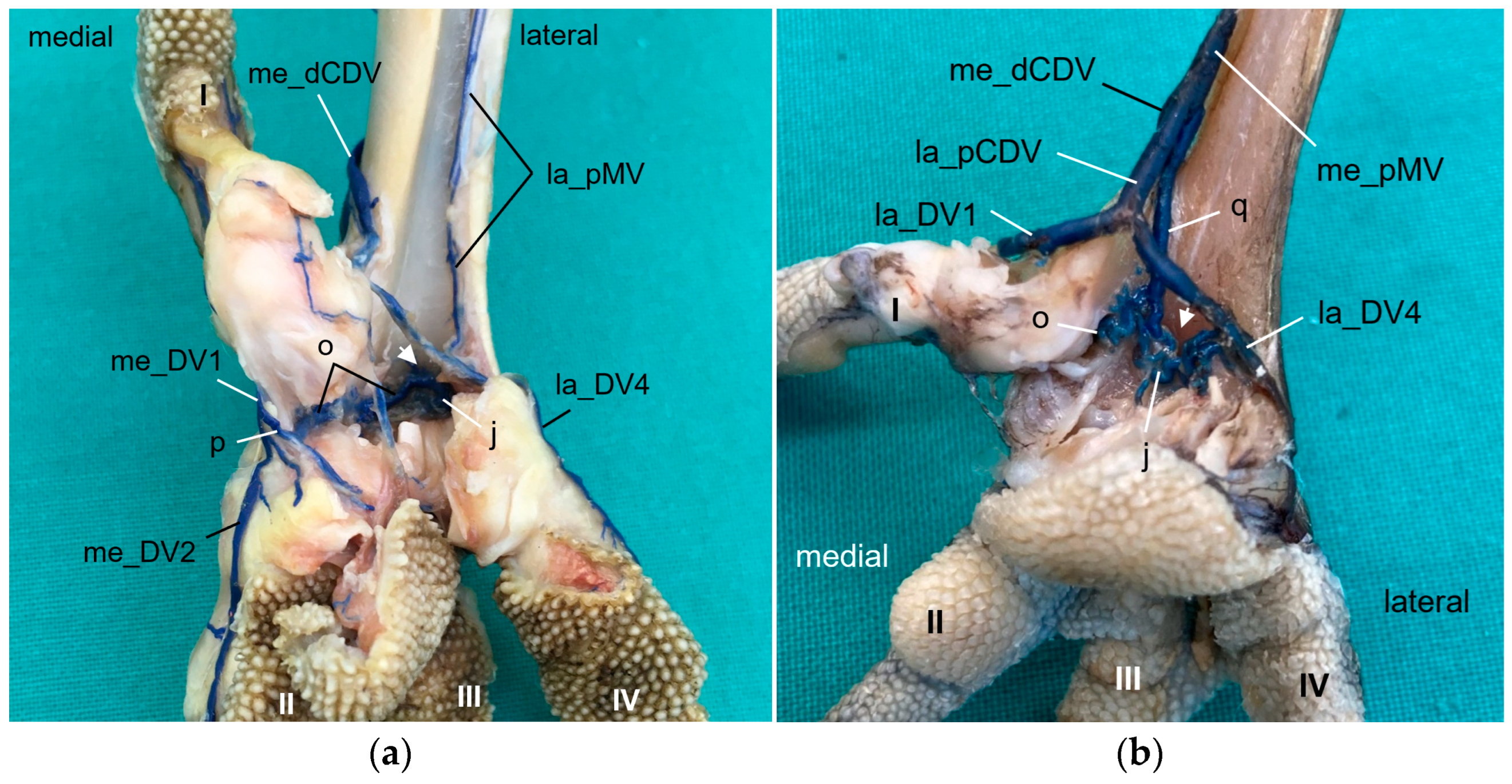
Looking at the pictures of the vein course in the Eurasian eagle–owl, it occurred to us that each of the two digital veins that ran next to each other united into one
common digital vein. Unlike the arteries, these common digital veins reunited in turn to form metatarsal veins. Therefore, we have introduced the term “common digital” for the veins only. This connection pattern of six out of eight proper digital veins had already been described by Vollmerhaus and Hegner [
29] for the domestic chicken, but no specific term had been given to the veins. In this way, a symmetric basic pattern of digital veins was created: four common digital veins, one lateral and one medial each on the dorsal and plantar sides of the foot. This pattern was perfectly visible not only in the Eurasian eagle-owl, but in all examined owl species including the barn owl. In northern goshawks and common buzzards as well as in one single specimen of the examined falcons, a variant of this pattern was seen, as the medial plantar common digital vein was missing. The two corresponding proper digital veins joined into the medial dorsal common digital vein separately instead. This observation is in accordance with a drawing showing the pedal veins of the medial side of a foot of a northern goshawk [
4], even if the nomenclature used is different to ours. In all falcons examined except one (see above), the medial digital vein of the first toe was missing. At this point we cannot tell whether the vein was just not filled or actually missing which would imply a difference in the venous branching pattern between species of the order
Accipitriformes and
Falconiformes. It should be noted that, also due to the variants in the branching pattern of the veins, the different species were grouped, but this did not completely coincide with the grouping based on the arterial distribution pattern. However, venous differences were only minor compared to arterial ones.
In order to join the lateral vein of toe I and to form the lateral plantar common digital vein, the strong lateral vein of toe IV curved medially superficial to the flexor tendons in all species examined. This arch-like venous course was shown in drawings made by Vollmerhaus and Hegner [
29] as well as Midtgård [
22]. However, this prolonged curved part of the lateral vein of toe IV was called “superficial venous plantar arch”, a term we omitted in the present study. Therefore, we would like to propose to keep the term “digital vein” for the arch-like part of the lateral vein of toe IV and to introduce the term “lateral plantar common digital vein”, after the connection with the lateral vein of toe I. We believe this would better reflect the symmetrical pattern of the venous system with four common digital veins. Until now, the superficial venous plantar arch was described to be the main plantar vein, into which the digital veins joined [
4,
22,
32]. Vollmerhaus and Hegner [
29] described the course of pedal veins in detail in the domestic chicken. They recognized the formation of common digital veins in three locations without using the term. Due to the fact that in the chicken the respective lateral vein of toe I and of toe IV joined separately into the medial superficial plantar metatarsal vein, Vollmerhaus and Hegner [
29] described the curved part of the lateral vein of toe IV as superficial venous plantar arch. In drawings from Midtgård [
22] of a gull and a raven, not only the la_DV4 entered into the superficial plantar arch; the med_DV3 or lat_DV1 did also.
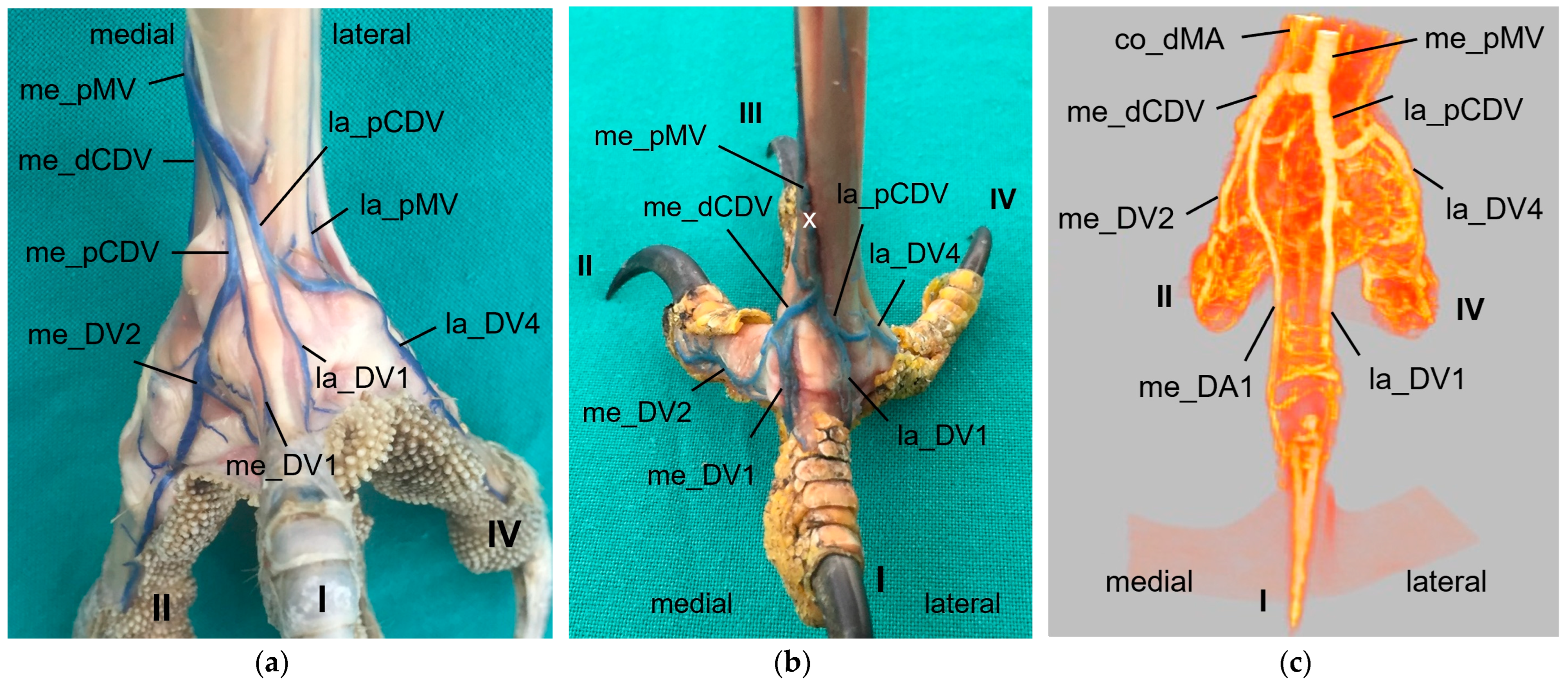
As mentioned above, the common digital veins reunited to form metatarsal veins. In the present study, we discovered four metatarsal veins in most species examined. On the dorsal side of the tarsometatarsus, the lateral dorsal common digital vein created by the union of the la_DV3 and me_DV4 became the
lateral dorsal metatarsal vein after the distal intermetatarsal vein was added. This branch passed through the distal vascular foramen and originated from the plantar venous arch deep under the flexor tendons in all species examined. This pattern was also observed in the domestic chicken [
29]. The
medial dorsal metatarsal vein originated from the medial dorsal common digital vein, at or only very shortly after the union of la_DV2 and me_DV3 in all avian species examined. While the origin of the medial dorsal metatarsal vein was consistent with the findings in the domestic chicken [
29], its further course differed. The medial dorsal metatarsal artery joined into the lateral one in the domestic chicken creating a common dorsal metatarsal vein that continued proximally [
29]. The latter was also mentioned in the northern goshawk [
25]. In contrast to this description, the medial dorsal metatarsal vein ran separately proximally towards the proximal vascular foramina in all species examined in the present study. The different findings might be due to several venovenous anastomoses between the two dorsal metatarsal veins, which might suggest a confluence of the two veins. Additionally, various venovenous anastomoses to the large medial plantar metatarsal vein were observed in the species examined, which is in agreement with results in the domestic chicken [
29]. These anastomoses may explain why, in two studies [
25,
29], the common dorsal metatarsal vein was reported to continue proximally along the tibiotarsus as cranial tibial vein. However, Baumel [
32] stated that the common dorsal metatarsal vein joined the medial superficial plantar metatarsal vein in the domestic chicken. Thus, differences in the course of metatarsal veins between studies or species mentioned above might reflect actual differences but could also be associated with this frequently cross-linked metatarsal venous system leading to different interpretations by the authors. In accordance with the findings of Vollmerhaus and Hegner [
29], we found a high number of venous collaterals of the dorsal metatarsal veins. The destination of the medial and lateral dorsal metatarsal veins remained unclear in the present study, because the area proximal to the tarsus was not investigated further. In the literature, a venous rete in the dorsal region of the intertarsal joint was reported, similar to the (tibio)-tarsal arterial rete. The venous rete was formed by collaterals of the dorsal veins and was also named
rete mirabile [
29,
31,
32].
On the plantar side of the tarsometatarsus, we discovered two metatarsal veins, the large medial plantar metatarsal vein and the small lateral plantar metatarsal vein, in almost all species studied. This observation is in line with other reports [
22,
29,
32]. Exceptions of this pattern were seen in long-eared owls, gyr–saker falcons, and common kestrels, in which the
lateral plantar metatarsal vein was not present. Here again, the question arises whether the vein was actually absent or whether it was just not depicted because of its small size. In all species examined in which the lateral plantar metatarsal vein was present, the origin of it was the prolonged and curved part of the lateral vein of toe IV. Previously, this part of the vein was called the superficial plantar venous arch, and described as the origin of the lateral metatarsal vein [
4,
22,
29,
32]. If present, the lateral plantar metatarsal vein continued proximally underneath the flexor tendons, passed through the lateral proximal vascular foramen as lateral proximal intermetatarsal vein and joined the common dorsal metatarsal vein on the dorsal side. The same course was described for the domestic chicken, but the vein was named the “
deep plantar metatarsal vein” [
29]. This was due to the observation that a second lateral plantar metatarsal vein was found on the lateral side of the tarsometatarsus in the domestic chicken [
29]. The authors described this superficial vein as a thin cutaneous vein. According to Vollmerhaus and Hegner [
29], in the domestic chicken, this vein also arose from the “superficial plantar arch”. We did not find this superficial lateral plantar metatarsal vein in any of the specimens examined. Therefore, we could omit the termini “superficial” and “deep” because there was only one plantar metatarsal vein on the lateral side. However, according to Baumel [
32], Harcourt-Brown [
4], and Midtgård [
22], the lateral plantar metatarsal vein joined the caudal tibial vein proximal to the intertarsal joint, and a vein passing through the proximal foramen of the tarsometatarsus was not mentioned. This indicates that these authors might describe the same superficial vein as Vollmerhaus and Hegner [
29].
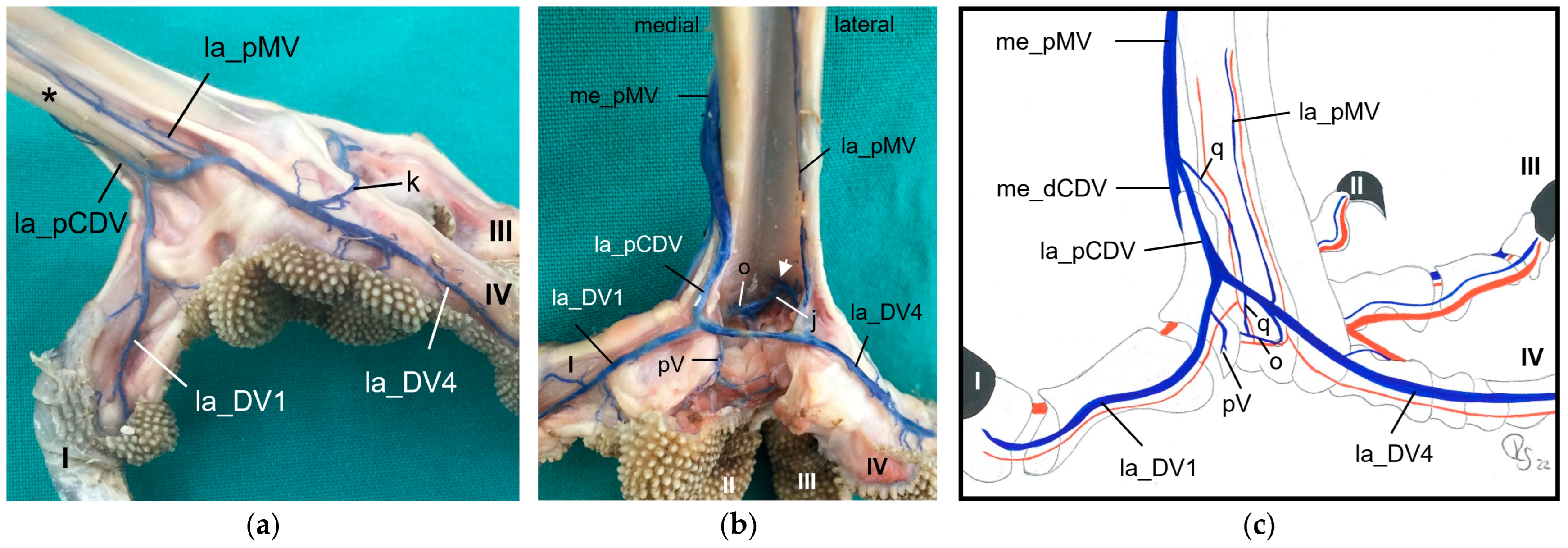
On the medial side of the tarsometatarsus, one very large
medial plantar metatarsal vein was present in all avian species studied. This vein was formed by the union of at least two common digital veins, i.e., the medial dorsal and the lateral plantar common digital vein. In owls, the medial plantar common digital vein was added to that union. The description and the drawings in the majority of previous publications agreed with our findings. However, they described that after the major digital veins had joined into a superficial plantar arch; this arch continued proximally as the medial superficial plantar metatarsal artery [
4,
22,
29,
32].
We also found a
venous plantar arch deep underneath the flexor tendons, which created a connection from a medial digital vein (toe I or toe II), except for long-eared owl and common kestrel, to the distal intermetatarsal vein, which passed through the distal vascular foramen. From this foramen, the arch also connected to the lateral vein of toe IV. Only in some gyr–saker falcons and peregrine falcons did we observe a strong additional vein connecting the deep venous arch to the me_dCDV and the me_pMV. In the domestic chicken, the deep plantar arch also had a medial connection to one of the medial common veins joining into the medial superficial plantar metatarsal vein and a lateral connection to the venous branch passing through the intermetatarsal distal foramen [
29]. In the domestic chicken, the deep plantar arch additionally received interdigital veins from the common veins of la_DV2 and me_DV3 and of la_DV3 and me_DV4, respectively [
29]. However, the connection to the curved lateral vein of toe IV in the superficial plantar arch, which we found in the birds of prey and owls investigated in the present study, was not described in the domestic chicken. Former studies on birds of prey and owls did not mention a deep venous arch at all [
4,
22,
25].
The medial plantar metatarsal vein provided the main drainage of the foot and continued proximally as caudal tibial vein after passing the intertarsal joint, which was in agreement with previous studies [
22,
25,
29,
30,
31,
32].


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}