
Characterization of Extraintestinal Pathogenic Escherichia coli Strains Causing Canine Pneumonia in China: Antibiotic Resistance, Virulence Genes, and Sequence Typing
 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Main Reagents and Main Instruments
2.2. Animals
2.3. Experimental Methods
2.3.1. Case History Investigation
2.3.2. Isolation, Cultivation, and Identification of Pathogens
2.3.3. Pathological Autopsy Observations
2.3.4. Bacterial Whole-Genome Sequencing
2.3.5. Bacterial Antimicrobial Phenotype and Resistance Genes
2.3.6. Virulence Gene Detection
2.3.7. MLST
2.3.8. Detection of Escherichia coli and CNF-I Virulence Gene in Drinking Water Sources
3. Results
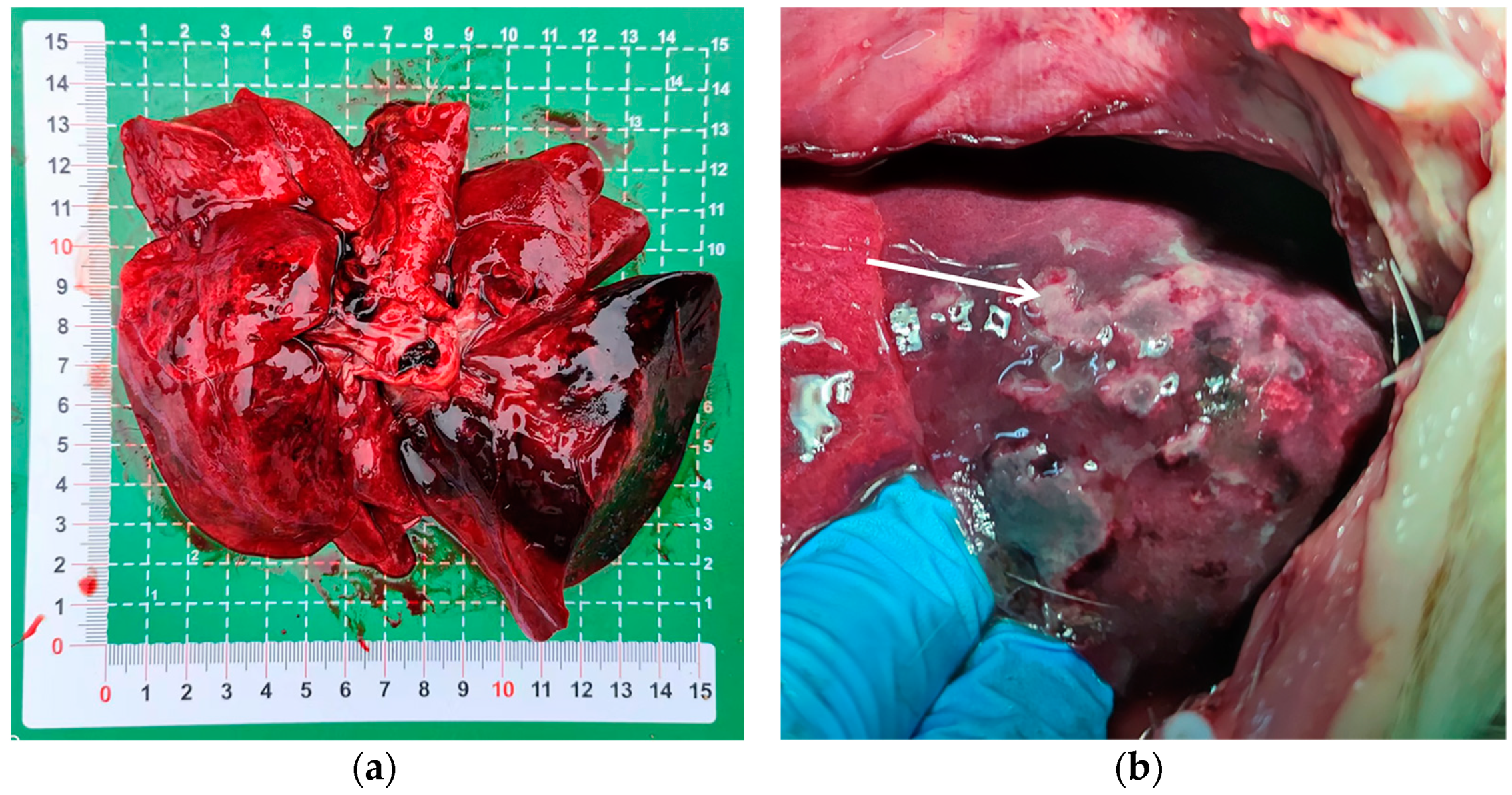
3.1. Identification of the Pathogen
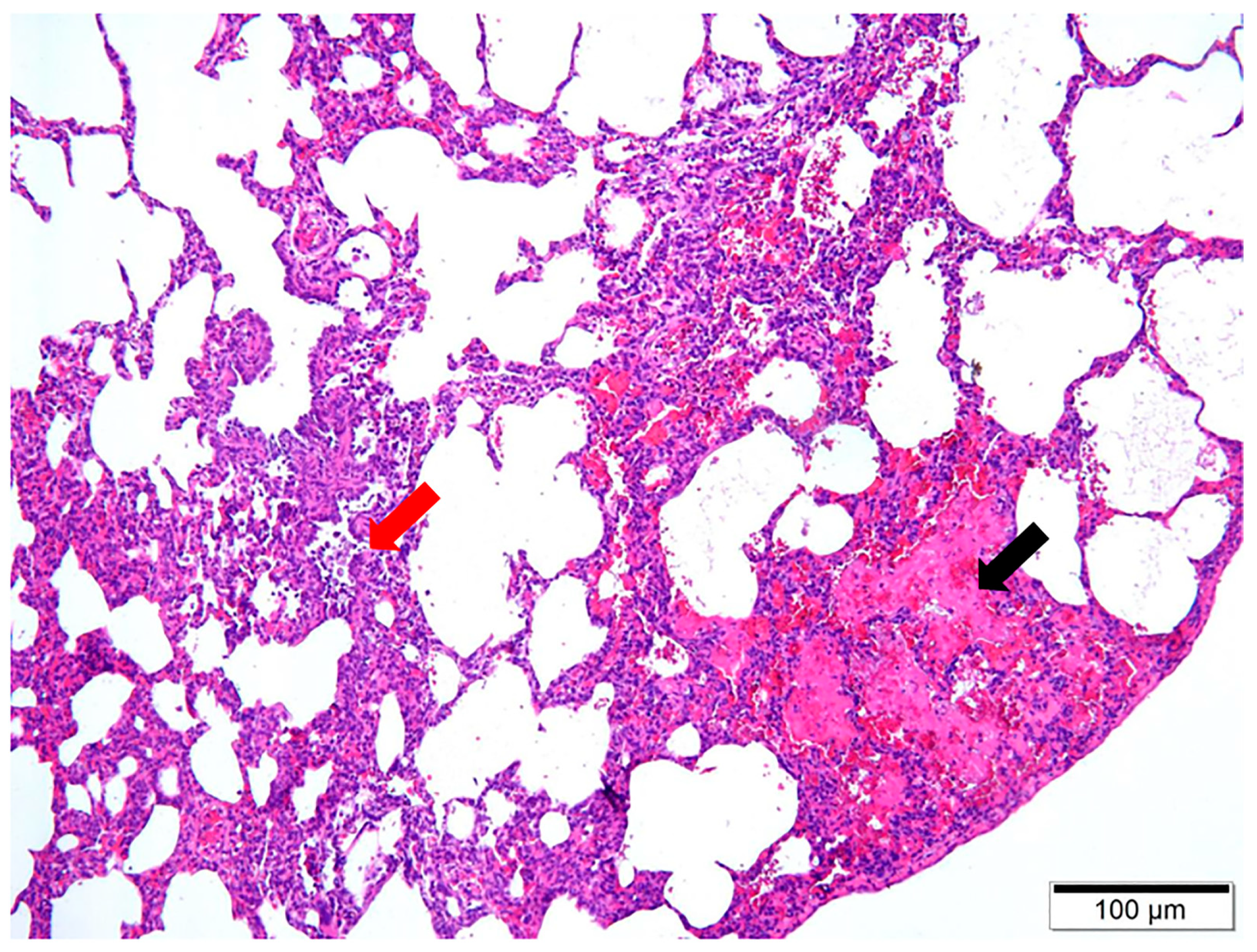
3.2. Pulmonary Histology
3.3. Evaluation of Whole-Genome Sequencing Data Quality for Bacteria
3.4. Antibiotic Susceptibility Testing
3.5. Detection of Virulence Genes
3.6. MLST
3.7. Detection of Escherichia coli and CNF-I Virulence Gene in Drinking Water
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ → 3′) | Length (bp) |
|---|---|---|
| 16S rRNA-F | AGAGTTTGATCCTGGCTCAG | 1500 |
| 16S rRNA-R | TACGGYTACCTTGTTACGACTT | |
| hlyF-F | GGCCACAGTCGTTTAGGGTGCTTACC | 450 |
| hlyF-R | GGCGGTTTAGGCATTCCGATACTCAG | |
| fyuA-F | TGATTAACCCCGCGACGGGAA | 787 |
| fyuA-R | CGCAGTAGGCACGATGTTGTA | |
| fimC-F | GGGTAGAAAATGCCGATGGTG | 497 |
| fimC-R | CGTAATTTTGGGGGTAAGTGC | |
| papC-F | GTGGCAGTATGAGTAATGACCGTTA | 296 |
| papC-R | ATATCCTTTCTGCAGGGATGCAATA | |
| iuc D-F | ACAAAAAGTTCTATCGCTTCC | 714 |
| iuc D-R | CCTGATCCAGATGATGCTC | |
| ompA-F | AGCTATCGCGATTGCAGTG | 919 |
| ompA-R | GGTGTTGCCAGTAACCGG | |
| fimH-F | CTGGTCATTCGCCTGTAAAACCGCCA | 878 |
| fimH-R | GTCACGCCAATAATCGATTGCACATTCCCT | |
| CNF-I-F | GAACTTATTAAGGATAGT | 543 |
| CNF-I-R | CATTATTTATAACGCTG | |
| cva/cvi-F | TCCAAGCGGACCCCTTATAG | 598 |
| cva/cvi-R | CGCAGCATAGTTCCATGCT | |
| vat-F | TCCATGCTTCAACGTCTCAGAG | 939 |
| vat-R | CTGTTGTCAGTGTCGTGAACG | |
| irp2-F | AAGGATTCGCTGTTACCGGA | 301 |
| irp2-R | TCGGCCAGGATGATTCGTCG | |
| iss-F | ATCACATAGGATTCTGCCG | 309 |
| iss-R | CAGCGGAGTATAGATGCCA | |
| iroN-F | AAGTCAAAGCAGGGGTTGCCCG | 667 |
| iroN-R | GACGCCGACATTAAGACGCAG | |
| tsh-F | ATAGGATGACAGGCTACCGAC | 598 |
| tsh-R | GTCTGTCAGACGTCTGTGTTTC | |
| STa-F | GGCTGGACATCATGGGAACTGG | 302 |
| STa-R | CGTCGGGAACGGGTAGAATCG | |
| STb-F | GAACGGCGGACTGTTAAT | 1391 |
| STb-R | TTGAACATTCAGAGTACCGGG | |
| K99-F | TAGCGATGTTAGAATAGGA | 314 |
| K99-R | AGATGGGATTACATTTAGG | |
| eae-F | ATGCTGGCATTTGGTCAGGTCGG | 233 |
| eae-R | TGACTCATGCCAGCCGCTCATGCG | |
| adk-F | ATTCTGCTTGGCGCTCCGGG | 583 |
| adk-R | CCGTCAACTTTCGCGTATTT | |
| fumC-F | TCACAGGTCGCCAGCGCTTC | 806 |
| fumC-R | GTACGCAGCGAAAAAGATTC | |
| gyrB-F | TCGGCGACACGGATGACGGC | 880 |
| gyrB-R | ATCAGGCCTTCACGCGCATC | |
| icd-F | ATGGAAAGTAAAGTAGTTGTTCCGGCACA | 878 |
| icd-R | GGACGCAGCAGGATCTGTT | |
| mdh-F | TTAACGAACTCCTGCCCCAGAGCGATATCTTTCTT | 932 |
| mdh-R | ATGAAAGTCGCAGTCCTCGGCGCTGCTGGCGG | |
| purA-F | CGCGCTGATGAAAGAGATGA | 816 |
| purA-R | CATACGGTAAGCCACGCAGA | |
| recA-F | CGCATTCGCTTTACCCTGACC | 780 |
| recA-R | TCTCGATCAGCTTCTCTTTT | |
| E.coli-F | GCGGTTTGTTAAGTCAGATGTGAA | 416 |
| E.coli-R | TGGATGTCAAGACCAGGTAAGG |
References
- Dale, A.P.; Woodford, N. Extra-intestinal pathogenic Escherichia coli (ExPEC): Disease, carriage and clones. J. Infect. 2015, 71, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Breitschwerdt, E.B.; DebRoy, C.; Mexas, A.M.; Brown, T.T.; Remick, A.K. Isolation of necrotoxigenic Escherichia coli from a dog with hemorrhagic pneumonia. J. Am. Vet. Med. Assoc. 2005, 226, 2016–2019. [Google Scholar] [CrossRef] [PubMed]
- Coe, S.E.; Magagna, M.A.; Zimmerman, A.; George, A.; Carter, C.; Dean, C.; Nelson, K. Extraintestinal Pathogenic Escherichia coli Causes Necrohemorrhagic Pneumonia in Multiple Research Dogs. Toxicol. Pathol. 2022, 50, 904–909. [Google Scholar] [CrossRef]
- Yu, Y.; Hu, B.; Fan, H.; Zhang, H.; Lian, S.; Li, H.; Li, S.; Yan, X.; Wang, S.; Bai, X. Molecular Epidemiology of Extraintestinal Pathogenic Escherichia coli Causing Hemorrhagic Pneumonia in Mink in Northern China. Front. Cell Infect. Microbiol. 2021, 11, 781068. [Google Scholar] [CrossRef]
- Huang, T.M.; Lin, T.L.; Wu, C.C. Antimicrobial susceptibility and resistance of chicken Escherichia coli, Salmonella spp., and Pasteurella multocida isolates. Avian Dis. 2009, 53, 89–93. [Google Scholar] [CrossRef]
- Farizqi, M.T.I.; Effendi, M.H.; Adikara, R.T.S.; Yudaniayanti, I.S.; Putra, G.D.S.; Khairullah, A.R.; Kurniawan, S.C.; Silaen, O.S.M.; Ramadhani, S.; Millannia, S.K.; et al. Detection of extended-spectrum β-lactamase-producing Escherichia coli genes isolated from cat rectal swabs at Surabaya Veterinary Hospital, Indonesia. Vet. World 2023, 16, 1917–1925. [Google Scholar] [CrossRef]
- Sukrama, I.D.M.; Franciska, J.; Suardana, I.W. Evaluation of the bacteriocin produced by strain 9 lactic acid bacteria isolate for biopreservation. Vet. World. 2020, 13, 2012–2019. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 7th ed.; CLSI Supplement VET01S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2024. [Google Scholar]
- Bin Kim, Y.; Yoon, M.Y.; Ha, J.S.; Seo, K.W.; Noh, E.B.; Son, S.H.; Lee, Y.J. Molecular characterization of avian pathogenic Escherichia coli from broiler chickens with colibacillosis. Poult. Sci. 2020, 99, 1088–1095. [Google Scholar] [CrossRef]
- Ramírez-Castillo, F.Y.; Loera-Muro, A.; Vargas-Padilla, N.D.; Moreno-Flores, A.C.; Avelar-González, F.J.; Harel, J.; Jacques, M.; Oropeza, R.; Barajas-García, C.C.; Guerrero-Barrera, A.L. Incorporation of Actinobacillus pleuropneumoniae in Preformed Biofilms by Escherichia coli Isolated from Drinking Water of Swine Farms. Front. Vet. Sci. 2018, 5, 184. [Google Scholar] [CrossRef]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum β-lactamases (ESBLs) in the developed world. J. Travel. Med. 2017, 24 (Suppl. S1), S44–S51. [Google Scholar] [CrossRef]
- Sun, L.; Meng, N.; Wang, Z.; Hong, J.; Dai, Y.; Wang, Z.; Wang, J.; Jiao, X. Genomic Characterization of ESBL/AmpC-Producing Escherichia coli in Stray Dogs Sheltered in Yangzhou, China. Infect. Drug Resist. 2022, 15, 7741–7750. [Google Scholar] [CrossRef] [PubMed]
- Bortolami, A.; Zendri, F.; Maciuca, E.I.; Wattret, A.; Ellis, C.; Schmidt, V.; Pinchbeck, G.; Timofte, D. Diversity, Virulence, and Clinical Significance of Extended-Spectrum β-Lactamase- and pAmpC-Producing Escherichia coli from Companion Animals. Front. Microbiol. 2019, 10, 1260. [Google Scholar] [CrossRef] [PubMed]
- Tamang, M.D.; Nam, H.M.; Jang, G.C.; Kim, S.R.; Chae, M.H.; Jung, S.C.; Byun, J.-W.; Park, Y.H.; Lim, S.-K. Molecular characterization of extended-spectrum-β-lactamase-producing and plasmid-mediated AmpC β-lactamase-producing Escherichia coli isolated from stray dogs in South Korea. Antimicrob. Agents Chemother. 2012, 56, 2705–2712. [Google Scholar] [CrossRef]
- Shin, S.R.; Noh, S.M.; Jung, W.K.; Shin, S.; Park, Y.K.; Moon, D.C.; Lim, S.-K.; Park, Y.H.; Park, K.T. Characterization of Extended-Spectrum β-Lactamase-Producing and AmpC β-Lactamase-Producing Enterobacterales Isolated from Companion Animals in Korea. Antibiotics 2021, 10, 249. [Google Scholar] [CrossRef]
- Corbella, M.; Caltagirone, M.; Gaiarsa, S.; Mariani, B.; Sassera, D.; Bitar, I.; Muzzi, A.; Migliavacca, R.; Scudeller, L.; Stronati, M. Characterization of an Outbreak of Extended-Spectrum β-Lactamase-Producing Klebsiella pneumoniae in a Neonatal Intensive Care Unit in Italy. Microb. Drug Resist. 2018, 24, 1128–1136. [Google Scholar] [CrossRef]
- Liu, H.; Liu, B.; Zheng, F.; Chen, X.; Ye, L.; He, Y. Distribution of pathogenic bacteria in lower respiratory tract infection in lung cancer patients after chemotherapy and analysis of integron resistance genes in respiratory tract isolates of uninfected patients. J. Thorac. Dis. 2020, 12, 4216–4223. [Google Scholar] [CrossRef]
- Kuhnert, P.; Boerlin, P.; Frey, J. Target genes for virulence assessment of Escherichia coli isolates from water, food and the environment. FEMS Microbiol. Rev. 2000, 24, 107–117. [Google Scholar] [CrossRef]
- Fabbri, A.; Travaglione, S.; Fiorentini, C. Escherichia coli cytotoxic necrotizing factor 1 (CNF1): Toxin biology, in vivo applications and therapeutic potential. Toxins 2010, 2, 283–296. [Google Scholar] [CrossRef]
- Moore, M.E.G.; Paulin-Curlee, G.; Johnston, B.D.; Clabots, C.; DebRoy, C.; Johnson, T.J.; Weber, B.; Porter, S.; Armién, A.G.; Johnson, J.R. Molecular Characteristics, Ecology, and Zoonotic Potential of Escherichia coli Strains That Cause Hemorrhagic Pneumonia in Animals. Appl. Environ. Microbiol. 2021, 87, e0147121. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.S.; Moon, B.Y.; Hwang, M.H.; Lee, S.K.; Ku, B.K.; Lee, K. Characterization of the pathogenicity of extraintestinal pathogenic Escherichia coli isolates from pneumonia-infected lung samples of dogs and cats in South Korea. Sci. Rep. 2023, 13, 5575. [Google Scholar] [CrossRef] [PubMed]
- Riley, L.W. Distinguishing Pathovars from Nonpathovars: Escherichia coli. Microbiol. Spectr. 2020, 8, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Maiden, M.C.J.; Bygraves, J.A.; Feil, E.; Morelli, G.; Russell, J.E.; Urwin, R.; Zhang, Q.; Zhou, J.; Zurth, K.; Caugant, D.A.; et al. Multilocus sequence typing: A portable approach to the identification of clones within populations of pathogenic microorganisms. Proc. Natl. Acad. Sci. USA 1998, 95, 3140–3145. [Google Scholar] [CrossRef]
- Nemoy, L.L.; Kotetishvili, M.; Tigno, J.; Keefer-Norris, A.; Harris, A.D.; Perencevich, E.N.; Johnson, J.A.; Torpey, D.; Sulakvelidze, A.; Morris, J.G.; et al. Multilocus sequence typing versus pulsed-field gel electrophoresis for characterization of extended-spectrum beta-lactamase-producing Escherichia coli isolates. J. Clin. Microbiol. 2005, 43, 1776–1781. [Google Scholar] [CrossRef]
- Maiden, M.C.J.; van Rensburg, M.J.J.; Bray, J.E.; Earle, S.G.; Ford, S.A.; Jolley, K.A.; McCarthy, N.D. MLST revisited: The gene-by-gene approach to bacterial genomics. Nat. Rev. Microbiol. 2013, 11, 728–736. [Google Scholar] [CrossRef]
- Dautzenberg, M.J.; Haverkate, M.R.; Bonten, M.J.; Bootsma, M.C. Epidemic potential of Escherichia coli ST131 and Klebsiella pneumoniae ST258: A systematic review and meta-analysis. BMJ Open 2016, 6, e009971. [Google Scholar] [CrossRef]
- Kondo, T.; Sakamoto, K.; Morinaga, Y.; Miyata, Y.; Yanagihara, K.; Sakai, H. Escherichia coli ST131 isolated from urological patients can acquire plasmid-mediated extended spectrum β-lactamase from other bacteria with high frequency. Int. J. Urol. 2022, 29, 587–594. [Google Scholar] [CrossRef]
- Kidsley, A.K.; O’dea, M.; Saputra, S.; Jordan, D.; Johnson, J.R.; Gordon, D.M.; Turni, C.; Djordjevic, S.P.; Abraham, S.; Trott, D.J. Genomic analysis of phylogenetic group B2 extraintestinal pathogenic E. coli causing infections in dogs in Australia. Vet. Microbiol. 2020, 248, 108783. [Google Scholar] [CrossRef]
- Li, D.; Wyrsch, E.R.; Elankumaran, P.; Dolejska, M.; Marenda, M.S.; Browning, G.F.; Bushell, R.N.; McKinnon, J.; Chowdhury, P.R.; Hitchick, N.; et al. Genomic comparisons of Escherichia coli ST131 from Australia. Microb. Genom. 2021, 7, 000721. [Google Scholar] [CrossRef]
- Zurita, J.; Sevillano, G.; Miño, A.P.Y.; Haro, N.; Larrea-Álvarez, M.; Alcocer, I.; Ortega-Paredes, D. Dominance of ST131, B2, blaCTX-M-15, and papA-papC-kpsMII-uitA among ESBL Escherichia coli isolated from bloodstream infections in Quito, Ecuador: A 10-year surveillance study (2009–2019). J. Appl. Microbiol. 2023, 134, lxad269. [Google Scholar] [CrossRef] [PubMed]
- Elankumaran, P.; Cummins, M.L.; Browning, G.F.; Marenda, M.S.; Reid, C.J.; Djordjevic, S.P. Genomic and Temporal Trends in Canine ExPEC Reflect Those of Human ExPEC. Microbiol. Spectr. 2022, 10, e0129122. [Google Scholar] [CrossRef]
- Valat, C.; Drapeau, A.; Beurlet, S.; Bachy, V.; Boulouis, H.-J.; Pin, R.; Cazeau, G.; Madec, J.-Y.; Haenni, M. Pathogenic Escherichia coli in Dogs Reveals the Predominance of ST372 and the Human-Associated ST73 Extra-Intestinal Lineages. Front. Microbiol. 2020, 11, 580. [Google Scholar] [CrossRef] [PubMed]
- Turchetto, S.; Ustulin, M.; Citterio, C.V.; Zanardello, C.; Vascellari, M.; Vio, D.; Conedera, G.; Milone, N.M.F.; Cocchi, M. Phenotypic features and phylogenetic background of extraintestinal hemolytic Escherichia coli responsible of mortality in puppies. Vet. Microbiol. 2015, 179, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Handt, L.K.; Stoffregen, D.A.; Prescott, J.S.; Pouch, W.J.; Ngai, D.T.; Anderson, C.A.; Gatto, N.T.; DebRoy, C.; Fairbrother, J.M.; Motzel, S.L.; et al. Clinical and microbiologic characterization of hemorrhagic pneumonia due to extraintestinal pathogenic Escherichia coli in four young dogs. Comp. Med. 2003, 53, 663–670. [Google Scholar]
- Caruso, G.; Giammanco, A.; Cardamone, C.; Oliveri, G.; Mascarella, C.; Capra, G.; Fasciana, T. Extra-Intestinal Fluoroquinolone-Resistant Escherichia coli Strains Isolated from Meat. Biomed. Res. Int. 2018, 2018, 8714975. [Google Scholar] [CrossRef] [PubMed]
- Davidova-Gerzova, L.; Lausova, J.; Sukkar, I.; Nesporova, K.; Nechutna, L.; Vlkova, K.; Chudejova, K.; Krutova, M.; Palkovicova, J.; Kaspar, J.; et al. Hospital and community wastewater as a source of multidrug-resistant ESBL-producing Escherichia coli. Front. Cell Infect. Microbiol. 2023, 13, 1184081. [Google Scholar] [CrossRef]
- Meijs, A.P.; Gijsbers, E.F.; Hengeveld, P.D.; Dierikx, C.M.; de Greeff, S.C.; van Duijkeren, E. ESBL/pAmpC-producing Escherichia coli and Klebsiella pneumoniae carriage among veterinary healthcare workers in The Netherlands. Antimicrob. Resist. Infect. Control. 2021, 10, 147. [Google Scholar] [CrossRef]
- Newell, D.G.; La Ragione, R.M. Enterohaemorrhagic and other Shiga toxin-producing Escherichia coli (STEC): Where are we now regarding diagnostics and control strategies? Transbound. Emerg Dis. 2018, 65 (Suppl. S1), 49–71. [Google Scholar] [CrossRef]
- Lappin, M.; Blondeau, J.; Boothe, D.; Breitschwerdt, E.; Guardabassi, L.; Lloyd, D.; Papich, M.; Rankin, S.; Sykes, J.; Turnidge, J.; et al. Antimicrobial use Guidelines for Treatment of Respiratory Tract Disease in Dogs and Cats: Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases. J. Vet. Intern. Med. 2017, 31, 279–294. [Google Scholar] [CrossRef]
- Bostek, C.C. Oxygen toxicity: An introduction. Aana J. 1989, 57, 231–237. [Google Scholar] [PubMed]
- Tannenbaum, J.; Bennett, B.T. Russell and Burch’s 3Rs then and now: The need for clarity in definition and purpose. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 120–132. [Google Scholar] [PubMed]
| Antimicrobial Categories | Antimicrobial Agent | Disk Content/μg | Standards/mm | ||
|---|---|---|---|---|---|
| S | I | R | |||
| Cephalosporins | Ceftazidime | 30 | ≥21 | 18–20 | ≤17 |
| Cefazolin | 30 | ≥23 | 20–22 | ≤19 | |
| Cephalexin | 30 | ≥21 | 18–20 | ≤17 | |
| Cefpodoxime | 10 | ≥21 | 18–20 | ≤17 | |
| Ceftiofur | 30 | ≥21 | 18–20 | ≤17 | |
| Cephalothin | 30 | ≥18 | 15–17 | ≤14 | |
| Tetracyclines | Tetracycline | 30 | ≥15 | 12–14 | ≤11 |
| Doxycycline | 30 | ≥16 | 13–15 | ≤12 | |
| Aminoglycosides | Gentamicin | 10 | ≥16 | 13–15 | ≤12 |
| Amikacin | 30 | ≥20 | 17–19 | ≤16 | |
| Neomycin | 30 | ≥17 | 13–16 | ≤12 | |
| Quinolones | Levofloxacin | 5 | ≥21 | 17–20 | ≤16 |
| Pradofloxacin | 5 | ≥24 | 20–23 | ≤19 | |
| Marbofloxacin | 5 | ≥20 | 15–19 | ≤14 | |
| Enrofloxacin | 10 | ≥19 | 15–18 | ≤14 | |
| Penicillins | Ampicillin | 10 | ≥17 | - | - |
| β-lactam combination agents | Amoxicillin–clavulanate | 20 | ≥18 | – | – |
| Piperacillin–tazobactam | 100/10 | ≥25 | 21–24 | ≤20 | |
| Carbapenems | Imipenem | 10 | ≥23 | 20–22 | ≤19 |
| Chloramphenicols | Chloramphenicol | 30 | ≥21 | 18–20 | ≤17 |
| Sulfonamides | Cotrimoxazole | 23.75/1.25 | ≥16 | 10–15 | ≤9 |
| Sample | Raw Reads | Clean Reads | Average Read Length (bp) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|
| 1 | 10,005,434 | 10,000,031 | 149.10 | 94.69 | 50.76 |
| 2 | 10,133,302 | 10,127,526 | 149.48 | 95.60 | 50.73 |
| 3 | 12,429,312 | 12,421,978 | 149.24 | 96.14 | 50.58 |
| 4 | 7,883,010 | 7,878,437 | 149.79 | 93.85 | 50.43 |
| Antimicrobial Categories | Antimicrobial Name | Antibiotic Susceptibility Testing | Sensitivity | ||
|---|---|---|---|---|---|
| R | I | S | |||
| Cephalosporins | Ceftazidime | 3 | 1 | 0 | 0% |
| Cefazolin | 4 | 0 | 0 | 0% | |
| Cephalexin | 4 | 0 | 0 | 0% | |
| Cefpodoxime | 4 | 0 | 0 | 0% | |
| Ceftiofur | 4 | 0 | 0 | 0% | |
| Cephalothin | 4 | 0 | 0 | 0% | |
| Tetracyclines | Tetracycline | 4 | 0 | 0 | 0% |
| Doxycycline | 1 | 3 | 0 | 0% | |
| Aminoglycosides | Gentamicin | 1 | 0 | 3 | 75% |
| Amikacin | 0 | 0 | 4 | 100% | |
| Neomycin | 0 | 0 | 4 | 100% | |
| Quinolones | Levofloxacin | 1 | 0 | 3 | 75% |
| Pradofloxacin | 0 | 0 | 4 | 100% | |
| Marbofloxacin | 0 | 0 | 4 | 100% | |
| Enrofloxacin | 0 | 0 | 0 | 100% | |
| Penicillins | Ampicillin | 4 | 0 | 0 | 0% |
| β-lactamase inhibitors | Amoxicillin-clavulanate | 0 | 0 | 4 | 100% |
| Piperacillin-tazobactam | 0 | 0 | 4 | 100% | |
| Carbapenems | Imipenem | 0 | 0 | 4 | 100% |
| Chloramphenicol | Chloramphenicol | 0 | 0 | 4 | 100% |
| Sulfonamides | Cotrimoxazole | 0 | 0 | 4 | 100% |
| Gene Name | Gene Type | Mechanism |
|---|---|---|
| TEM-206 | TEM beta-lactamase | antibiotic inactivation |
| CTX-M-55 | CTX-M beta-lactamase | |
| mecA | methicillin-resistant PBP2 | antibiotic target replacement |
| mecC | ||
| tet(X4) | tetracycline inactivation enzyme | antibiotic inactivation |
| tetB(P) | tetracycline-resistant ribosomal protection protein | antibiotic target protection |
| tet(Q) | ||
| tetB(60) | ATP-binding cassette (ABC) antibiotic efflux pump | antibiotic efflux |
| tetA(46) | ||
| tetB(46) | ||
| tetA(60) | ||
| tetB(46) | ||
| tet(30) | major facilitator superfamily (MFS) antibiotic efflux pump | |
| tet(C) | ||
| tetA(58) | ||
| cmlA9 | ||
| cmlA6 | ||
| AAC(3)-IId | AAC(3) | antibiotic inactivation |
| dfrA21 | trimethoprim-resistant dihydrofolate reductase dfr | antibiotic target replacement |
| sul2 | sulfonamide resistant sul | |
| sul4 |
| Gene Name | Gene Type | Function |
|---|---|---|
| CNF-I | Cytotoxic necrotizing factor 1 | Induces the formation of actin stress fibers and membrane ruffling, necrosis |
| fim2 | Serotype 2 fimbrial subunit precursor | Fimbriae may mediate the binding of Bordetella to respiratory epithelium via the major fimbrial subunits and to monocytes via FimD |
| fimA | Type 1 fimbrial protein, A chain precursor, fimbrial protein | Makes an important contribution to the colonization of the bladder |
| fimB | Type 1 fimbriae regulatory protein fimB | |
| fimD | Outer membrane usher protein fimD precursor | |
| fimE | Type 1 fimbriae regulatory protein fimE | |
| fimF | FimF protein precursor | |
| fimG | FimG protein precursor | |
| fimH | FimH protein precursor | |
| fimI | Fimbrin-like protein fimI precursor | |
| fimC | Chaperone protein fimC (precursor) | Adherence; invasion |
| fyuA | Pesticin/yersiniabactin receptor | FyuA/Psn-Irp system uses yersiniabactin, a siderophore that can remove iron from a number of mammalian proteins due to its extremely high affinity for ferric iron |
| irp1 | Yersiniabactin biosynthetic protein Irp1 | |
| Irp2 | Yersiniabactin biosynthetic protein Irp2 | |
| iroN | Salmochelin receptor IroN | Catecholate siderophore receptor, mediates utilization of the siderophore enterobactin |
| ompA | Outer membrane protein P5 (ompA), human factor H binding protein | Interacts with CEACAM1, a member of the carcinoembryonic antigen (CEA) family of cell adhesion molecules, a glycoprotein expressed by respiratory epithelial cell |
| papA | P pilus major subunit PapA | Pap pili expression, which is mannose-resistant, is tightly regulated by environmental and nutritional factors and a methylation-dependent phase variation mechanism. The pap operon is a key example of pilus assembly, featuring conserved elements like PapD, an Ig-like domain chaperone essential for transporting pilus subunits from the cytoplasm to the outer membrane. PapD–subunit complexes are directed to the PapC usher in the outer membrane, forming a pore for pilus translocation. The main subunit, PapA, assembles into a 6.8 nm helical rod anchored by PapH. The pilus rod’s distal end has a 2 nm linear tip fibrillum made of PapE connected to the PapA rod by PapK. PapG attaches to the PapE tip fibrillum via the adapter protein PapF. |
| papB | Regulatory protein PapB | |
| papC | Usher protein PapC | |
| papE | P pilus minor subunit PapE | |
| papF | P pilus minor subunit PapF | |
| papG | P pilus tip adhesin PapG | |
| papH | P pilus termination subunit PapH | |
| papI | Regulatory protein PapI | |
| papJ | P pilus assembly protein PapJ | |
| papK | P pilus minor subunit PapK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, J.; Long, H.; Zhao, Z.; Rao, G.; Ou, Z.; Li, J.; Zhou, Z.; Hu, M.; Ni, Q. Characterization of Extraintestinal Pathogenic Escherichia coli Strains Causing Canine Pneumonia in China: Antibiotic Resistance, Virulence Genes, and Sequence Typing. Vet. Sci. 2024, 11, 491. https://doi.org/10.3390/vetsci11100491
Lai J, Long H, Zhao Z, Rao G, Ou Z, Li J, Zhou Z, Hu M, Ni Q. Characterization of Extraintestinal Pathogenic Escherichia coli Strains Causing Canine Pneumonia in China: Antibiotic Resistance, Virulence Genes, and Sequence Typing. Veterinary Sciences. 2024; 11(10):491. https://doi.org/10.3390/vetsci11100491
Chicago/Turabian StyleLai, Jianyi, Haibin Long, Zhihong Zhao, Gan Rao, Zhaojia Ou, Jiajie Li, Zhidong Zhou, Minhua Hu, and Qingchun Ni. 2024. "Characterization of Extraintestinal Pathogenic Escherichia coli Strains Causing Canine Pneumonia in China: Antibiotic Resistance, Virulence Genes, and Sequence Typing" Veterinary Sciences 11, no. 10: 491. https://doi.org/10.3390/vetsci11100491
APA StyleLai, J., Long, H., Zhao, Z., Rao, G., Ou, Z., Li, J., Zhou, Z., Hu, M., & Ni, Q. (2024). Characterization of Extraintestinal Pathogenic Escherichia coli Strains Causing Canine Pneumonia in China: Antibiotic Resistance, Virulence Genes, and Sequence Typing. Veterinary Sciences, 11(10), 491. https://doi.org/10.3390/vetsci11100491