


Urinary Proteins of Female Domestic Dog (Canis familiaris) during Ovarian Cycle
 ,
,  , ,
, ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Animals
2.2. Sample Collection
Urine Sampling
- One animal in estrus only; single sample (Female 1).
- Two animals in estrus; two samples each (Female 2 and Female 3):
- ○
- One estrus sample of Female 3 contributed to pool P1.
- ○
- Additional sample of Female 2 in anestrus, out of which a portion was analyzed independently and the remaining contributed to pool P2.
- One animal in estrus and anestrus; one sample each (Female 4):
- ○
- Estrus sample was used as part of pool P1.
- ○
- Due to low volume, anestrus sample was used as part of pool P2.
- Four immature ones (constituting experimental pools P3: Female 5 and Female 6; P4: Female 7 and Female 8).
- Exact sample coding is described in Supplementary Materials along with the specific protein results. Sample scheme is also further described in Table 1.
2.3. Determination of the Phases of Cycle
2.3.1. Vaginal Cytological Examination
2.3.2. Progesterone Level Evaluation
2.4. Proteomic Approach
2.4.1. BCA Assay
2.4.2. Sample Pooling
2.4.3. Reduction, Alkylation, Digestion, and Tandem Mass Tag (TMT) Labeling
2.4.4. LC–MS/MS
2.4.5. Protein Identification and Quantification
2.5. Statistical Methods
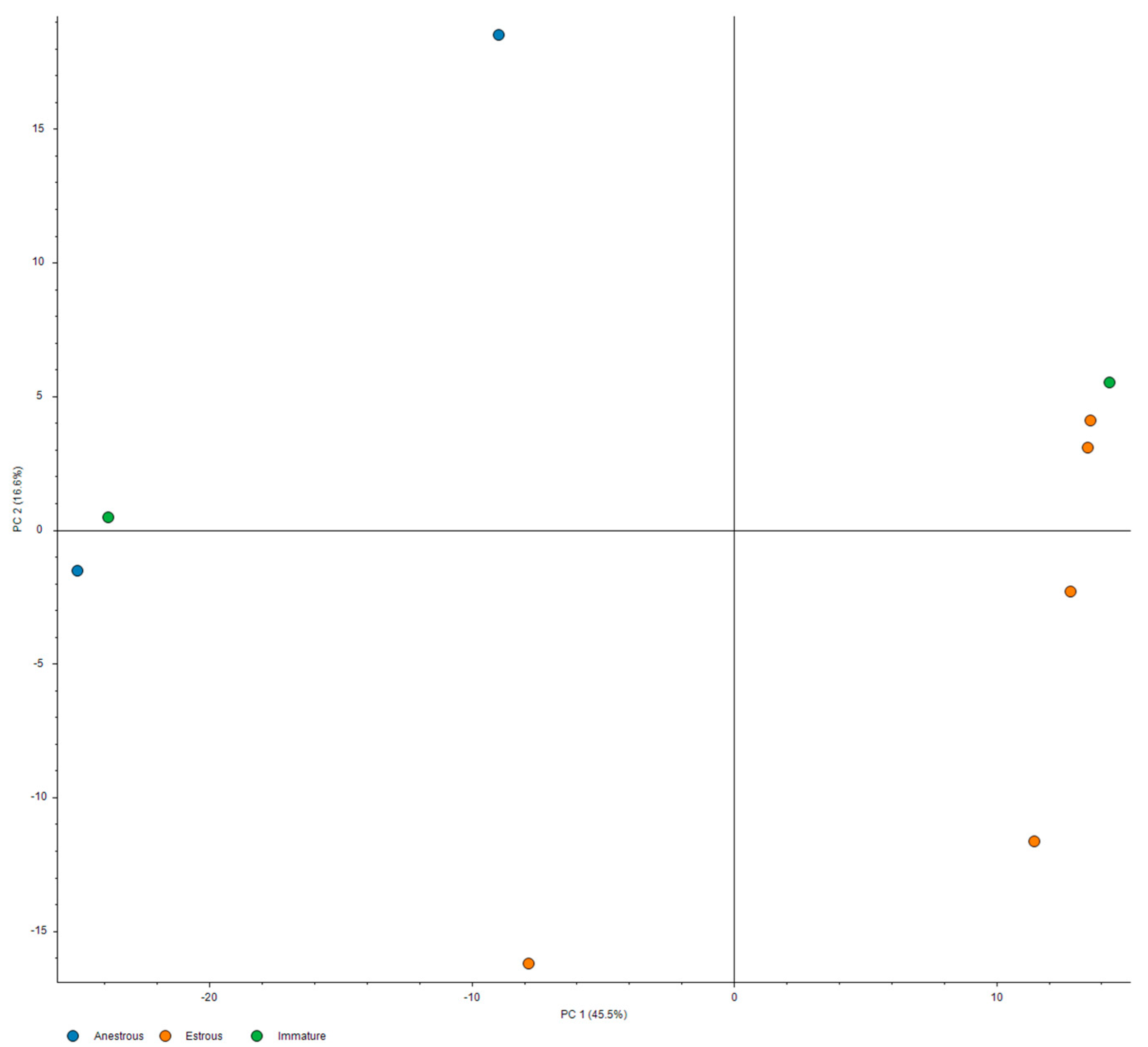
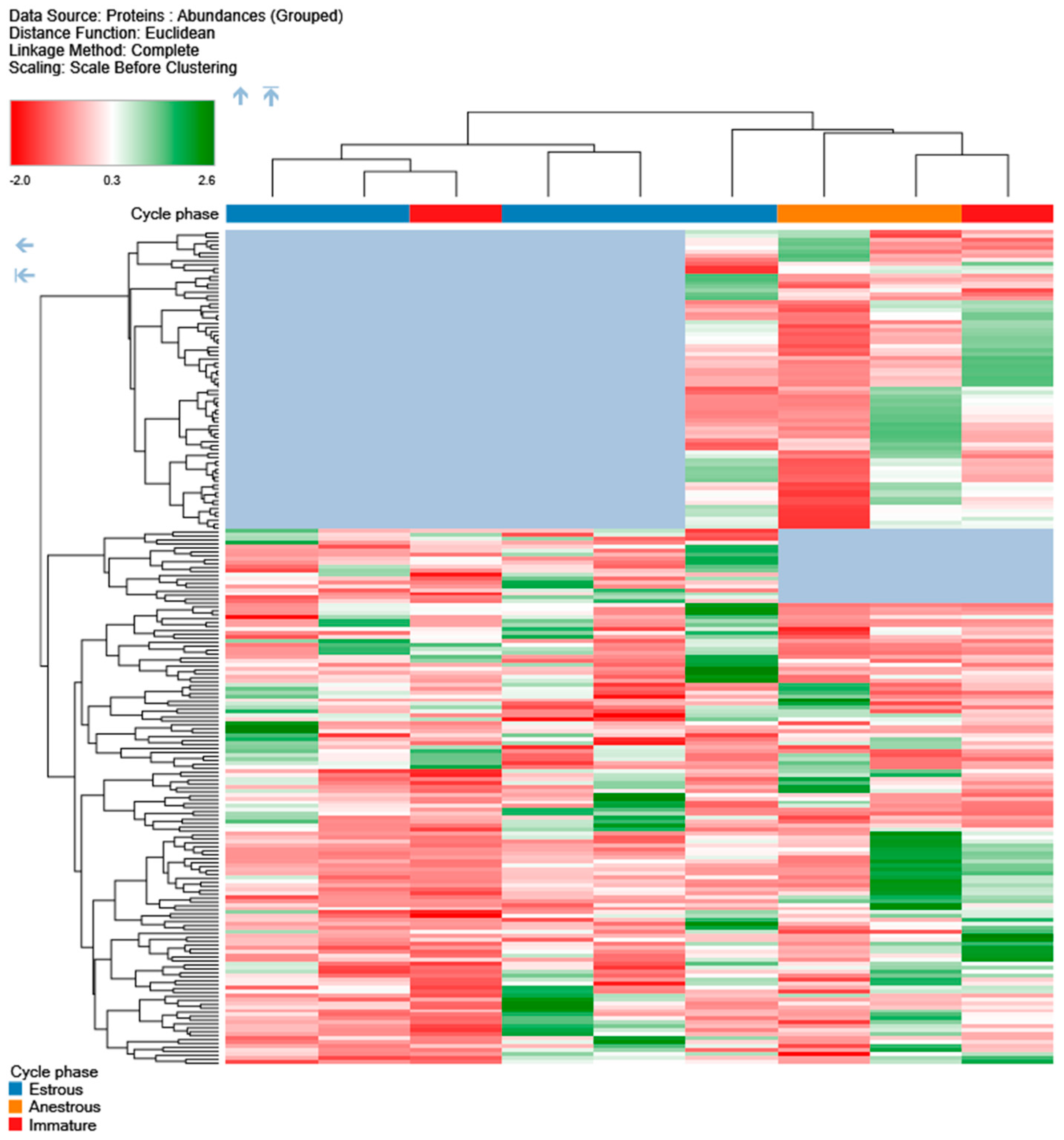
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signatures, 2nd ed.; Cambridge University Press, Ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Kokocińska-Kusiak, A.; Woszczyło, M.; Zybala, M.; Maciocha, J.; Barłowska, K.; Dzięcioł, M. Canine Olfaction: Physiology, Behavior, and Possibilities for Practical Applications. Animals 2021, 11, 2463. [Google Scholar] [CrossRef] [PubMed]
- Anisko, J.J. Hormonal Substrate of Estrus Odor Preference in Beagles. Physiol. Behav. 1977, 18, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Kruse, S.M.; Howard, W.E. Canid Sex Attractant Studies. J. Chem. Ecol. 1983, 9, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Kustritz, M.V.R. Collection of Tissue and Culture Samples from the Canine Reproductive Tract. Theriogenology 2006, 66, 567–574. [Google Scholar] [CrossRef]
- Dzięcioł, M.; Stańczyk, E.; Noszczyk-Nowak, A.; Niżański, W.; Ochota, M.; Kozdrowski, R. Influence of Bitches Sex Pheromones on the Heart Rate and Other Chosen Parameters of Blood Flow in Stud Dogs (Canis Familiaris). Res. Vet. Sci. 2012, 93, 1241–1247. [Google Scholar] [CrossRef]
- Jezierski, T.; Dzięcioł, M.; Szumny, A.; Niżański, W.; Woszczyło, M.; Pieczewska, B.; Godzińska, E.J. Discrimination of Estrus Odor in Urine by Male Dogs in Different Experimental Settings. J. Vet. Behav. 2019, 29, 25–30. [Google Scholar] [CrossRef]
- Goodwin, M.; Gooding, K.M.; Regnier, F. Sex Pheromone in the Dog. Science 1979, 203, 559–561. [Google Scholar] [CrossRef]
- Dzięcioł, M.; Niżański, W.; Jezierski, T.; Szumny, A.; Godzińska, E.J.; Ochota, M.; Stańczyk, E.; Najder-Kozdrowska, L.; Woszczyło, M.; Pieczewska, B. The Efficiency of Synthetic Sex Pheromones in Sexual Arousal Stimulation in Domestic Dogs. Pol. J. Vet. Sci. 2017, 20, 429–437. [Google Scholar] [CrossRef]
- Dzięcioł, M.; Woszczylo, M.; Szumny, A.; Jezierski, T.; Kupczyński, R.; Godzińska, E.J.; Pieczewska, B.; Niżański, W. Identification of Putative Volatile Sex Pheromones in Female Domestic Dogs (Canis Familiaris). Anim. Reprod. Sci. 2018, 197, 87–92. [Google Scholar] [CrossRef]
- Woszczyło, M.; Jezierski, T.; Szumny, A.; Dzięcioł, M.; Niżański, W. The Role of Urine in Semiochemical Communication between Females and Males of Domestic Dog (Canis Familiaris) during Estrus. Animals 2020, 10, 2112. [Google Scholar] [CrossRef]
- Dzięcioł, M.; Podgórski, P.; Stańczyk, E.; Szumny, A.; Woszczyło, M.; Pieczewska, B.; Niżański, W.; Nicpoń, J.; Wrzosek, M.A. MRI Features of the Vomeronasal Organ in Dogs (Canis Familiaris). Front. Vet. 2020, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Stowers, L. Bespoke Behavior: Mechanisms That Modulate Pheromone-Triggered Behavior. Curr. Opin. Neurobiol. 2020, 64, 143–150. [Google Scholar] [CrossRef] [PubMed]
- McGlone, J.J.; Archer, C.; Henderson, M. Interpretive Review: Semiochemicals in Domestic Pigs and Dogs. Front. Vet. Sci. 2022, 9, 967980. [Google Scholar] [CrossRef] [PubMed]
- Marchlewska-Koj, A. Pheromonal Regulation of Bank Vole (Clethrionomys Glareolus) Reproduction. In Chemical Signals in Vertebrates 9; Springer: Boston, MA, USA, 2001; pp. 391–396. ISBN 9781461351870. [Google Scholar]
- Rasmussen; Schulte, B.; Goodwin, T.; Whitehouse, A.; Loizi, H. Sexual Dimorphism in the Performance of Chemosensory Investigatory Behaviours by African Elephants (Loxodonta Africana). Behaviour 2009, 146, 373–392. [Google Scholar] [CrossRef]
- Beynon, R.J.; Hurst, J.L. Multiple Roles of Major Urinary Proteins in the House Mouse, Mus Domesticus. Biochem. Soc. Trans. 2003, 31, 142–146. [Google Scholar] [CrossRef]
- Novotny, M.; Harvey, S.; Jemiolo, B.; Alberts, J. Synthetic Pheromones That Promote Inter-Male Aggression in Mice. Proc. Natl. Acad. Sci. USA 1985, 82, 2059–2061. [Google Scholar] [CrossRef]
- Mucignat, C.; Benati, D.; Righetti, C.; Zancanaro, C. High-Resolution Magnetic Resonance Spectroscopy of the Mouse Vomeronasal Organ. Chem. Senses 2004, 29, 693–696. [Google Scholar] [CrossRef]
- Logan, D.W.; Marton, T.F.; Stowers, L. Species Specificity in Major Urinary Proteins by Parallel Evolution. PLoS ONE 2008, 3, e3280. [Google Scholar] [CrossRef]
- Rajkumar, R.; Ilayaraja, R.; Liao, C.C.; Archunan, G.; Achiraman, S. Detection of Alpha (2u)-Globulin and Its Bound Putative Pheromones in the Preputial Gland of the Indian Commensal Rat (Rattus Rattus) Using Mass Spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 721–728. [Google Scholar] [CrossRef]
- Dehnhard, M.; Hildebrandt, T.; Knauf, T.; Ochs, A.; Ringleb, J.; Göritz, F. Chemical Signals in Giant Panda Urine (Ailuropoda Melanoleuca). In Chemical Signals in Vertebrates 10; Springer: Boston, MA, USA, 2006; pp. 110–117. ISBN 9780387251592. [Google Scholar]
- Rajanarayanan, S.; Archunan, G. Identification of Urinary Sex Pheromones in Female Buffaloes and Their Influence on Bull Reproductive Behaviour. Res. Vet. Sci. 2011, 91, 301–305. [Google Scholar] [CrossRef]
- Dehnhard, M.; Hildebrandt, T.B.; Meerheim, C.; Valentine, I.; Göritz, F. Chemical Signals in Giant Panda Urine (Ailuropoda Melanoleuca). In Chemical Signals in Vertebrates 13; Springer International Publishing: Cham, Switzerland, 2016; pp. 363–379. ISBN 9783319220253. [Google Scholar]
- Ramachandran, R.; Vinothkumar, A.; Sankarganesh, D.; Suriyakalaa, U.; Aathmanathan, V.S.; Kamalakkannan, S.; Nithya, V.; Angayarkanni, J.; Archunan, G.; Akbarsha, M.A.; et al. Detection of Estrus Biomarkers in the Body Exudates of Kangayam Cattle (Bos Indicus) from Interplay of Hormones and Behavioral Expressions. Domest. Anim. Endocrinol. 2020, 72, 106392. [Google Scholar] [CrossRef]
- Nagnan-Le Meillour, P.; Descamps, A.; Le Danvic, C.; Grandmougin, M.; Saliou, J.-M.; Klopp, C.; Milhes, M.; Bompard, C.; Chesneau, D.; Poissenot, K.; et al. Identification of Potential Chemosignals in the European Water Vole Arvicola Terrestris. Sci. Rep. 2019, 9, 18378. [Google Scholar] [CrossRef]
- Sankarganesh, D.; Suriyakalaa, U.; Ramachandran, R.; Achiraman, S.; Arunachalam, S.; Angayarkanni, J. Urinary Volatile Metabolomics as a Viable Alternative Diagnostic Tool for Polycystic Ovary Syndrome: An Exploratory Hypothesis. Med. Hypotheses 2019, 124, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Winiarczyk, D.; Michalak, K.; Adaszek, Ł.; Winiarczyk, M.; Winiarczyk, S. Urinary Proteome of Dogs with Kidney Injury during Babesiosis. BMC Vet. Res. 2019, 15, 439. [Google Scholar] [CrossRef]
- Ajayi, A.F.; Akhigbe, R.E. Staging of the Estrus Cycle and Induction of Estrus in Experimental Rodents: An Update. Fertil. Res. Pract. 2020, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Ferlizza, E.; Isani, G.; Dondi, F.; Andreani, G.; Vasylyeva, K.; Bellei, E.; Almeida, A.M.; Matzapetakis, M. Urinary Proteome and Metabolome in Dogs (Canis Lupus Familiaris): The Effect of Chronic Kidney Disease. J. Proteom. 2020, 222, 103795. [Google Scholar] [CrossRef] [PubMed]
- Dubin, R.F.; Rhee, E.P. Proteomics and Metabolomics in Kidney Disease, Including Insights into Etiology, Treatment, and Prevention. Clin. J. Am. Soc. Nephrol. 2020, 15, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Miller, I.; Schlosser, S.; Palazzolo, L.; Veronesi, M.C.; Eberini, I.; Gianazza, E. Some More about Dogs: Proteomics of Neglected Biological Fluids. J. Proteom. 2020, 218, 103724. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Gatlin, C.L.; Makusky, A.J.; Russo, P.S.; Schatz, C.R.; Miller, S.S.; Su, Q.; McGrath, A.M.; Estock, M.A.; Parmar, P.P.; et al. The Human Serum Proteome: Display of Nearly 3700 Chromatographically Separated Protein Spots on Two-Dimensional Electrophoresis Gels and Identification of 325 Distinct Proteins. Proteomics 2003, 3, 1345–1364. [Google Scholar] [CrossRef]
- Miller, I.; Friedlein, A.; Tsangaris, G.; Maris, A.; Fountoulakis, M.; Gemeiner, M. The Serum Proteome of Equus Caballus. Proteomics 2004, 4, 3227–3234. [Google Scholar] [CrossRef] [PubMed]
- Presslmayer, A. Identifizierung Caniner Serumproteine Mittels Elektrophoretischer Verfahren. Master’s Thesis, University of Natural Resources and Applied Life Science Sciences, Vienna, Austria, 2002. [Google Scholar]
- Atherton, M.J.; Braceland, M.; Harvie, J.; Burchmore, R.J.; Eadie, S.; Eckersall, P.D.; Morris, J.S. Characterisation of the Normal Canine Serum Proteome Using a Novel Electrophoretic Technique Combined with Mass Spectrometry. Vet. J. 2013, 196, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Atherton, M.J.; Braceland, M.; Fontaine, S.; Waterston, M.M.; Burchmore, R.J.; Eadie, S.; Eckersall, P.D.; Morris, J.S. Changes in the Serum Proteome of Canine Lymphoma Identified by Electrophoresis and Mass Spectrometry. Vet. J. 2013, 196, 320–324. [Google Scholar] [CrossRef]
- Szczubiał, M.; Wawrzykowski, J.; Dąbrowski, R.; Krawczyk, M.; Kankofer, M. Preliminary Study on Plasma Proteins in Pregnant and Non-Pregnant Female Dogs. Theriogenology 2017, 97, 1–8. [Google Scholar] [CrossRef]
- Dąbrowski, R.; Kostro, K.; Szczubiał, M. Concentrations of C-Reactive Protein, Serum Amyloid A, and Haptoglobin in Uterine Arterial and Peripheral Blood in Bitches with Pyometra. Theriogenology 2013, 80, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Franco-Martínez, L.; Horvatić, A.; Gelemanović, A.; Samardžija, M.; Mrljak, V.; Contreras-Aguilar, M.D.; Martínez-Subiela, S.; Dabrowski, R.; Tvarijonaviciute, A. Changes in the Salivary Proteome Associated with Canine Pyometra. Front. Vet. Sci. 2020, 7, 277. [Google Scholar] [CrossRef] [PubMed]
- Miller, I.; Preßlmayer-Hartler, A.; Wait, R.; Hummel, K.; Sensi, C.; Eberini, I.; Razzazi-Fazeli, E.; Gianazza, E. In between—Proteomics of Dog Biological Fluids. J. Proteom. 2014, 106, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Teinfalt, M.; Miller, I.; Loupal, G.; Thalhammer, J.G.; Gemeiner, M. Quantitative Determination of a Canine Prostate Specific Protein and Its Clinical Relevance. Tierärztliche Praxis. Ausgabe K Kleintiere/Heimtiere 2000, 28, 127–131. [Google Scholar]
- Kustritz, M.V.R. Reproductive Behavior of Small Animals. Theriogenology 2005, 64, 734–746. [Google Scholar] [CrossRef]
- Arlt, S. Canine Ovulation Timing: A Survey on Methodology and an Assessment on Reliability of Vaginal Cytology. Reprod. Domest. Anim. 2018, 53 (Suppl. S3), 53–62. [Google Scholar] [CrossRef]
- Brugger, N.; Otzdorff, C.; Walter, B.; Hoffmann, B.; Braun, J. Quantitative Determination of Progesterone (P4) in Canine Blood Serum Using an Enzyme-Linked Fluorescence Assay. Reprod. Domest. Anim. 2011, 46, 870–873. [Google Scholar] [CrossRef]
- Orsburn, B.C. Proteome Discoverer-A Community Enhanced Data Processing Suite for Protein Informatics. Proteomes 2021, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, S.; Rajesh, D.; Saibaba, G.; Alagesan, A.; Rengarajan, R.L.; Archunan, G. Urinary Lipocalin Protein in a Female Rodent with Correlation to Phases in the Estrus Cycle: An Experimental Study Accompanied by in Silico Analysis. PLoS ONE 2013, 8, e371357. [Google Scholar] [CrossRef]
- Virtanen, T.; Zeiler, T.; Mäntyjärvi, R. Important Animal Allergens Are Lipocalin Proteins: Why Are They Allergenic? Int. Arch. Allergy Immunol. 1999, 120, 247–258. [Google Scholar] [CrossRef]
- Rosenberg, M.E.; Silkensen, J. Clusterin: Physiologic and Pathophysiologic Considerations. Int. J. Biochem. Cell 1995, 27, 633–645. [Google Scholar] [CrossRef]
- Jones, S.E.; Jomary, C. Clusterin. Int. J. Biochem. Cell Biol. 2002, 34, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Jenne, D.E.; Tschopp, J. Clusterin: The Intriguing Guises of a Widely Expressed Glycoprotein. Trends Biochem. Sci. 1992, 17, 154–159. [Google Scholar] [CrossRef]
- Fahiminiya, S.; Reynaud, K.; Labas, V.; Batard, S.; Chastant-Maillard, S.; Gérard, N. Steroid Hormones Content and Proteomic Analysis of Canine Follicular Fluid during the Preovulatory Period. Reprod. Biol. Endocrinol. 2010, 8, 132. [Google Scholar] [CrossRef]
- Zwain, I.; Amato, P. Clusterin Protects Granulosa Cells from Apoptotic Cell Death during Follicular Atresia. Exp. Cell Res. 2000, 257, 101–110. [Google Scholar] [CrossRef]
- Jin, D.F.; Muffly, K.E.; Okulicz, W.C.; Kilpatrick, D.L. Estrus Cycle- and Pregnancy-Related Differences in Expression of the Proenkephalin and Proopiomelanocortin Genes in the Ovary and Uterus. Endocrinology 1988, 122, 1466–1471. [Google Scholar] [CrossRef]
- Muffly, K.E.; Jin, D.F.; Okulicz, W.C.; Kilpatrick, D.L. Gonadal Steroids Regulate Proenkephalin Gene Expression in a Tissue-Specific Manner within the Female Reproductive System. Mol. Endocrinol. 1988, 2, 979–985. [Google Scholar] [CrossRef]
- Konturek, S.J.; Tasler, J.; Krol, R.; Dembinski, A.; Coy, D.H.; Schally, A.V. Effect of Somatostatin Analogs on Gastric and Pancreatic Secretion. Proc. Soc. Exp. Biol. Med. 1977, 155, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Tan, C.C.; Craik, D.J.; Clark, R.J. Structural and Functional Analysis of Human Liver-Expressed Antimicrobial Peptide 2. Chembiochem 2010, 11, 2057–2148. [Google Scholar] [CrossRef]
- Valdez-Miramontes, C.E.; De Haro-Acosta, J.; Aréchiga-Flores, C.F.; Verdiguel-Fernández, L.; Rivas-Santiago, B. Antimicrobial Peptides in Domestic Animals and Their Applications in Veterinary Medicine. Peptides 2021, 142, 170576. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.E.; Dagnall, G.J.R. Some Observations on the Aerobic Bacterial Flora of the Genital Tract of the Dog and Bitch. J. Small Anim. Pract. 1982, 23, 325–335. [Google Scholar] [CrossRef]
- Maksimović, A.; Maksimović, Z.; Filipović, S.; Beširović, H.; Rifatbegović, M. Vaginal and Uterine Bacteria of Healthy Bitches during Different Stages of Their Reproductive Cycle. Vet. Rec. 2012, 171, 375. [Google Scholar] [CrossRef]
- Merkx, J.; Slob, A.K.; van der Werff ten Bosch, J.J. Vaginal Bacterial Flora Partially Determines Sexual Attractivity of Female Rats. Physiol. Behav. 1988, 44, 147–149. [Google Scholar] [CrossRef]
- Ungerfeld, R.; Silva, L. The Presence of Normal Vaginal Flora Is Necessary for Normal Sexual Attractiveness of Estrus Ewes. Appl. Anim. Behav. Sci. 2005, 93, 245–250. [Google Scholar] [CrossRef]
- Dzięcioł, M.; Niżański, W.; Stańczyk, E.; Kozdrowski, R.; Najder-Kozdrowska, L.; Twardoń, J. The Influence of Antibiotic Treatment of Bitches in Estrus on Their Attractiveness to Males during Mating. Pol. J. Vet. Sci. 2013, 16, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Bienboire-Frosini, C.; Durairaj, R.; Pelosi, P.; Pageat, P. The Major Cat Allergen Fel d 1 Binds Steroid and Fatty Acid Semiochemicals: A Combined in Silico and in Vitro Study. Int. J. Mol. Sci. 2020, 21, 1365. [Google Scholar] [CrossRef]
- Krishna, N.S.; Getchell, M.L.; Margolis, F.L.; Getchell, T.V. Differential Expression of Vomeromodulin and Odorant-Binding Protein, Putative Pheromone and Odorant Transporters, in the Developing Rat Nasal Chemosensory Mucosae. J. Neurosci. Res. 1995, 40, 54–71. [Google Scholar] [CrossRef]
- Janssenswillen, S.; Roelants, K.; Carpentier, S.; de Rooster, H.; Metzemaekers, M.; Vanschoenwinkel, B.; Proost, P.; Bossuyt, F. Odorant-Binding Proteins in Canine Anal Sac Glands Indicate an Evolutionarily Conserved Role in Mammalian Chemical Communication. BMC Ecol. Evol. 2021, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P. The Role of Perireceptor Events in Vertebrate Olfaction. Cell. Mol. Life Sci. 2001, 58, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Archunan, G. Odorant Binding Proteins: A Key Player in the Sense of Smell. Bioinformation 2018, 14, 036–037. [Google Scholar] [CrossRef]
- Pelosi, P.; Knoll, W. Odorant-Binding Proteins of Mammals. Biol. Rev. Camb. Philos. Soc. 2022, 97, 20–44. [Google Scholar] [CrossRef]
- Mitchell, G.B.; Clark, M.E.; Lu, R.; Caswell, J.L. Localization and Functional Characterization of Pulmonary Bovine Odorant-Binding Protein. Vet. Pathol. 2011, 48, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- Černá, M.; Kuntová, B.; Talacko, P.; Stopková, R.; Stopka, P. Differential Regulation of Vaginal Lipocalins (OBP, MUP) during the Estrus Cycle of the House Mouse. Sci. Rep. 2017, 7, 11674. [Google Scholar] [CrossRef] [PubMed]
- Ulutas, P.A.; Musal, B.; Kiral, F.; Bildik, A. Acute Phase Protein Levels in Pregnancy and Estrus Cycle in Bitches. Res. Vet. Sci. 2009, 86, 373–376. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Individual Samples | Pooled Samples | ||||
|---|---|---|---|---|---|
| Sample Number | Dog | Ovarian Cycle Phase | Sample Number | Dogs | Ovarian Cycle Phase |
| S1 | Female2 | Estrus | P1 | Female3, Female4 | Estrus |
| S4 | Female1 | Estrus | P2 | Female3, Female4 | Anestrus |
| S5 | Female3 | Estrus | P3 | Female5, Female6 | Immature |
| S6 | Female2 | Estrus | P4 | Female7, Female8 | Immature |
| S7 | Female3 | Anestrus | |||
| Annotation | Description | Count | p-Value | BH p-Value |
|---|---|---|---|---|
| Annotation Cluster 1 | ||||
| UP_KW_DOMAIN | Signal | 8 | 1.5 × 10−2 | 7.7 × 10−1 |
| GOTERM_CC_FAT | extracellular region | 6 | 1.6 × 10−2 | 3.9 × 10−1 |
| UP_KW_CELLULAR_COMPONENT | Secreted | 4 | 1.7 × 10−2 | 1.7 × 10−1 |
| GOTERM_BP_FAT | response to other organism | 3 | 8.2 × 10−2 | 1.0 |
| GOTERM_BP_FAT | response to external biotic stimulus | 3 | 8.3 × 10−2 | 1.0 |
| GOTERM_BP_FAT | response to biotic stimulus | 3 | 9.1 × 10−2 | 1.0 |
| GOTERM_BP_FAT | response to external stimulus | 4 | 1.2 × 10−1 | 1.0 |
| UP_KW_PTM | Disulfide bond | 5 | 1.4 × 10−1 | 9.7 × 10−1 |
| Annotation Cluster 2 | ||||
| GOTERM_CC_FAT | extracellular region | 6 | 1.6 × 10−2 | 3.9 × 10−1 |
| GOTERM_BP_FAT | cellular amide metabolic process | 4 | 4.7 × 10−2 | 1.0 |
| GOTERM_CC_FAT | extracellular space | 4 | 7.5 × 10−1 | 1.0 |
| GOTERM_CC_FAT | extracellular region part | 4 | 1.2 × 10−1 | 1.0 |
| UP_KW_PTM | Glycoprotein | 3 | 7.4 × 10−1 | 1.0 |
| NCBI Gene ID | Protein Name | Relevant GO Term | Abundance Change (Estrus vs. Anestrus) |
|---|---|---|---|
| 442971 | Clusterin (CLU) | Smoothened signaling pathway GO:0007224 | 2.153 |
| 609750 | Liver-expressed antimicrobial peptide 2 (LEAP2) | Antimicrobial humoral immune response mediated by antimicrobial peptide GO:0061844 | 1000 |
| 100687307 | Proenkephalin (PENK) | signal transduction GO:0007165 sensory perception GO:0007600 | 1000 |
| NCBI Gene ID | Protein Name | Relevant GO Term | Abundance Change (Estrus vs. Anestrus) |
|---|---|---|---|
| 442971 | Clusterin (CLU) | Smoothened signaling pathway GO:0007224 | 2.153 |
| 609750 | Liver-expressed antimicrobial peptide 2 (LEAP2) | Antimicrobial humoral immune response mediated by antimicrobial peptide GO:0061844 | 1000 |
| 487447 | MBL associated serine protease 2 (MASP2) | cell surface pattern recognition receptor signaling pathway GO:0006958 complement activation, classical pathway GO:0002752 positive regulation of opsonization GO:1903028 | 1000 |
| 100687307 | Proenkephalin (PENK) | signal transduction GO:0007165 | 1000 |
| NCBI Gene ID | Protein Name | Relevant GO Term | Abundance Change (Estrus vs. Anestrus) |
|---|---|---|---|
| 403831 | Beta glucuronidase (GUSB) | Enzyme. Plays an important role in the degradation of dermatan and keratan sulfates. Present in lysosomes (very much inside the cell, not secreted). | 0.432 |
| 490630 | Granzyme B (GZMB) | Cytosolic enzyme released by lymphocytes to kill virus-infected cells. Not signaling. | 2.147 |
| 609112 | Lymphocyte antigen 6 family member D (LY6D) | Involved in leukocyte differentiation, affected by stilbenoid, present on the cell surface, but not really secreted or signaling. GO:1900740: positive regulation of protein insertion into mitochondrial membrane involved in apoptotic signaling pathway GO:0017148: negative regulation of translation. | 1000 |
| 489503 | Myosin heavy chain 13 (MYH13) | Myosin is a structural protein involved in muscle contractions. | 0.396 |
| 476279 | Sphingomyelin phosphodiesterase acid-like 3A (SMPDL3A) | Enzyme regulating levels of cyclic AMP. | 1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woszczyło, M.; Pasikowski, P.; Devaraj, S.; Kokocińska, A.; Szumny, A.; Skwark, M.J.; Niżański, W.; Dzięcioł, M. Urinary Proteins of Female Domestic Dog (Canis familiaris) during Ovarian Cycle. Vet. Sci. 2023, 10, 292. https://doi.org/10.3390/vetsci10040292
Woszczyło M, Pasikowski P, Devaraj S, Kokocińska A, Szumny A, Skwark MJ, Niżański W, Dzięcioł M. Urinary Proteins of Female Domestic Dog (Canis familiaris) during Ovarian Cycle. Veterinary Sciences. 2023; 10(4):292. https://doi.org/10.3390/vetsci10040292
Chicago/Turabian StyleWoszczyło, Martyna, Paweł Pasikowski, Sankarganesh Devaraj, Agata Kokocińska, Antoni Szumny, Marcin J. Skwark, Wojciech Niżański, and Michał Dzięcioł. 2023. "Urinary Proteins of Female Domestic Dog (Canis familiaris) during Ovarian Cycle" Veterinary Sciences 10, no. 4: 292. https://doi.org/10.3390/vetsci10040292
APA StyleWoszczyło, M., Pasikowski, P., Devaraj, S., Kokocińska, A., Szumny, A., Skwark, M. J., Niżański, W., & Dzięcioł, M. (2023). Urinary Proteins of Female Domestic Dog (Canis familiaris) during Ovarian Cycle. Veterinary Sciences, 10(4), 292. https://doi.org/10.3390/vetsci10040292