An Update to the TraVA Database: Time Series of Capsella bursa-pastoris Shoot Apical Meristems during Transition to Flowering


Abstract
1. Summary
2. Data Description
2.1. Time Series of Capsella Shoot Apical Meristems
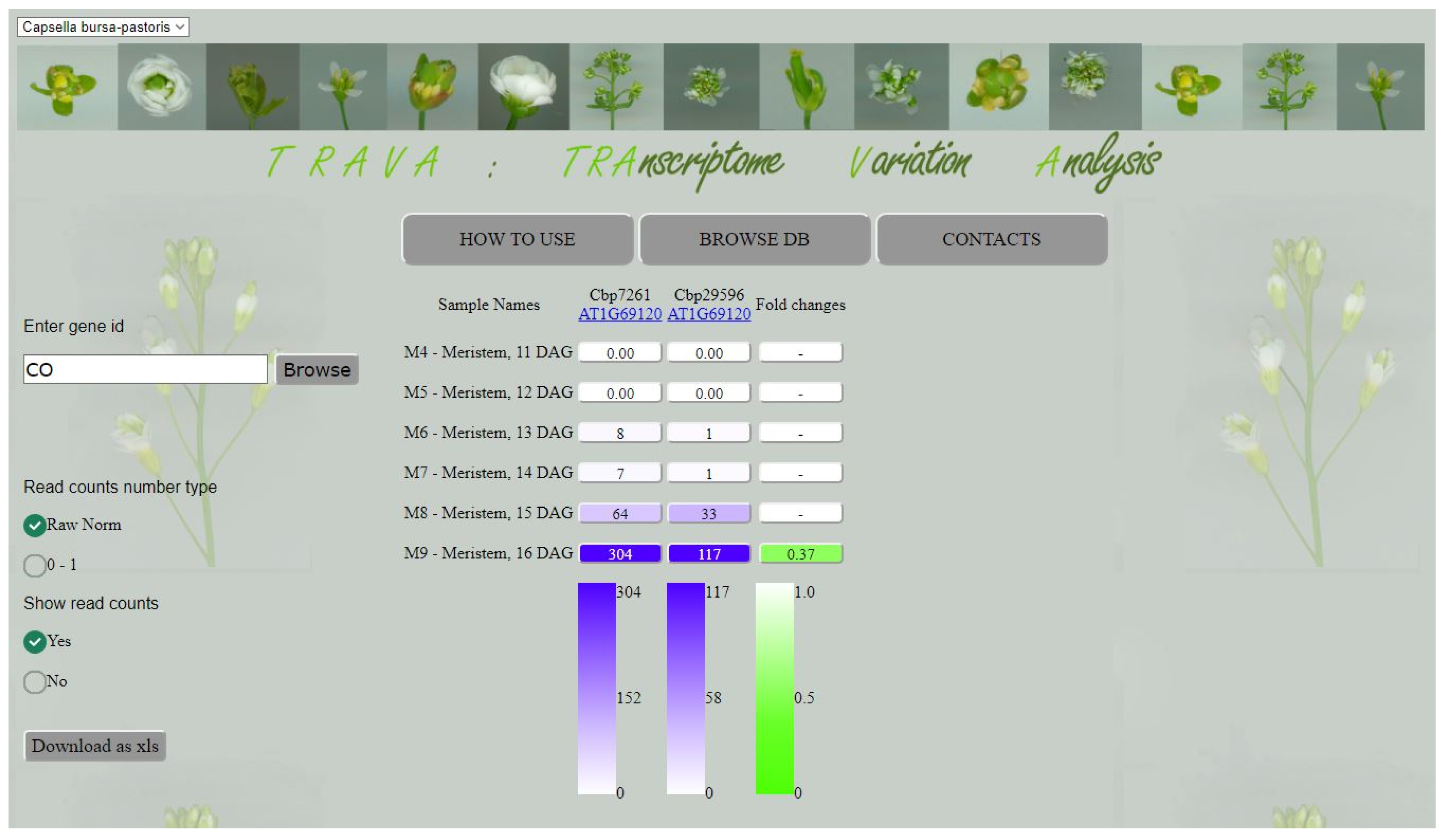
2.2. An Update of Transcriptome Variance Analysis Database
2.3. The Area of Dataset Application
3. Methods
3.1. Plant Growth and Sample Collection
3.2. RNA Extraction and Sequencing
3.3. Data Processing
3.4. Data Availability
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Izawa, T. Adaptation of flowering-time by natural and artificial selection in Arabidopsis and rice. J. Exp. Bot. 2007, 58, 3091–3097. [Google Scholar] [CrossRef] [PubMed]
- Franks, S.J.; Sim, S.; Weis, A.E. Rapid evolution of flowering time by an annual plant in response to a climate fluctuation. Proc. Natl. Acad. Sci. USA 2007, 104, 1278–1282. [Google Scholar] [CrossRef] [PubMed]
- Parra-Tabla, V.; Vargas, C.F. Phenology and Phenotypic Natural Selection on the Flowering Time of a Deceit-pollinated Tropical Orchid, Myrmecophila christinae. Ann. Bot. 2004, 94, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.T.; Lee, C.-R.; Mitchell-Olds, T. Life-history QTLs and natural selection on flowering time in boechera stricta, a perennial relative of arabidopsis: Flowering time in Boechera stricta. Evolution 2011, 65, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Conti, L. Hormonal control of the floral transition: Can one catch them all? Dev. Biol. 2017, 430, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Gaudinier, A.; Blackman, B.K. Evolutionary processes from the perspective of flowering time diversity. New Phytol. 2020, 225, 1883–1898. [Google Scholar] [CrossRef] [PubMed]
- Bar-Joseph, Z.; Gitter, A.; Simon, I. Studying and modelling dynamic biological processes using time-series gene expression data. Nat. Rev. Genet. 2012, 13, 552–564. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.; Fornara, F.; Vincent, C.; Andrés, F.; Nordström, K.; Göbel, U.; Knoll, D.; Schoof, H.; Coupland, G. Analysis of the Arabidopsis shoot meristem transcriptome during floral transition identifies distinct regulatory patterns and a leucine-rich repeat protein that promotes flowering. Plant Cell 2012, 24, 444–462. [Google Scholar] [CrossRef] [PubMed]
- Klepikova, A.V.; Logacheva, M.D.; Dmitriev, S.E.; Penin, A.A. RNA-seq analysis of an apical meristem time series reveals a critical point in Arabidopsis thaliana flower initiation. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Gos, G.; Steige, K.A.; Salcedo, A.; Holm, K.; Josephs, E.B.; Arunkumar, R.; Ågren, J.A.; Hazzouri, K.M.; Wang, W.; et al. Hybrid origins and the earliest stages of diploidization in the highly successful recent polyploid Capsella bursa-pastoris. Proc. Natl. Acad. Sci. USA 2015, 112, 2806–2811. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-R.; Yan, P.-C.; Lascoux, M.; Ge, X.-J. Flowering time and transcriptome variation in Capsella bursa-pastoris (Brassicaceae). New Phytol. 2012, 194, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Toorop, P.E.; Campos Cuerva, R.; Begg, G.S.; Locardi, B.; Squire, G.R.; Iannetta, P.P.M. Co-adaptation of seed dormancy and flowering time in the arable weed Capsella bursa-pastoris (shepherd’s purse). Ann. Bot. 2012, 109, 481–489. [Google Scholar] [CrossRef]
- Kasianov, A.S.; Klepikova, A.V.; Kulakovskiy, I.V.; Gerasimov, E.S.; Fedotova, A.V.; Besedina, E.G.; Kondrashov, A.S.; Logacheva, M.D.; Penin, A.A. High-quality genome assembly of Capsella bursa-pastoris reveals asymmetry of regulatory elements at early stages of Polyploid genome evolution. Plant J. 2017, 91, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Borner, R.; Kampmann, G.; Chandler, J.; Gleissner, R.; Wisman, E.; Apel, K.; Melzer, S. A MADS domain gene involved in the transition to flowering in Arabidopsis. Plant J. 2000, 24, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Oh, M.; Park, H.; Lee, I. SOC1 translocated to the nucleus by interaction with AGL24 directly regulates leafy. Plant J. 2008, 55, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Penin, A.A.; Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D. Comparative Analysis of Developmental Transcriptome Maps of Arabidopsis thaliana and Solanum lycopersicum. Genes 2019, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Kubota, A.; Kwon, M.S.; Covington, M.F.; Lee, N.; Taagen, E.R.; Laboy Cintrón, D.; Hwang, D.Y.; Akiyama, R.; Hodge, S.K.; et al. Molecular basis of flowering under natural long-day conditions in Arabidopsis. Nat. Plants 2018, 4, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample ID | DAG | Rosette Diameter, mm (Avg ± SD, 10 Plants) | Exact Number of Leaf/Floral Primordia | Number of Uniquely Mapped on Homeologs Reads (rep1/rep2) | r2 for Biological Replicates |
|---|---|---|---|---|---|
| M4 | 11 | 39.9 ± 1.4 | 15/0 | 11,280,688/12,029,745 | 0.96 |
| M5 | 12 | 44.8 ± 1.2 | 19/0 | 11,842,692/13,430,904 | 0.90 |
| M6 | 13 | 59.6 ± 2.3 | 20/0 | 13,055,110/15,358,743 | 0.93 |
| M7 | 14 | 78.0 ± 2.3 | 20/0 | 16,918,804/14,085,946 | 0.97 |
| M8 | 15 | 94.7 ± 2.5 | 24/0 | 9,323,196/10,942,328 | 0.97 |
| M9 | 16 | 95.3 ± 2.2 | 24/5 | 9,851,007/10,426,446 | 0.97 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klepikova, A.V.; Kasianov, A.S. An Update to the TraVA Database: Time Series of Capsella bursa-pastoris Shoot Apical Meristems during Transition to Flowering. Data 2020, 5, 58. https://doi.org/10.3390/data5030058
Klepikova AV, Kasianov AS. An Update to the TraVA Database: Time Series of Capsella bursa-pastoris Shoot Apical Meristems during Transition to Flowering. Data. 2020; 5(3):58. https://doi.org/10.3390/data5030058
Chicago/Turabian StyleKlepikova, Anna V., and Artem S. Kasianov. 2020. "An Update to the TraVA Database: Time Series of Capsella bursa-pastoris Shoot Apical Meristems during Transition to Flowering" Data 5, no. 3: 58. https://doi.org/10.3390/data5030058
APA StyleKlepikova, A. V., & Kasianov, A. S. (2020). An Update to the TraVA Database: Time Series of Capsella bursa-pastoris Shoot Apical Meristems during Transition to Flowering. Data, 5(3), 58. https://doi.org/10.3390/data5030058