
Whole Genome Sequencing of SARS-CoV-2 in Cats and Dogs in South Korea in 2021

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Detection of SARS-CoV-2
2.2. Next-Generation Sequencing and Assembly
2.3. Genotyping and Phylogenetic Analysis
3. Results
3.1. SARS-CoV-2 Whole Genome Sequencing in Cats and Dogs
3.2. Molecular and Phylogenetic Characterization of Viral Genomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization 2022. WHO, COVID-19 Dashboard. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 31 October 2022).
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The biological and clinical significance of emerging SARS-CoV-2 variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef]
- Gupta, A.M.; Chakrabarti, J.; Mandal, S. Non-synonymous mutations of SARS-CoV-2 leads epitope loss and segregates its variants. Microbes Infect. 2020, 22, 598–607. [Google Scholar] [CrossRef]
- Sun, C.; Xie, C.; Bu, G.L.; Zhong, L.Y.; Zeng, M.S. Molecular characteristics, immune evasion, and impact of SARS-CoV-2 variants. Signal Transduct. Target Ther. 2022, 7, 202. [Google Scholar] [CrossRef] [PubMed]
- Maher, M.C.; Bartha, I.; Weaver, S.; di Iulio, J.; Ferri, E.; Soriaga, L.; Lempp, F.A.; Hie, B.L.; Bryson, B.; Berger, B.; et al. Predicting the mutational drivers of future SARS-CoV-2 variants of concern. Sci. Transl. Med. 2022, 14, eabk3445. [Google Scholar] [CrossRef]
- WOAH COVID-19 Dashboard. World Organisation for Animal Health. Available online: https://www.woah.org/en/what-we-offer/emergency-preparedness/covid-19/ (accessed on 31 October 2022).
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef]
- Global Initiative for Sharing All Influenza Data (GISAID). EiCoV Dashboard. Available online: https://www.epicov.org/ (accessed on 31 October 2022).
- Oude Munnink, B.B.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; van der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M.; et al. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef]
- Hammer, A.S.; Quaade, M.L.; Rasmussen, T.B.; Fonager, J.; Rasmussen, M.; Mundbjerg, K.; Lohse, L.; Strandbygaard, B.; Jorgensen, C.S.; Alfaro-Nunez, A.; et al. SARS-CoV-2 Transmission between Mink (Neovison vison) and Humans, Denmark. Emerg. Infect. Dis. 2021, 27, 547–551. [Google Scholar] [CrossRef]
- Larsen, H.D.; Fonager, J.; Lomholt, F.K.; Dalby, T.; Benedetti, G.; Kristensen, B.; Urth, T.R.; Rasmussen, M.; Lassauniere, R.; Rasmussen, T.B.; et al. Preliminary report of an outbreak of SARS-CoV-2 in mink and mink farmers associated with community spread, Denmark, June to November 2020. Euro Surveill. 2021, 26, 2100009. [Google Scholar] [CrossRef]
- Yen, H.L.; Sit, T.H.C.; Brackman, C.J.; Chuk, S.S.Y.; Gu, H.; Tam, K.W.S.; Law, P.Y.T.; Leung, G.M.; Peiris, M.; Poon, L.L.M.; et al. Transmission of SARS-CoV-2 delta variant (AY.127) from pet hamsters to humans, leading to onward human-to-human transmission: A case study. Lancet 2022, 399, 1070–1078. [Google Scholar] [CrossRef]
- Haagmans, B.L.; Koopmans, M.P.G. Spreading of SARS-CoV-2 from hamsters to humans. Lancet 2022, 399, 1027–1028. [Google Scholar] [CrossRef]
- Kok, K.H.; Wong, S.C.; Chan, W.M.; Wen, L.; Chu, A.W.; Ip, J.D.; Lee, L.K.; Wong, I.T.; Lo, H.W.; Cheng, V.C.; et al. Co-circulation of two SARS-CoV-2 variant strains within imported pet hamsters in Hong Kong. Emerg. Microbes Infect. 2022, 11, 689–698. [Google Scholar] [CrossRef]
- Han, T.; Ryu, B.; Lee, S.; Song, Y.; Jeong, Y.; Kim, I.; Kim, J.; Kim, E.; Lee, W.; Lee, H.; et al. Management following the first confirmed case of SARS-CoV-2 in a domestic cat associated with a massive outbreak in South Korea. One Health 2021, 13, 100328. [Google Scholar] [CrossRef]
- Bushnell, B. BBTools Software Package. 2014. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 1 September 2022).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing, S. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Neph, S.; Kuehn, M.S.; Reynolds, A.P.; Haugen, E.; Thurman, R.E.; Johnson, A.K.; Rynes, E.; Maurano, M.T.; Vierstra, J.; Thomas, S.; et al. BEDOPS: High-performance genomic feature operations. Bioinformatics 2012, 28, 1919–1920. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef]
- Rambaut, A.; Holmes, E.C.; O’Toole, A.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef]
- O’Toole, A.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Animal Protection Management System, Ministry of Agriculture and Rural Affairs. Policy Promotion Dashboard. Available online: https:animal.go.kr/ (accessed on 18 December 2022).
- Korea Disease Control and Prevention Agency (KDCA), Republic of Korea. COVID-19 Dashboard. Available online: https://ncov.kdca.go.kr/ (accessed on 12 December 2022).
- Pango Lineages: Latest Epidemiological Lineages of SARS-CoV-2. Lineage List Dashboard. Available online: https://www.cov-lineages.org/ (accessed on 31 October 2022).
- Hamer, S.A.; Ghai, R.R.; Zecca, I.B.; Auckland, L.D.; Roundy, C.M.; Davila, E.; Busselman, R.E.; Tang, W.; Pauvolid-Correa, A.; Killian, M.L.; et al. SARS-CoV-2 B.1.1.7 variant of concern detected in a pet dog and cat after exposure to a person with COVID-19, USA. Transbound. Emerg. Dis. 2022, 69, 1656–1658. [Google Scholar] [CrossRef]
- Park, A.K.; Kim, I.H.; Man Kim, H.; Lee, H.; Lee, N.J.; Kim, J.A.; Woo, S.; Lee, C.Y.; Lee, J.; Oh, S.J.; et al. SARS-CoV-2 B.1.619 and B.1.620 Lineages, South Korea, 2021. Emerg. Infect. Dis. 2022, 28, 415–419. [Google Scholar] [CrossRef]
- Kwon, J.H.; Kim, J.M.; Lee, D.H.; Park, A.K.; Kim, I.H.; Kim, D.W.; Kim, J.Y.; Lim, N.; Cho, K.Y.; Kim, H.M.; et al. Genomic epidemiology reveals the reduction of the introduction and spread of SARS-CoV-2 after implementing control strategies in Republic of Korea, 2020. Virus Evol. 2021, 7, veab077. [Google Scholar] [CrossRef]
- Kim, I.H.; Park, A.K.; Lee, H.J.; Kim, H.M.; Kim, K.J.; Kim, J.A.; Noh, J.S.; Lee, C.Y.; Rhee, J.E.; Kim, E.J. Status and characteristics of the SARS-CoV-2 variant outbreak in the Republic of Korea in January 2021. Public Health Wkly Rep. (Korea Disease Control and Prevention Agency) 2022, 15, 497–509. [Google Scholar]


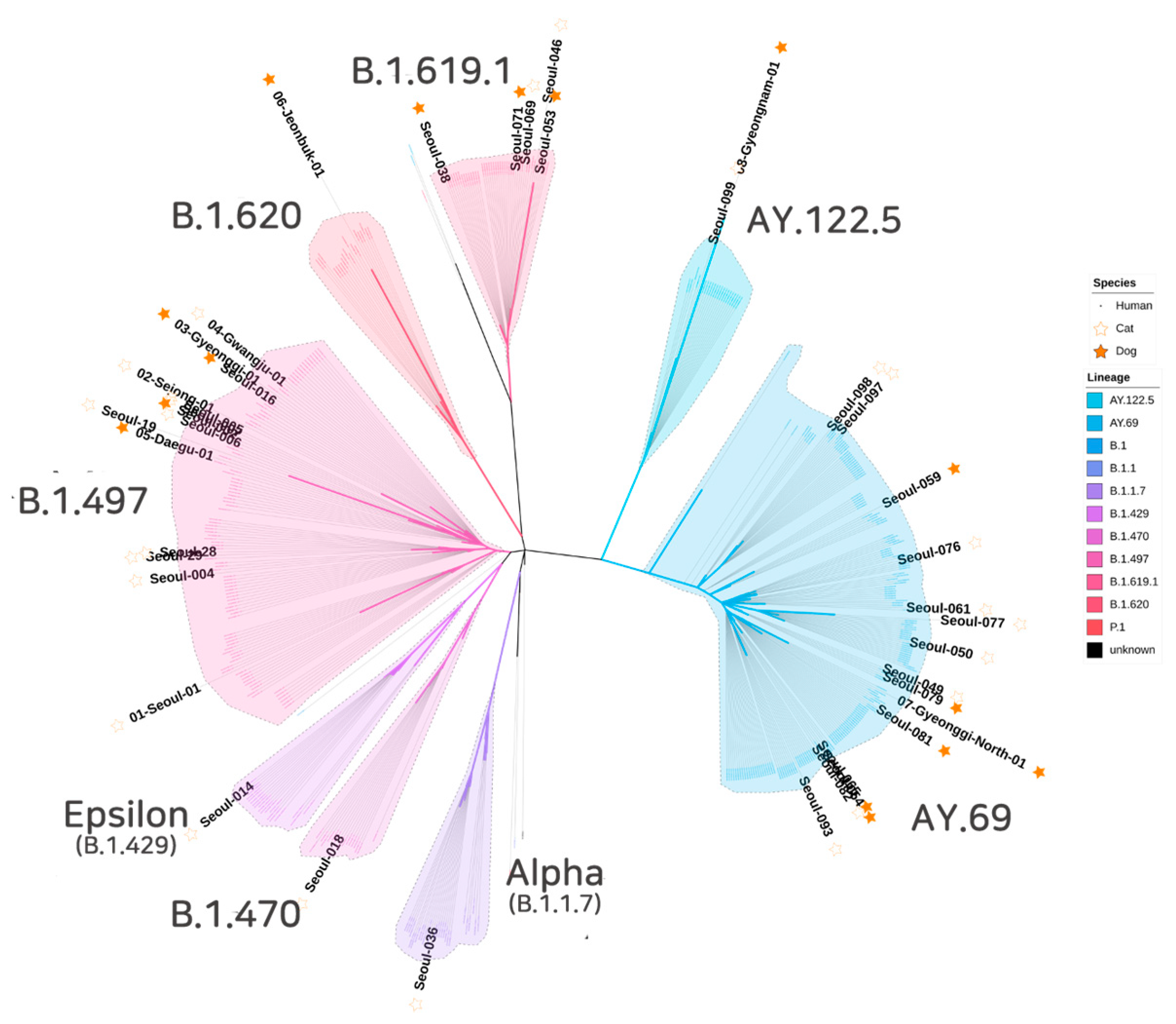{kind=link}
| Pango Lineage | Cat | (%) * | Dog | (%) * | Total | (%) * | |
|---|---|---|---|---|---|---|---|
| B.1.497 | 10 | 40.0 | 4 | 26.7 | 14 | 35.0 | |
| B.1.429 | (Epsilon variant) | 1 | 4.0 | - | 0.0 | 1 | 2.5 |
| B.1.470 | 1 | 4.0 | - | 0.0 | 1 | 2.5 | |
| B.1.620 | - | 0.0 | 1 | 6.7 | 1 | 2.5 | |
| B.1.1.7 | (Alpha variant) | 1 | 4.0 | - | 0.0 | 1 | 2.5 |
| B.1.619.1 | 2 | 8.0 | 3 | 20.0 | 5 | 12.5 | |
| AY.69 | (Delta variant) | 9 | 36.0 | 6 | 40.0 | 15 | 37.5 |
| AY.122.5 | (Delta variant) | 1 | 4.0 | 1 | 6.7 | 2 | 5.0 |
| Total | 25 | 15 | 40 |
| No. | Species | Strain | Sampling Date | Pango Lineage | Nextstrain Clade † | GISAID Clade ‡ | WHO Label | GISAID Accession ID. |
|---|---|---|---|---|---|---|---|---|
| 1 | cat | 01-Seoul-01 | 2021-02-13 | B.1.497 | 20C | GH | – | EPI_ISL_15775580 |
| 2 | cat | 02-Sejong-01 | 2021-02-16 | B.1.497 | 20C | GH | – | EPI_ISL_15775581 |
| 3 | dog | Seoul-002 | 2021-02-16 | B.1.497 | 20C | GH | – | EPI_ISL_15775592 |
| 4 | dog | 03-Gyeonggi-01 | 2021-02-19 | B.1.497 | 20C | GH | – | EPI_ISL_15788906 |
| 5 | cat | 04-Gwangju-01 | 2021-03-05 | B.1.497 | 20C | GH | – | EPI_ISL_15793141 |
| 6 | cat | Seoul-004 | 2021-03-08 | B.1.497 | 20C | GH | – | EPI_ISL_15775582 |
| 7 | cat | Seoul-005 | 2021-03-09 | B.1.497 | 20C | GH | – | EPI_ISL_15727682 |
| 8 | cat | Seoul-006 | 2021-03-09 | B.1.497 | 20C | GH | – | EPI_ISL_15727683 |
| 9 | cat | Seoul-007 | 2021-03-09 | B.1.497 | 20C | GH | – | EPI_ISL_15727684 |
| 10 | cat | Seoul-014 | 2021-03-26 | B.1.429 | 21C (Epsilon) | GH | Epsilon Variant | EPI_ISL_15727685 |
| 11 | dog | 05-Daegu-01 | 2021-03-30 | B.1.497 | 20C | GH | – | EPI_ISL_15727696 |
| 12 | dog | Seoul-016 | 2021-04-06 | B.1.497 | 20C | GH | – | EPI_ISL_15775593 |
| 13 | cat | Seoul-018 | 2021-04-11 | B.1.470 | 20A | GH | – | EPI_ISL_15727686 |
| 14 | cat | Seoul-19 | 2021-04-13 | B.1.497 | 20C | GH | – | EPI_ISL_15727687 |
| 15 | cat | Seoul-28 | 2021-05-04 | B.1.497 | 20C | GH | – | EPI_ISL_15727688 |
| 16 | cat | Seoul-29 | 2021-05-04 | B.1.497 | 20C | GH | – | EPI_ISL_15727689 |
| 17 | dog | 06-Jeonbuk-01 | 2021-05-28 | B.1.620 | 20A | G | – | EPI_ISL_15775590 |
| 18 | cat | Seoul-036 | 2021-06-04 | B.1.1.7 | 20I (Alpha, V1) | GRY | Alpha variant | EPI_ISL_15727690 |
| 19 | dog | Seoul-038 | 2021-06-06 | B.1.619.1 | 20A | GH | – | EPI_ISL_15775594 |
| 20 | cat | Seoul-046 | 2021-06-30 | B.1.619.1 | 20A | GH | – | EPI_ISL_15775583 |
| 21 | cat | Seoul-049 | 2021-07-02 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15727691 |
| 22 | cat | Seoul-050 | 2021-07-11 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15775584 |
| 23 | dog | Seoul-053 | 2021-07-13 | B.1.619.1 | 20A | GH | – | EPI_ISL_15727698 |
| 24 | dog | Seoul-054 | 2021-07-14 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15727699 |
| 25 | dog | Seoul-059 | 2021-07-20 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15727700 |
| 26 | cat | Seoul-061 | 2021-07-22 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15727692 |
| 27 | dog | Seoul-065 | 2021-07-27 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15793142 |
| 28 | cat | Seoul-069 | 2021-08-01 | B.1.619.1 | 20A | GH | – | EPI_ISL_15727693 |
| 29 | dog | Seoul-071 | 2021-08-03 | B.1.619.1 | 20A | GH | – | EPI_ISL_15727701 |
| 30 | cat | Seoul-076 | 2021-08-08 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15775585 |
| 31 | cat | Seoul-077 | 2021-08-08 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15727694 |
| 32 | dog | 07-Gyeonggi-North-01 | 2021-08-10 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15775591 |
| 33 | dog | Seoul-079 | 2021-08-10 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15775595 |
| 34 | dog | Seoul-081 | 2021-08-20 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15727702 |
| 35 | cat | Seoul-082 | 2021-08-24 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15727695 |
| 36 | cat | Seoul-093 | 2021-09-19 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15775586 |
| 37 | cat | Seoul-097 | 2021-10-08 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15775587 |
| 38 | cat | Seoul-098 | 2021-10-08 | AY.69 | 21I (Delta) | GK | Delta variant | EPI_ISL_15775588 |
| 39 | cat | Seoul-099 | 2021-10-24 | AY.122.5 | 21J (Delta) | GK | Delta variant | EPI_ISL_15775589 |
| 40 | dog | 08-Gyeongnam-01 | 2021-12-10 | AY.122.5 | 21J (Delta) | GK | Delta variant | EPI_ISL_15727697 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, Y.-K.; Kwon, O.-K.; Heo, J.; Nah, J.; Kang, H.-E.; Kang, Y.; Song, I.J.; Sung, H.-K. Whole Genome Sequencing of SARS-CoV-2 in Cats and Dogs in South Korea in 2021. Vet. Sci. 2023, 10, 6. https://doi.org/10.3390/vetsci10010006
Shin Y-K, Kwon O-K, Heo J, Nah J, Kang H-E, Kang Y, Song IJ, Sung H-K. Whole Genome Sequencing of SARS-CoV-2 in Cats and Dogs in South Korea in 2021. Veterinary Sciences. 2023; 10(1):6. https://doi.org/10.3390/vetsci10010006
Chicago/Turabian StyleShin, Yeun-Kyung, Oh-Kyu Kwon, Jinhwa Heo, Jinju Nah, Hae-Eun Kang, Yunhee Kang, In Jun Song, and Ho-Kyung Sung. 2023. "Whole Genome Sequencing of SARS-CoV-2 in Cats and Dogs in South Korea in 2021" Veterinary Sciences 10, no. 1: 6. https://doi.org/10.3390/vetsci10010006
APA StyleShin, Y.-K., Kwon, O.-K., Heo, J., Nah, J., Kang, H.-E., Kang, Y., Song, I. J., & Sung, H.-K. (2023). Whole Genome Sequencing of SARS-CoV-2 in Cats and Dogs in South Korea in 2021. Veterinary Sciences, 10(1), 6. https://doi.org/10.3390/vetsci10010006