
Study of Dietary Emodin on Immune Defense in Megalobrama amblycephala against Aeromonas hydrophila

 ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Fish and Breeding Management
2.3. Challenge Test
2.3.1. Preparation of Fungal Broth
2.3.2. Experiment Design
2.4. Sample Collection
2.5. Colony Enumeration
2.6. Immunological Assays
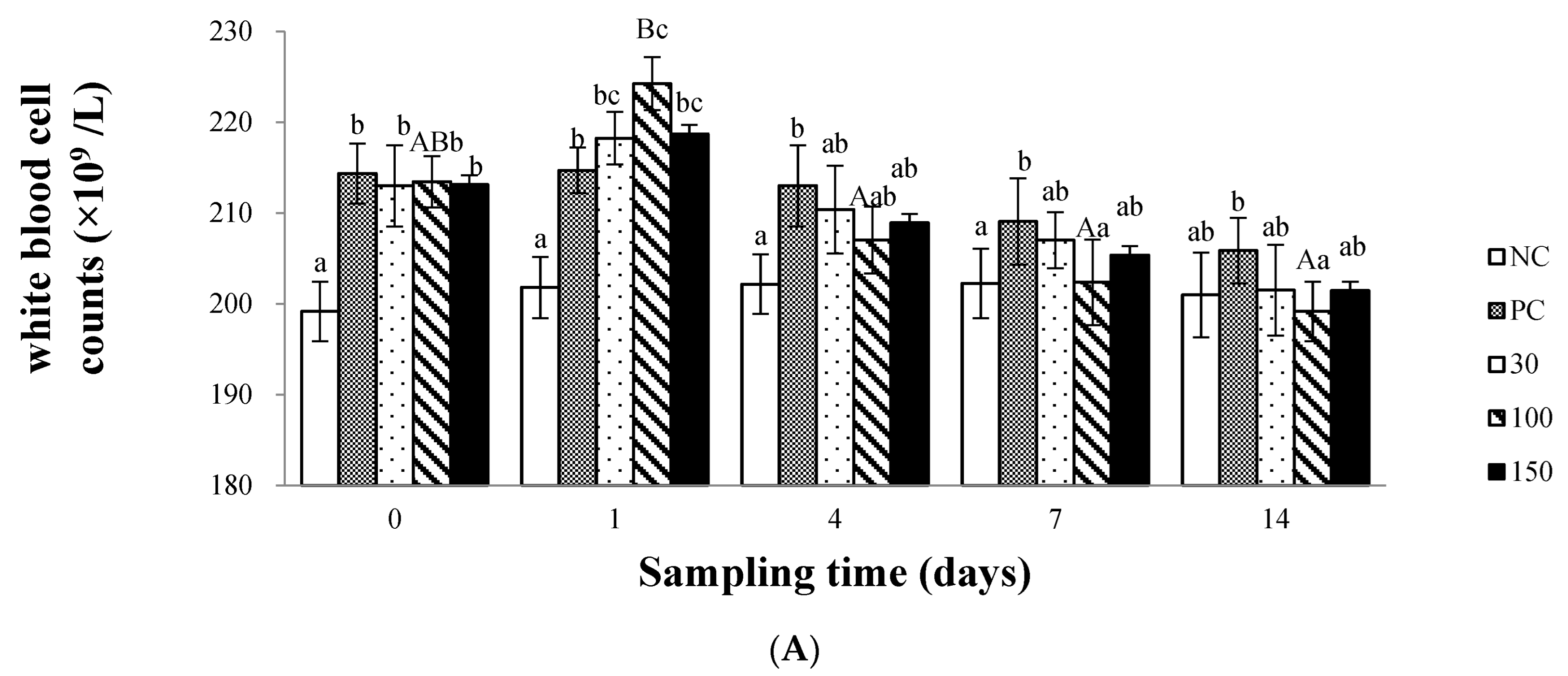
2.6.1. White Blood Cell Count (WBC)
2.6.2. Respiratory Burst Activity
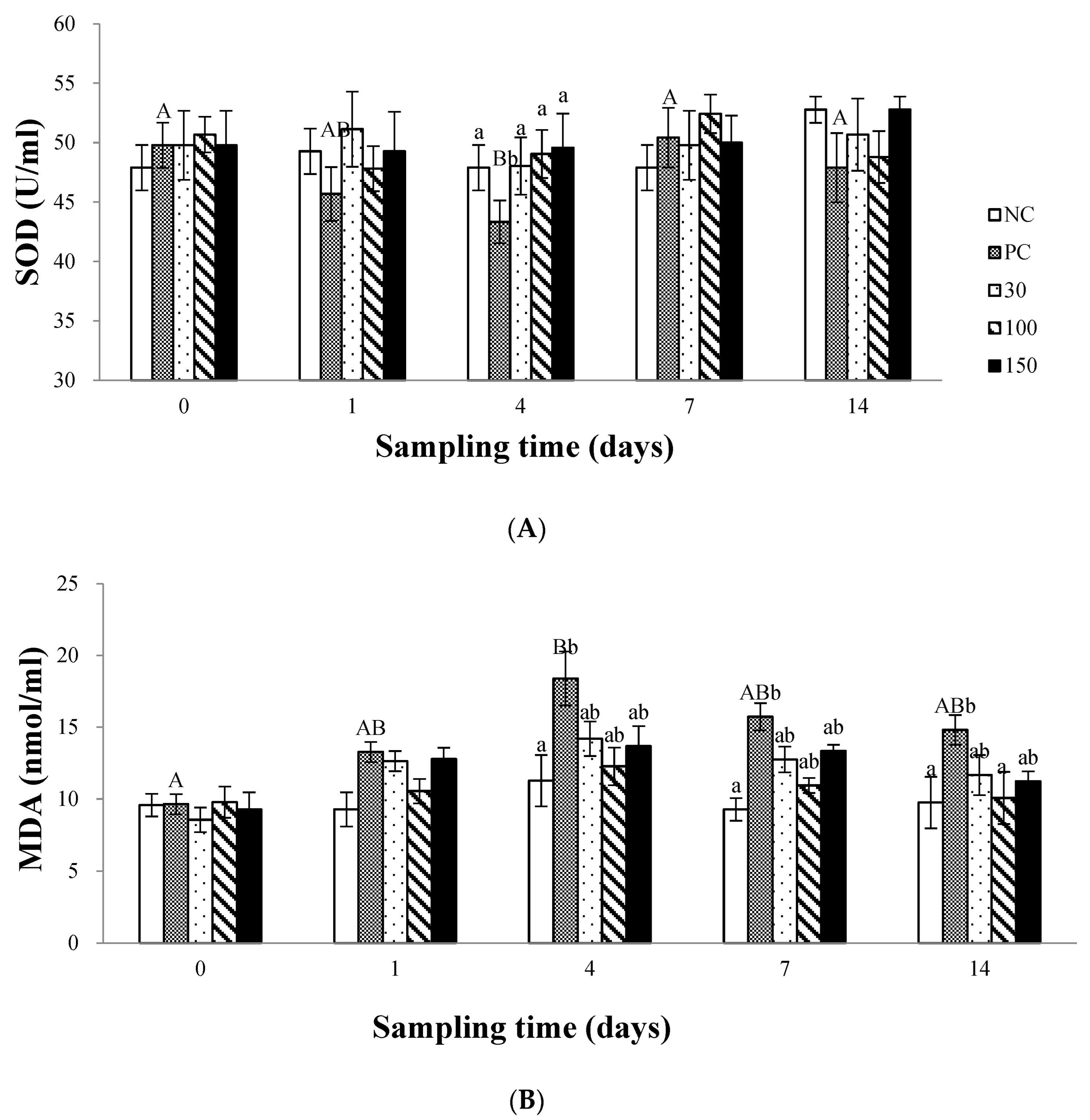
2.6.3. Plasma Superoxide Dismutase (SOD) and Malondialdehyde (MDA) Assay
2.6.4. Plasma Tumor Necrosis Factor-α (TNF-α) and Interleukin-1 (IL-1) Assay
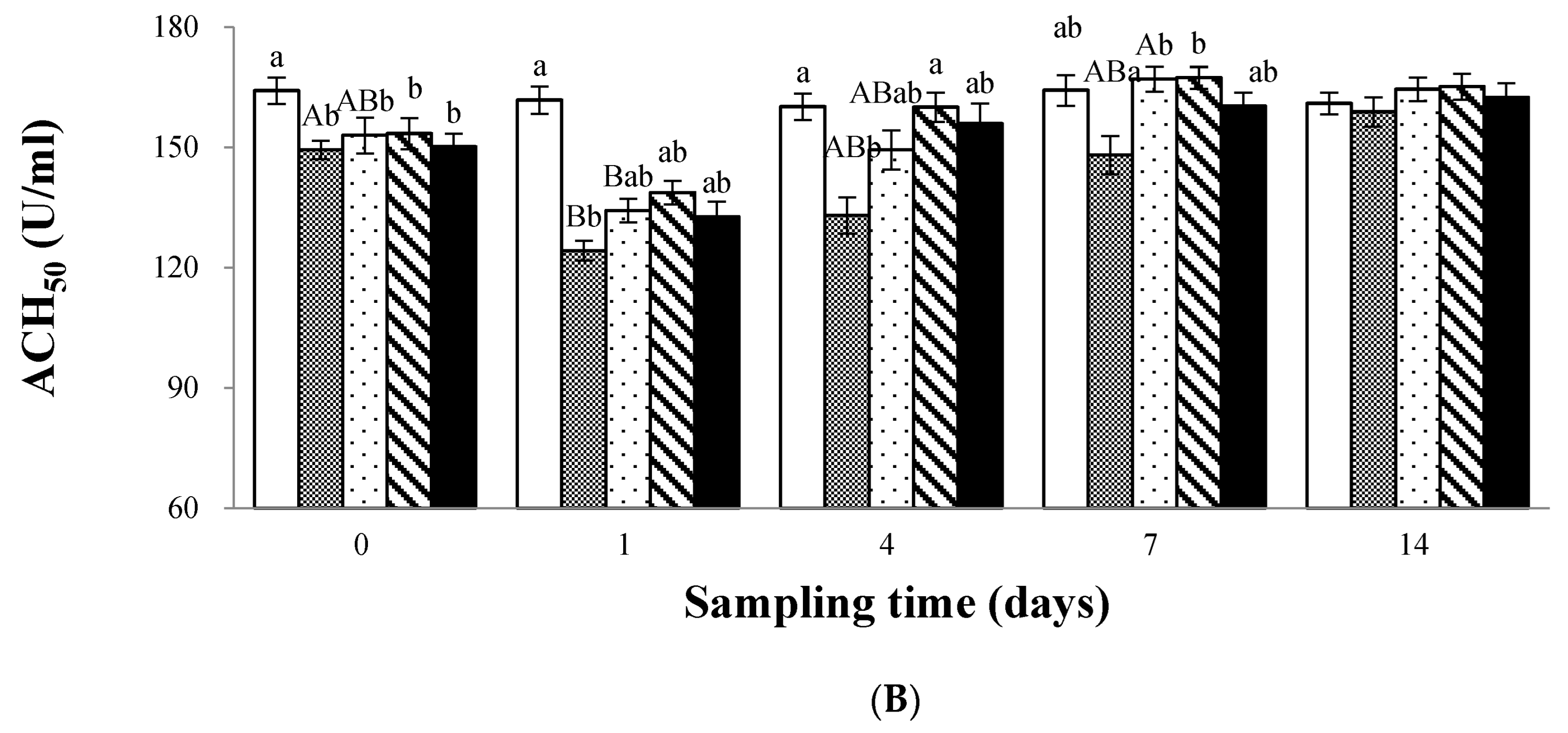
2.6.5. Complement Activity Assay
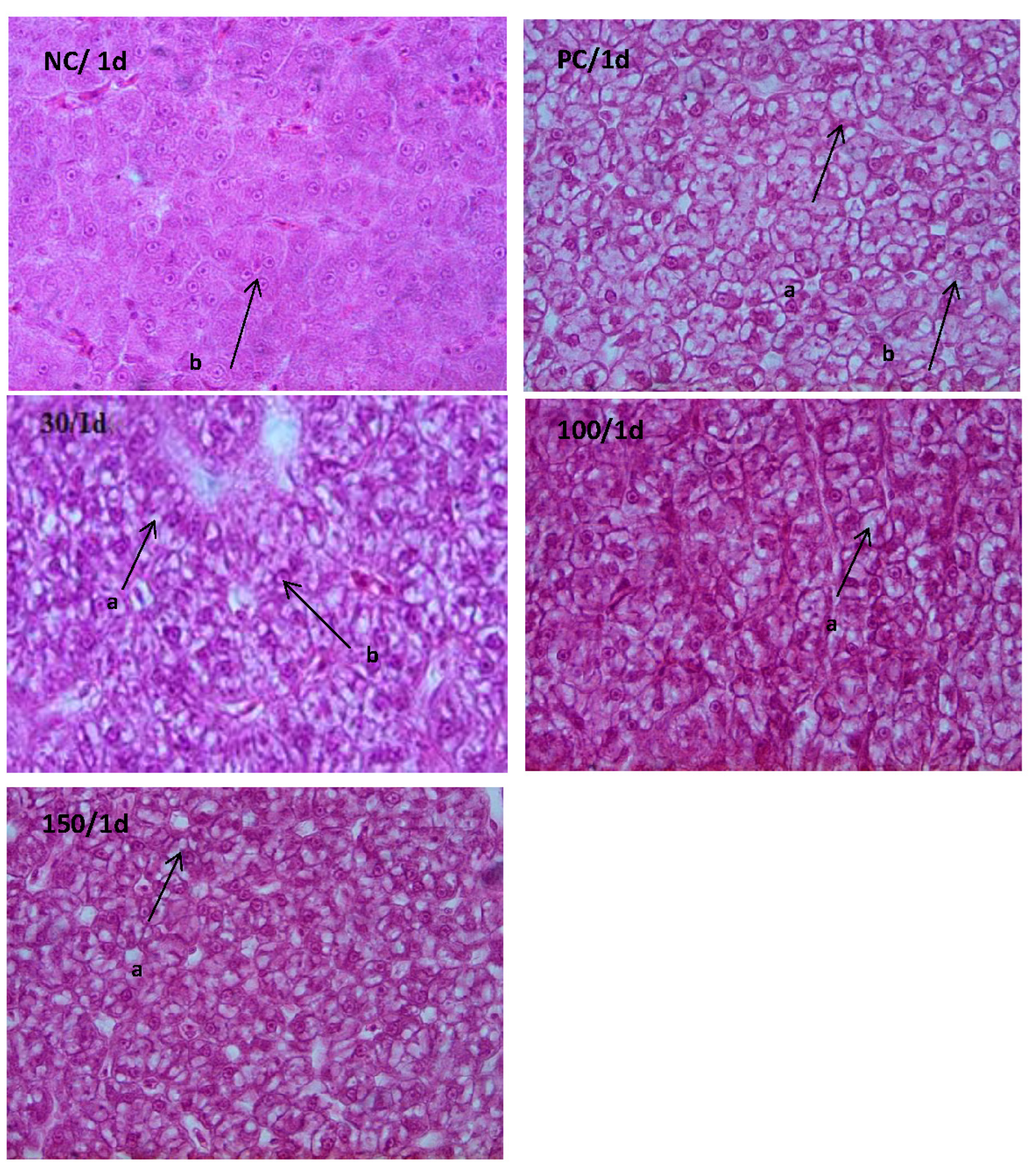
2.7. Histological Examination
2.8. Statistical Analysis
3. Results
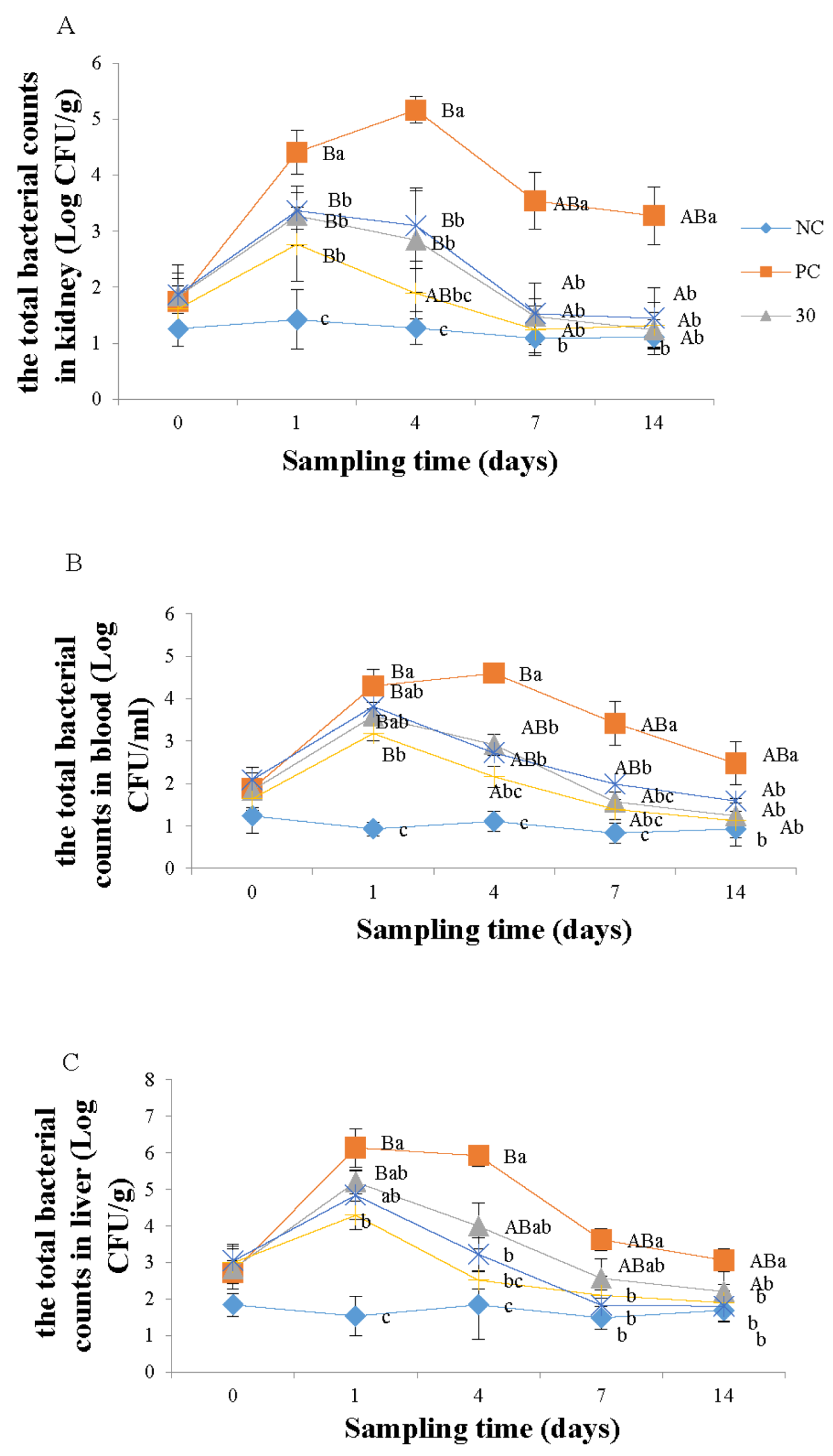
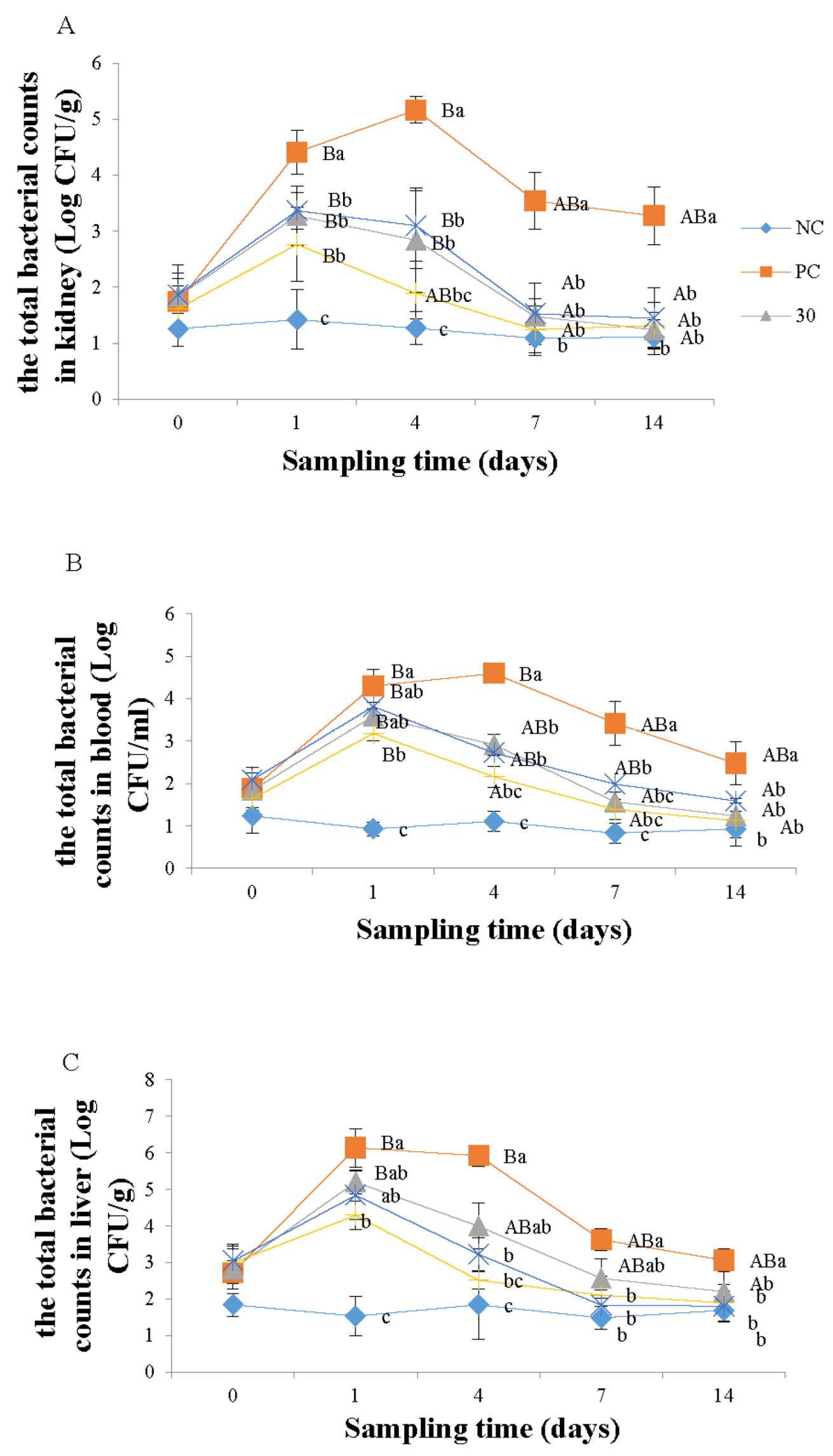
3.1. Enumeration of Monobacterial Colonies of the Genus Aeromonas spp.
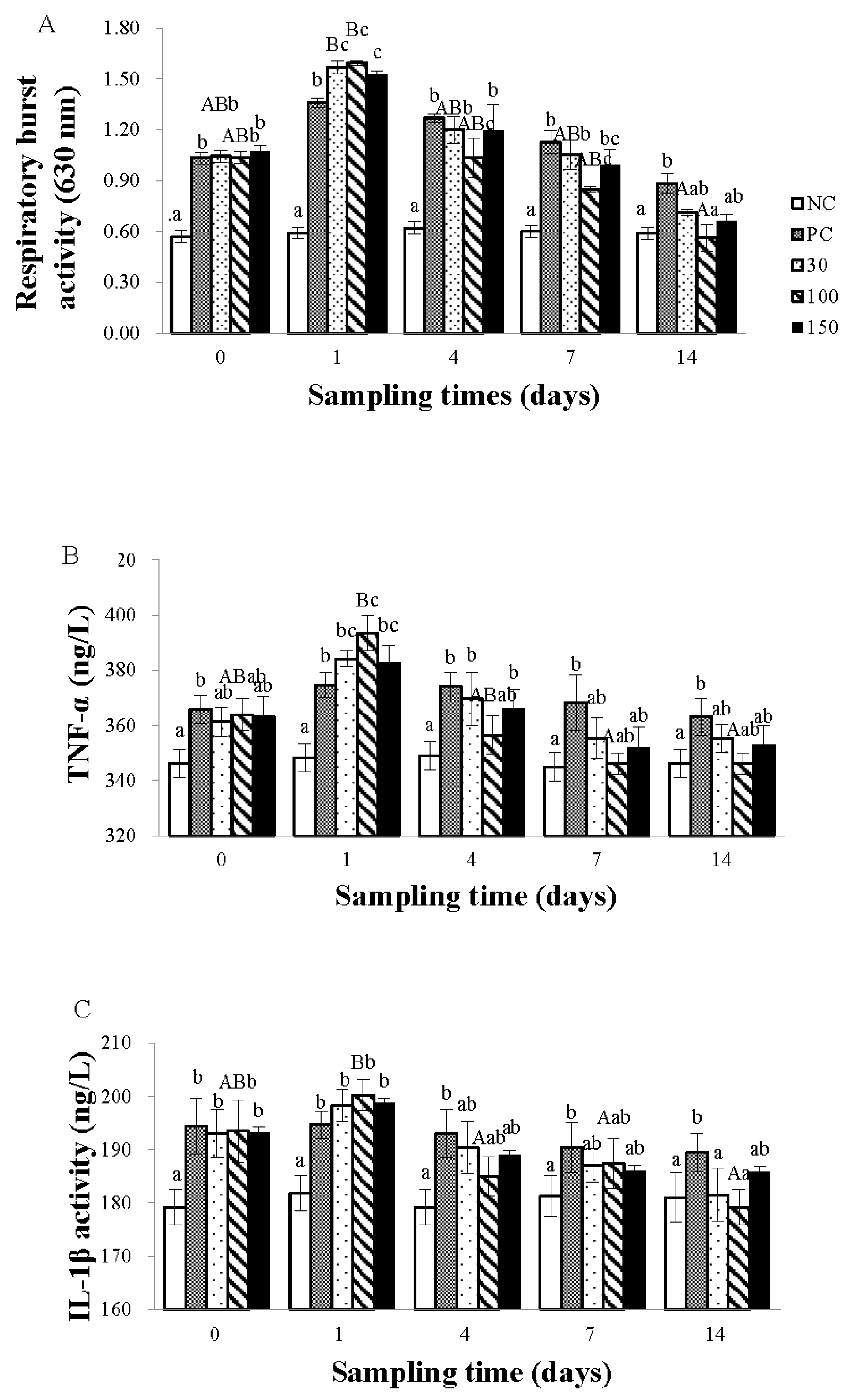
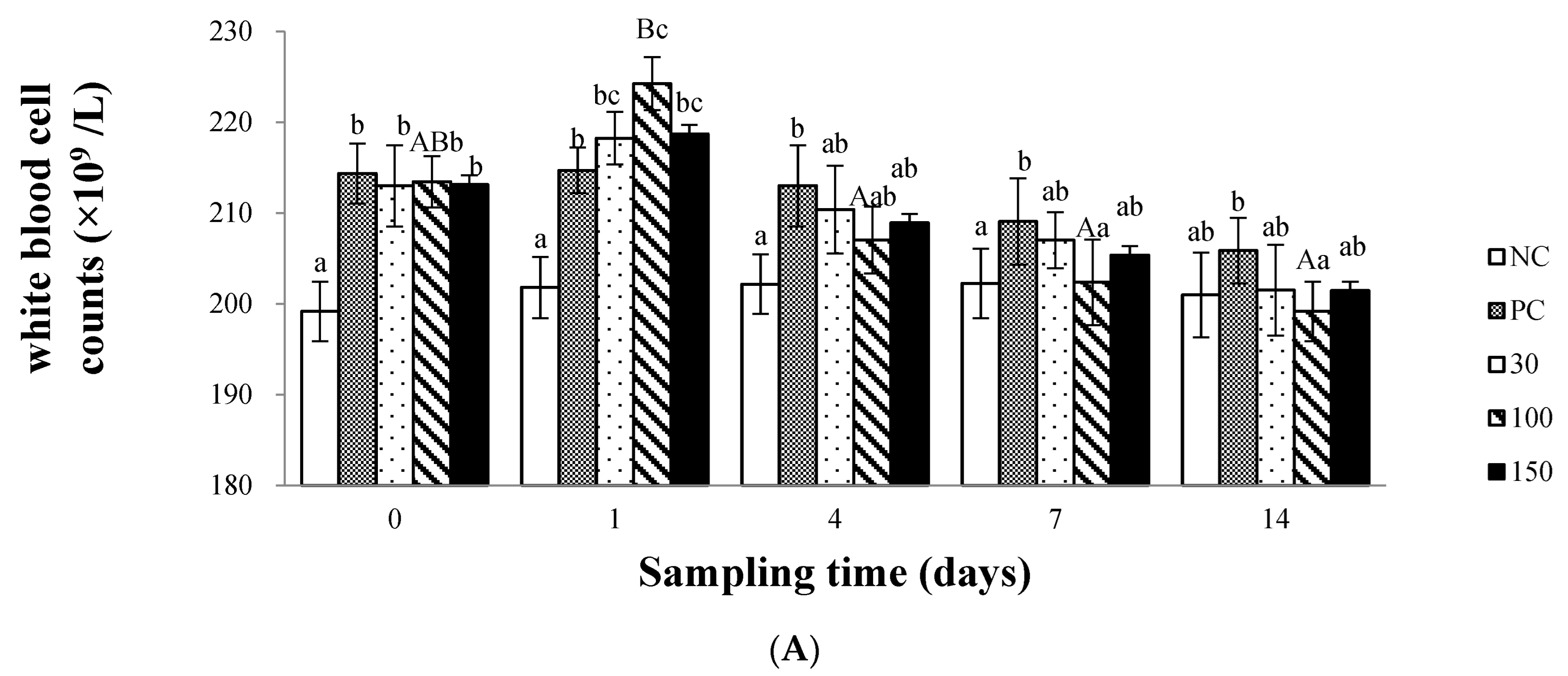
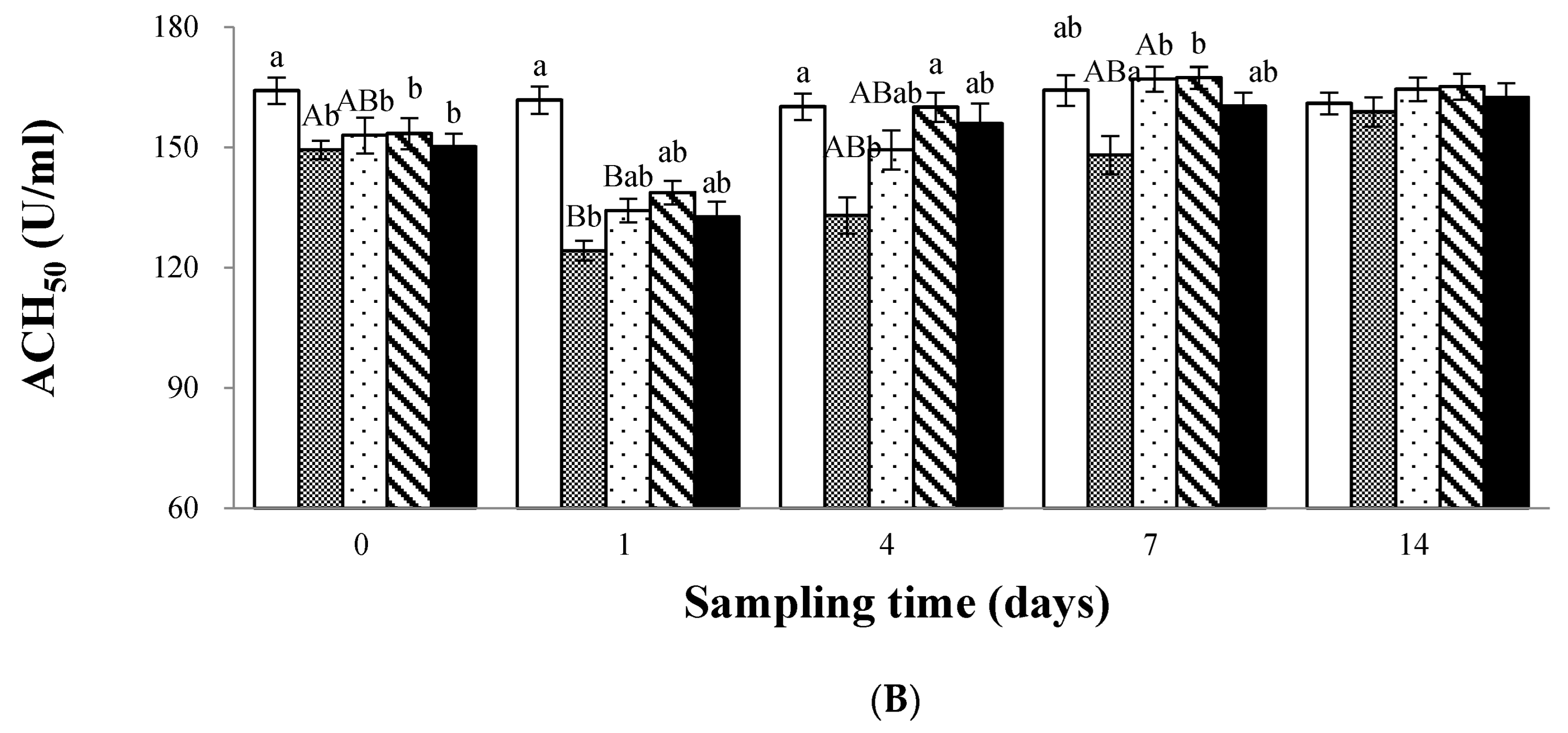
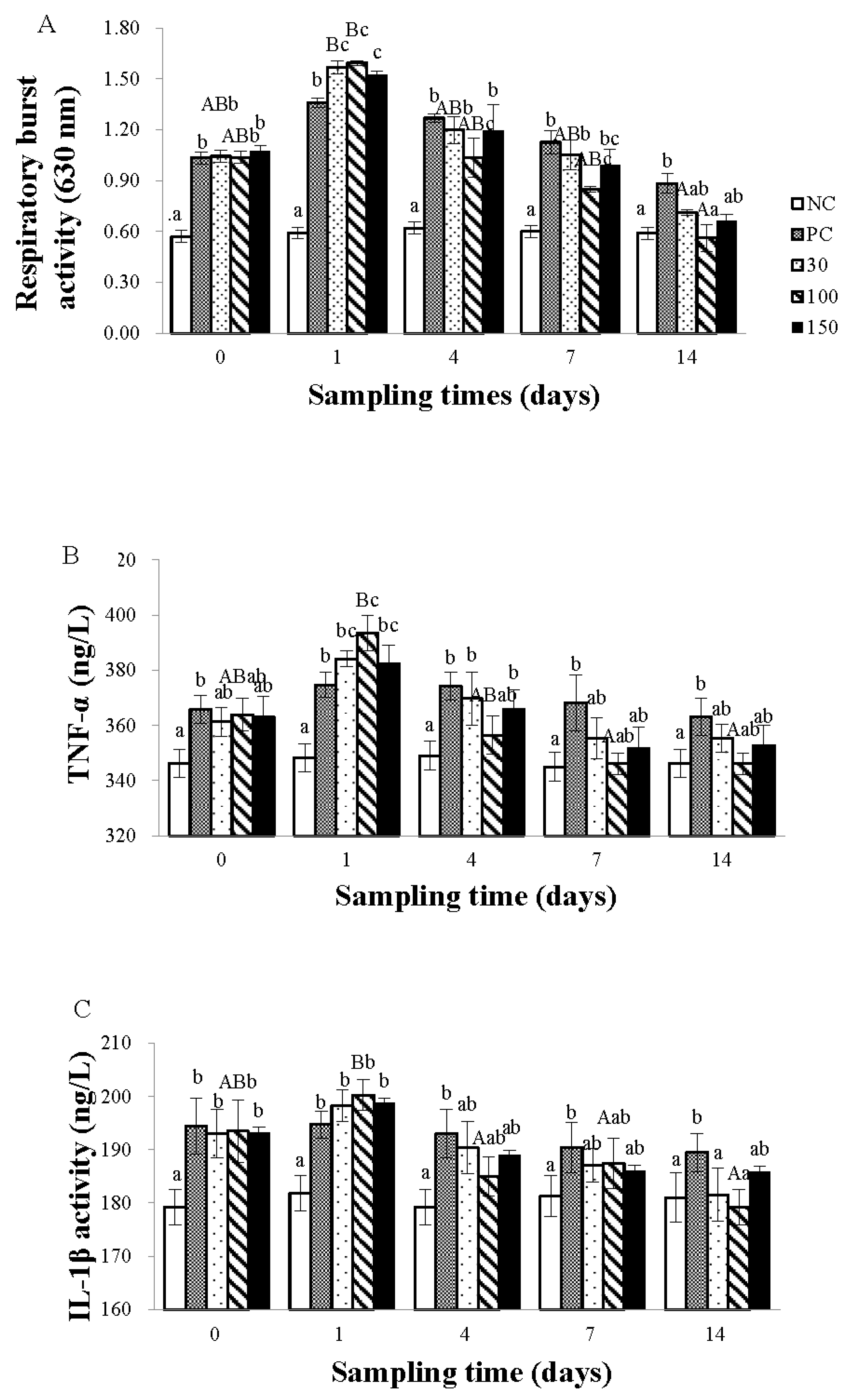
3.2. Immune Response
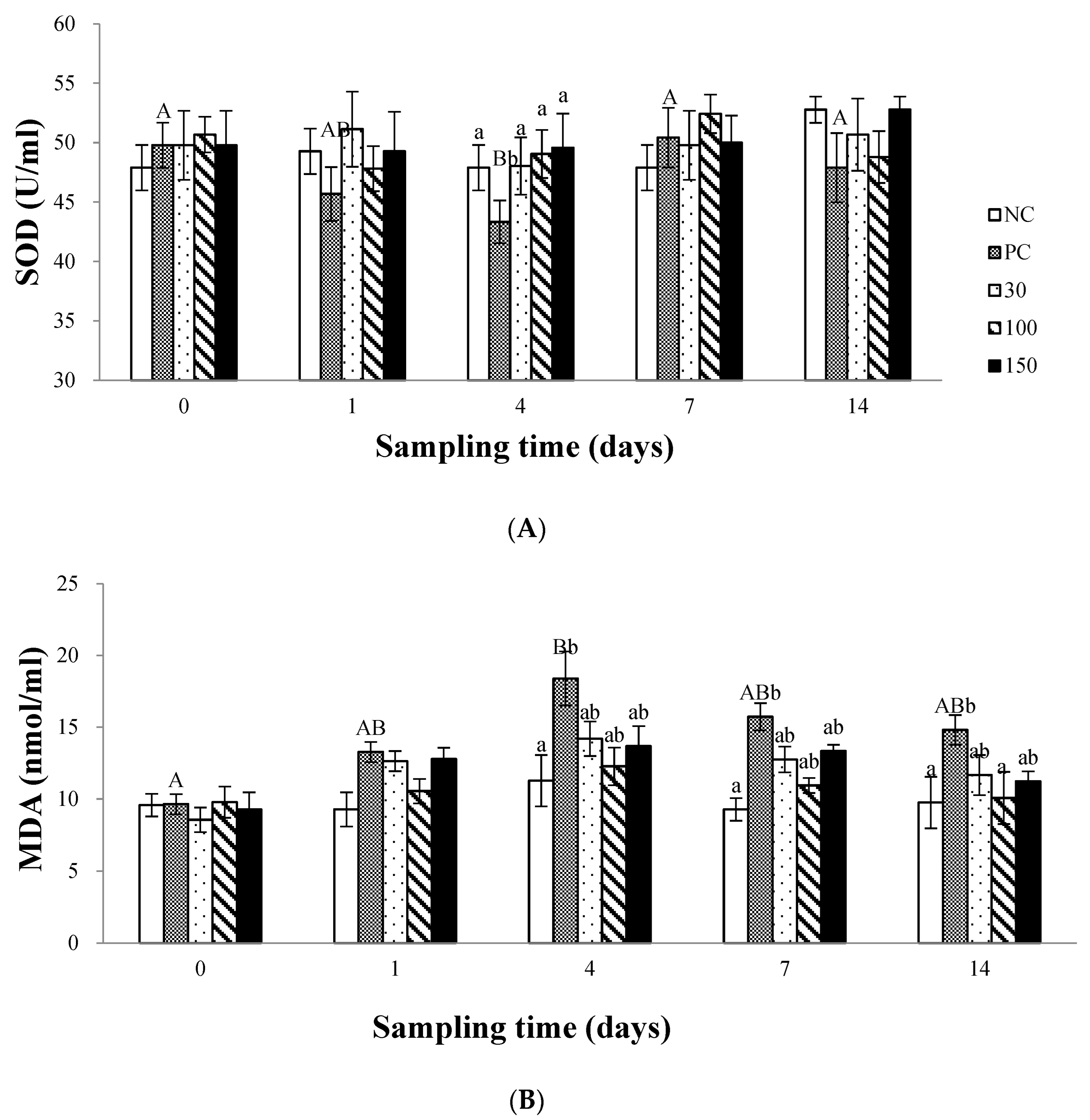
3.3. The Antioxidant Status in Plasma
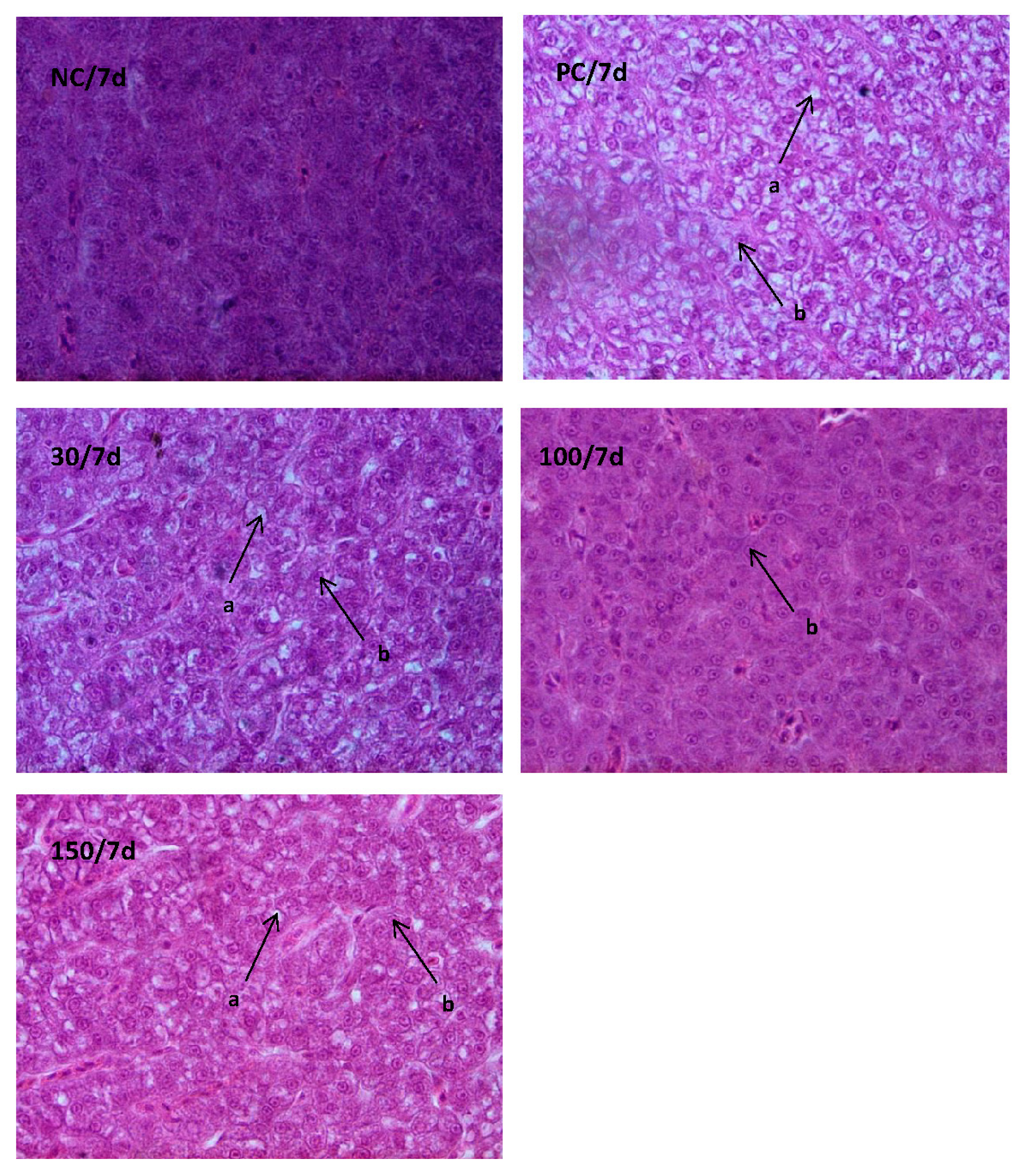
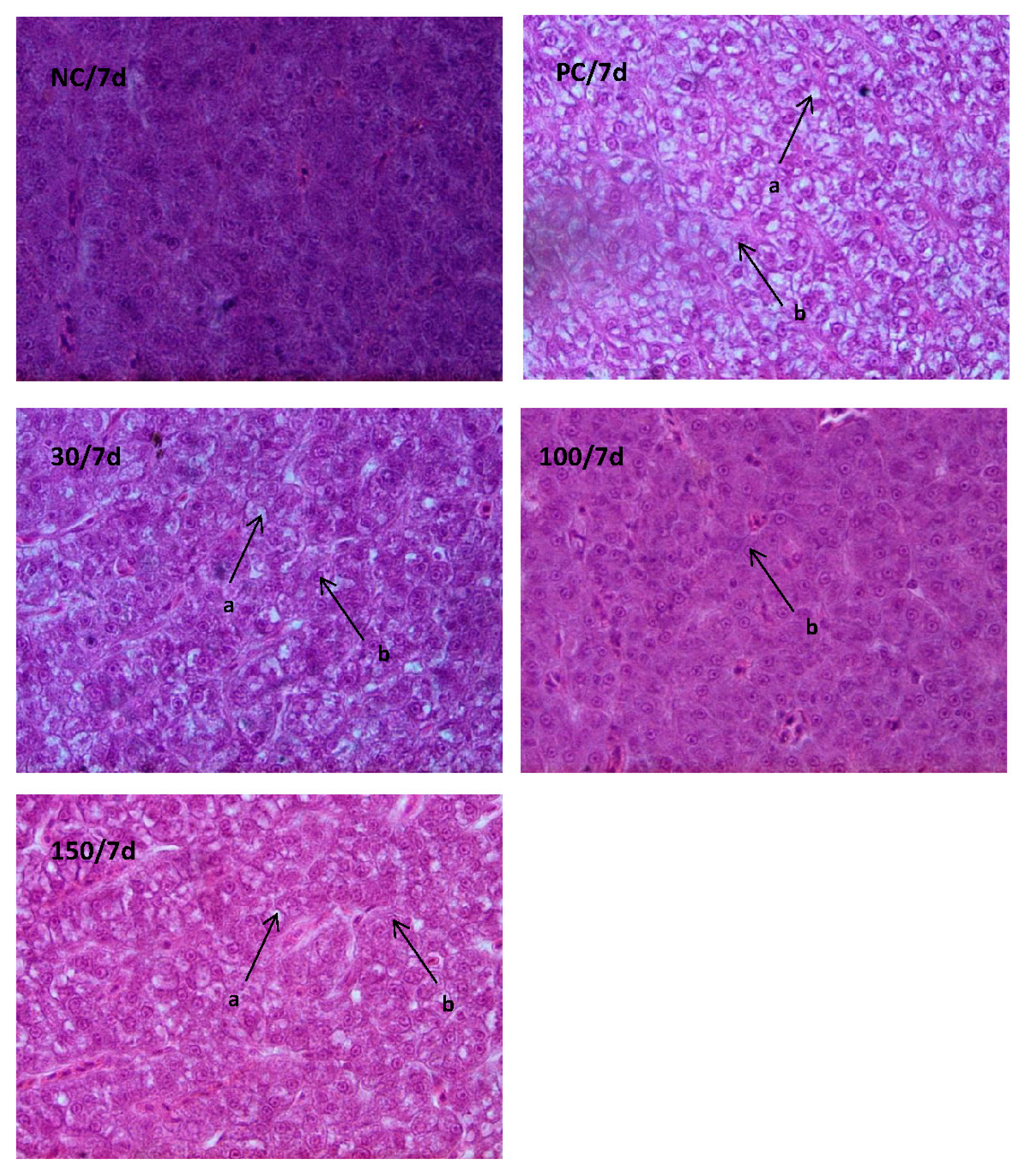
3.4. Histological Examination
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhai, G.C. The Biological Characteristics and Hazards of Aeromonas hydrophila. Inland Fish. 1997, 22, 21. (In Chinese) [Google Scholar]
- Larsen, J.L.; Jensen, N.J. An Aeromonas species implicated in ulcer-disease of the cod (Gadus morhua). Nord. Veterinaermedicin 1976, 29, 199–211. [Google Scholar]
- Feng, D.Y. Survey and evaluation of diseases found in freshwater fishes in 2009. Chin. J. Fish. 2010, 4, 60–65. [Google Scholar]
- Harikrishnan, R.; Balasundaram, C. In vitro and in vivo studies of the use of some medicinal herbals against the pathogen Aeromonas hydrophila in goldfish. J. Aquat. Anim. Health 2008, 20, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Liu, B.; Zhou, Q.L.; Su, Y.; He, Y.; Pan, L.; Xu, P. Effects of anthraquinones extract from rhubarb R. officinale Bail on the crowding stress response and growth of common carp (Cyprinus carpio var. Jian). Aquaculture 2008, 281, 5–11. [Google Scholar] [CrossRef]
- Chang, C.H.; Lin, C.C.; Yang, J.J.; Namba, T.; Hattori, M. Anti-inflammatory effects of emodin from ventilago leiocarpa. Am. J. Chin. Med. 1996, 24, 139–142. [Google Scholar] [CrossRef]
- Huang, S.S.; Yeh, S.F.; Hong, C.Y. Effect of anthraquinone derivatives on lipid peroxidation in rat heart mitochondria: Structure activity relationship. J. Nat. Prod. 1995, 58, 1365–1371. [Google Scholar] [CrossRef]
- Zhou, L.L.; Shu, X.C.; Wu, H.P.; Li, S.P.; Han, W.; Li, H. Curative effects of emodin on fatty livers in quails. Chin. J. Clin. Rehabil. 2006, 10, 57–59. (In Chinese) [Google Scholar]
- Lin, C.C.; Chang, C.H.; Yang, J.J.; Namba, T.; Hattori, M. Hepatoprotective effects of emodin from Ventilago Leiocarpa. J. Ethnopharmacol. 1996, 52, 107–111. [Google Scholar] [CrossRef]
- Wang, W.J.; Wu, X.Z.; Yao, Z.; Li, H.Q. The influence of emodin and danshensu on monocyte’s secretion of inflammatory cytokines. Chin. J. Immunol. 1995, 1, 370–382. (In Chinese) [Google Scholar]
- Liu, B.; Ge, X.; Xie, J.; Xu, P.; He, Y.; Cui, Y.; Pan, L. Effects of anthraquinone extract from Rheum officinale Bail on the physiological responses and HSP70 gene expression of Megalobrama amblycephala under Aeromonas hydrophila infection. Fish Shellfish. Immunol. 2012, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.H.; Xie, J.; Xu, P.; Ge, X.; Liu, W.; Ye, J. Effects of emodin and vitamin C on growth performance, biochemical parameters and two HSP70s mRNA expression of Wuchang bream (Megalobrama amblycephala Yih) under high temperature stress. Fish Shellfish. Immunol. 2012, 32, 651–661. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Liu, B.; Ge, X.P.; Pan, L. Comparative study of antibacterial properties of emodin and enrofloxacin against Aeromonas hydrophila. Isr. J. Aquac. Bamidgeh 2014, 66. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Liu, B.; Xie, J.; Liu, W.B.; Xie, J.; Ren, M.C.; Pan, L.K. Effects of dietary emodin supplementation on growth performance, non-specific immune responses, and disease resistance to Aeromonas hydrophila in juvenile Wuchang bream (Megalobrama amblycephala). Isr. J. Aquac. Bamidgeh 2014, 66. [Google Scholar] [CrossRef]
- Secombes, C.J.; Stolen, J.S..; Fletcher, T.C.; Anderson, D.P.; Robertsen, B.S.; Van Muiswinkel, W.B. Isolation of salmonid macrophages and analysis of their killing activity. In Techniques in Fish Immunol.; SOS Publications: Fair Haven, NJ, USA, 1990; pp. 137–154. [Google Scholar]
- Ai, Q.H.; Mai, K.S.; Zhang, L.; Tan, B.; Zhang, W.; Xu, W.; Li, H. Effects of dietary b-1,3 glucan on innate immune response of large yellow croaker, Pseudosciaena crocea. Fish Shellfish. Immunol. 2007, 22, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Granelli, K.; Björck, L.; Appelqvist, L.A. The variation of superoxide dismutase (SOD) and xanthine oxidase (XO) activities in milk using an improved method to quantitate SOD activity. J. Sci. Food Agric. 1995, 67, 85–91. [Google Scholar] [CrossRef]
- Drape, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free. Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- Zhao, Z.; Xie, J.; Liu, B.; Ge, X.; Song, C.; Ren, M.; Zhou, Q.; Miao, L.; Zhang, H.; Shan, F.; et al. The effects of emodin on cell viability, respiratory burst and gene expression of Nrf2-Keap1 signaling molecules in the peripheral blood leukocytes of blunt snout bream (Megalobrama amblycephala). Fish Shellfish. Immunol. 2017, 62, 75–85. [Google Scholar] [CrossRef]
- Li, X.W.; Jia, Z.H.; Wang, W.L.; Wang, L.L.; Liu, Z.Q.; Yang, B.; Jia, Y.K.; Song, X.R.; Yi, Q.L.; Qiu, L.M.; et al. Glycogen synthase kinase (gsk3) regulates TNF production and haemocyte phagocytosis in the immune response of Chinese mitten crab eriocheir sinensis. Dev. Comp. Immunol. 2017, 73, 144–155. [Google Scholar] [CrossRef]
- Ortuño, J.; Esteban, M.A.; Mulero, V.; Meseguer, J. Methods for studying the hae-molytic, chemoattractant and opsonic activities of seabream (Sparus aurata L.). In Methodology in Fish Diseases; Research, A.C., Barnes, G.A., Davidson, M., Hiney, D.M., Eds.; Albion Press: Aberdeen, UK, 1998; pp. 97–100. [Google Scholar]
- Khan, M.H.; Parvez, S. Hesperidin ameliorates heavy metal induced toxicity mediated by oxidative stress in brain of Wistar rat. J. Trace Elem. Med. Biol. 2015, 31, 53–60. [Google Scholar] [CrossRef]
- Humason, G.L. Animal Tissue Techniques, 3rd ed.; WH Freeman and Company: New York, NY, USA, 1972. [Google Scholar]
- Zheng, S.M.; Zhang, Y.; Wang, L.Y.; Zhou, Y.L. Prevention and treatment of bacterial septicemia in Carassius auratus with traditional Chinese medicine compound and its effect on immunity. J. Dalian Fish. Univ. 2006, 21, 31–36. (In Chinese) [Google Scholar]
- Chen, X.N. Experimental study on traditional Chinese medicine for the prevention and treatment of Aeromonas hydrophila infection in carp. Freshw. Fish. 2001, 31, 43–45. (In Chinese) [Google Scholar]
- Yang, S.Q.; Yang, G.; Guo, Z.F.; Li, Y.M. Study on the prevention and treatment of bacterial sepsis in grass carp with traditional Chinese medicine formula. Anhui Agric. Sci. 2010, 31, 17658–17659. (In Chinese) [Google Scholar]
- Goetz, F.W.; Planas, J.V.; MacKenzie, S. Tumor necrosis factors. Dev. Comp. Immunol. 2004, 28, 487–497. [Google Scholar] [CrossRef]
- Hardie, L.J.; Chappell, L.H.; Secombes, C.J. Human tumor necrosis factor α influences rainbow trout Oncorhynchus mykiss leucocyte responses. Vet. Immunol. Immunopathol. 1994, 40, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Whyte, S.K. The innate immune response of finfish–a review of current knowledge. Fish Shellfish. Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Nisha Rani, M.; Balasundaram, C. Hematological and biochemical parameters in common carp, Cyprinus carpio, following herbal treatment for Aeromonas hydrophila infection. Aquaculture 2003, 221, 41–50. [Google Scholar] [CrossRef]
- Sneath, P.H.A.; Mair, N.S.; Sharpe, M.E.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Whitman, W.B. Bergey’s Manual of Systematic Bacteriology; Williams & Wilkins: Philadelphia, PA, USA, 1986; Volume 2. [Google Scholar]
- Chen, X.Y.; Hou, Y.Y. Research progress on cytokines in fish. J. Hydrobiol. 2005, 28, 668–673. [Google Scholar]
- Muller-Eberhard, H.J. Molecular organization and function of the complement system. Annu. Rev. Biochem. 1988, 57, 321–347. [Google Scholar] [CrossRef]
- Liu, C.H.; Chiu, C.H.; Wang, S.W.; Cheng, W. Dietary administration of the probiotic, Bacillus subtilis E20, enhances the growth, innate immune responses, and disease resistance of the grouper, Epinephelus coioides. Fish Shellfish. Immunol. 2012, 33, 699–706. [Google Scholar] [CrossRef]
- Zhu, G.X.; Bai, D.Q.; Li, Y.H.; Ma, J.; Wu, X.; Ning, B. Preliminary study of 12 Chinese herbs as feed attractants on Turbot Juvenile (Scophtham lusmaximus). Agric. Sci. Technol. 2010, 11, 115–116. [Google Scholar]
- Bagnyukovaa, T.V.; Lushchaka, O.V.; Storeyb, K.B.; Lushchak, V.I. Oxidative stress and antioxidant defense responses by goldfish tissues to acute change of temperature from 3 to 23 °C. J. Therm. Biol. 2007, 32, 227–234. [Google Scholar] [CrossRef]
- Holmblad, T.; Soderhall, K. Cell adhesion molecules and anti-oxidative enzymes in a crustacean; possible role in immunity. Aquaculture 1999, 172, 111–123. [Google Scholar] [CrossRef]
- Lopes, P.A.; Pinheiro, T.; Santos, M.C.; Daluzmathias, M.; Collarespereira, M.; Viegascrespo, A. Response of antioxidant enzymes in freshwater fish populations (Leuciscus alburnoides complex) to inorganic pollutants exposure. Sci. Total Environ. 2001, 280, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Christybapita, D.; Divyagnaneswari, M.; Michael, R.D. Oral administration of Eclipta alba leaf aqueous extract enhances the non-specific immune responses and disease resistance of Oreochromis mossambicus. Fish Shellfish. Immunol. 2007, 23, 840–852. [Google Scholar] [CrossRef]
- Jing, X.; Ueki, N.; Cheng, J.; Imanishi, H.; Hada, T. Induction of apoptosis in hepatocellular carcinoma cell lines by emodin. Cancer Sci. 2002, 93, 874–882. [Google Scholar] [CrossRef]
- Jiang, S.; Song, M.J.; Shin, E.C.; Lee, M.O.; Kim, S.J.; Park, J.H. Apoptosis in human hepatoma cell lines by chemotherapeutic drugs via fas-dependent and fas-independent pathways. Hepatology 1999, 29, 101–110. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Percentage Dry Weight |
|---|---|
| Fish meal | 8 |
| Soybean meal | 18 |
| Rapeseed meal | 17 |
| Cotton meal | 16.5 |
| Rice bran | 8 |
| Wheat middling | 22 |
| Soybean oil | 4 |
| Lecithin | 1 |
| Choline chloride | 0.5 |
| Vitamin premix a | 1 |
| Mineral premix b | 1 |
| Powdered zeolite | 1 |
| Calcium dihydrogen phosphate | 2 |
| Proximate composition (%) | |
| Crude protein | 31.27 |
| Crude lipid | 8.15 |
| Crude ash | 11.02 |
| Gross energy (kJ g−1) c | 16.32 |
| RS Medium * | (g/L) |
|---|---|
| L-Ornithine hydrochloride | 6.5 |
| L-Lysine hydrochloride | 5.0 |
| L-Cysteine hydrochloride | 0.3 |
| Maltose | 3.5 |
| Sodium thiosulfate | 6.8 |
| Ferric ammonium citrate | 0.8 |
| Sodium chloride | 5.0 |
| Sodium deoxycholate | 1.0 |
| Yeast extract | 3.0 |
| Bromothymol blue | 0.03 |
| novobiocin | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.-Y.; Xu, P.; Wang, X.-L.; Song, L.-P.; Wu, J.; Wang, B.-L.; Hu, B.; Mao, S.-Q.; Liu, B.; Ge, X.-P. Study of Dietary Emodin on Immune Defense in Megalobrama amblycephala against Aeromonas hydrophila. Vet. Sci. 2023, 10, 588. https://doi.org/10.3390/vetsci10090588
Zhang Y-Y, Xu P, Wang X-L, Song L-P, Wu J, Wang B-L, Hu B, Mao S-Q, Liu B, Ge X-P. Study of Dietary Emodin on Immune Defense in Megalobrama amblycephala against Aeromonas hydrophila. Veterinary Sciences. 2023; 10(9):588. https://doi.org/10.3390/vetsci10090588
Chicago/Turabian StyleZhang, Yuan-Yuan, Peng Xu, Xiao-Li Wang, Li-Ping Song, Jun Wu, Bing-Li Wang, Bin Hu, Shu-Quan Mao, Bo Liu, and Xian-Ping Ge. 2023. "Study of Dietary Emodin on Immune Defense in Megalobrama amblycephala against Aeromonas hydrophila" Veterinary Sciences 10, no. 9: 588. https://doi.org/10.3390/vetsci10090588
APA StyleZhang, Y.-Y., Xu, P., Wang, X.-L., Song, L.-P., Wu, J., Wang, B.-L., Hu, B., Mao, S.-Q., Liu, B., & Ge, X.-P. (2023). Study of Dietary Emodin on Immune Defense in Megalobrama amblycephala against Aeromonas hydrophila. Veterinary Sciences, 10(9), 588. https://doi.org/10.3390/vetsci10090588