Red Rooibos Tea Stimulates Osteoblast Mineralization in a Dose-Dependent Manner


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Preparation of Red Rooibos Tea, Determination of Total Polyphenol Content, and Antioxidant Activity
2.3. Cell Culture
2.4. Mineralization
2.5. Cellular Activity Assay
2.6. RT-qPCR
2.7. Immunoblotting
2.8. ALP Activity Assay
2.9. NPP1 Activity Assay
2.10. Statistical Analysis
3. Results
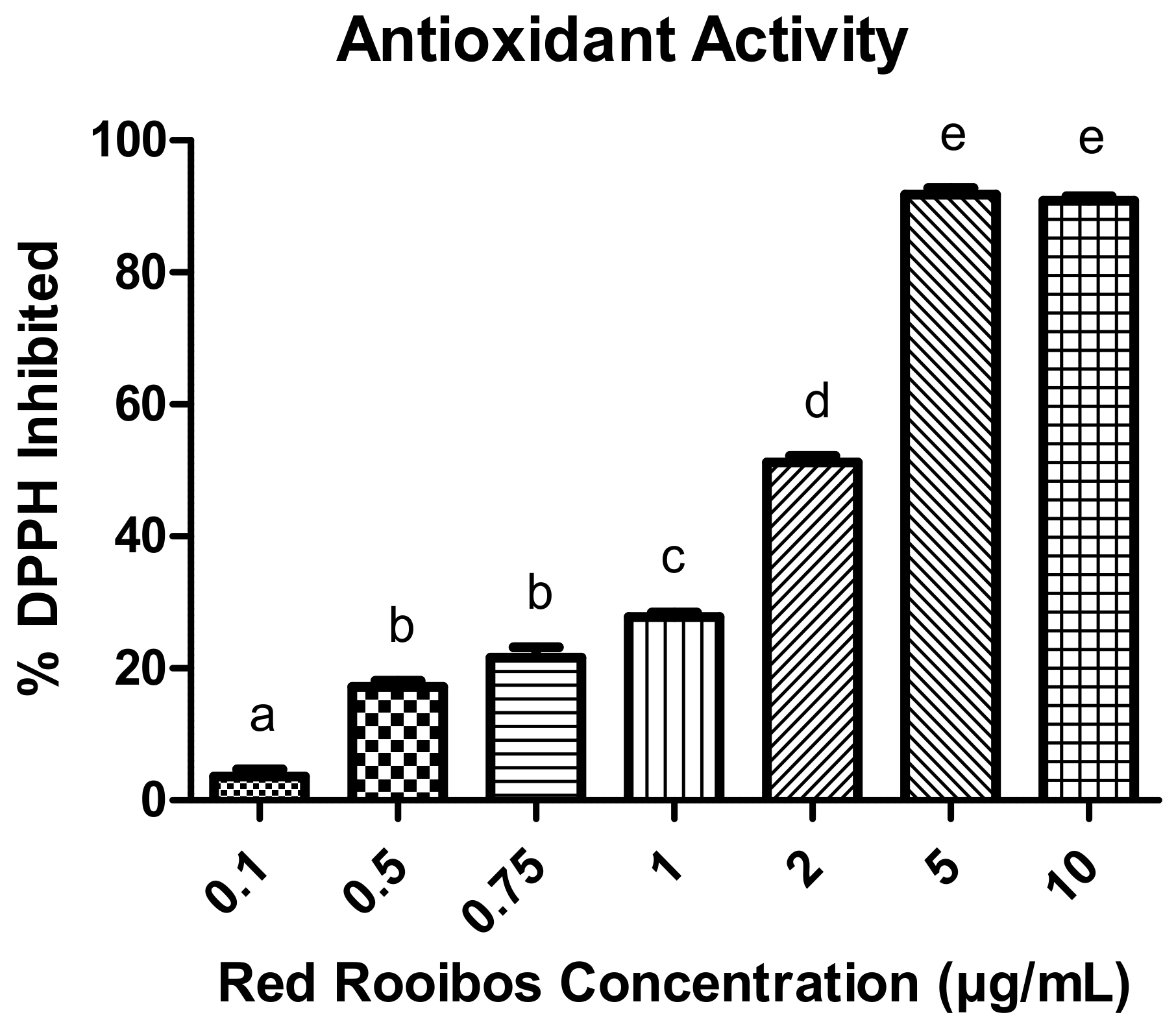
3.1. Antioxidant Activity
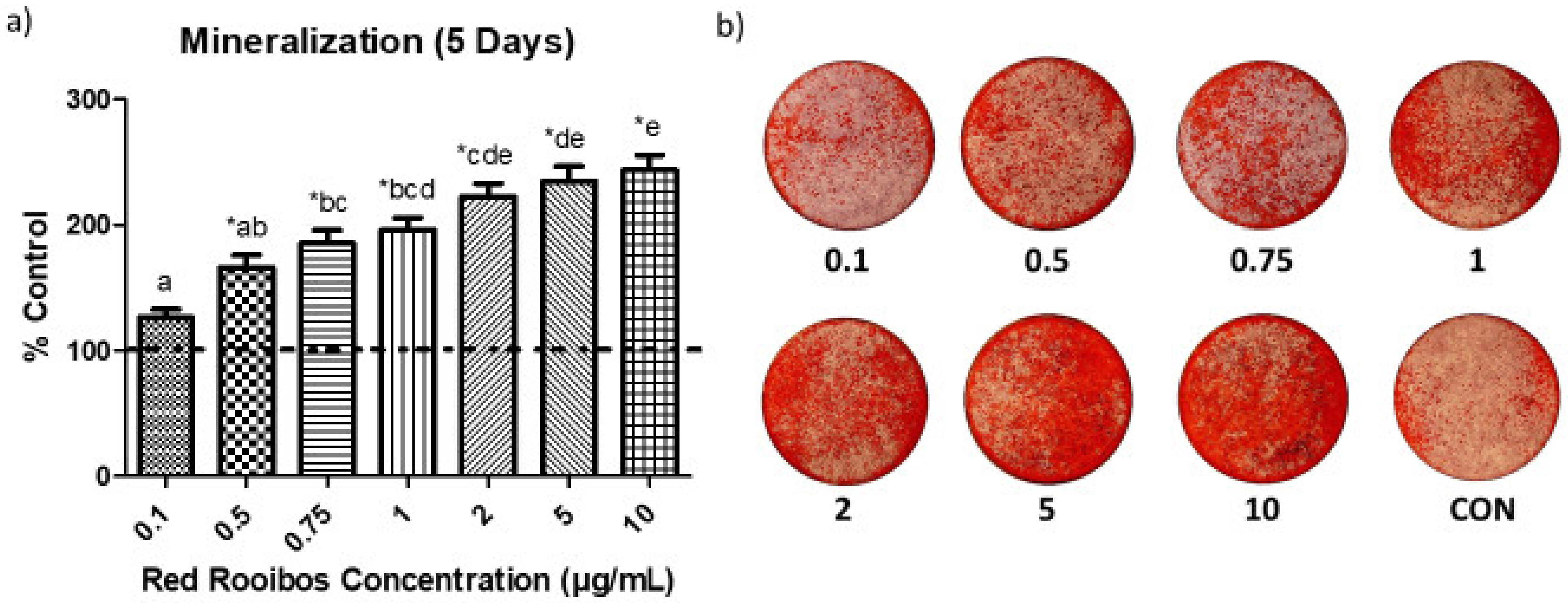
3.2. Mineralization
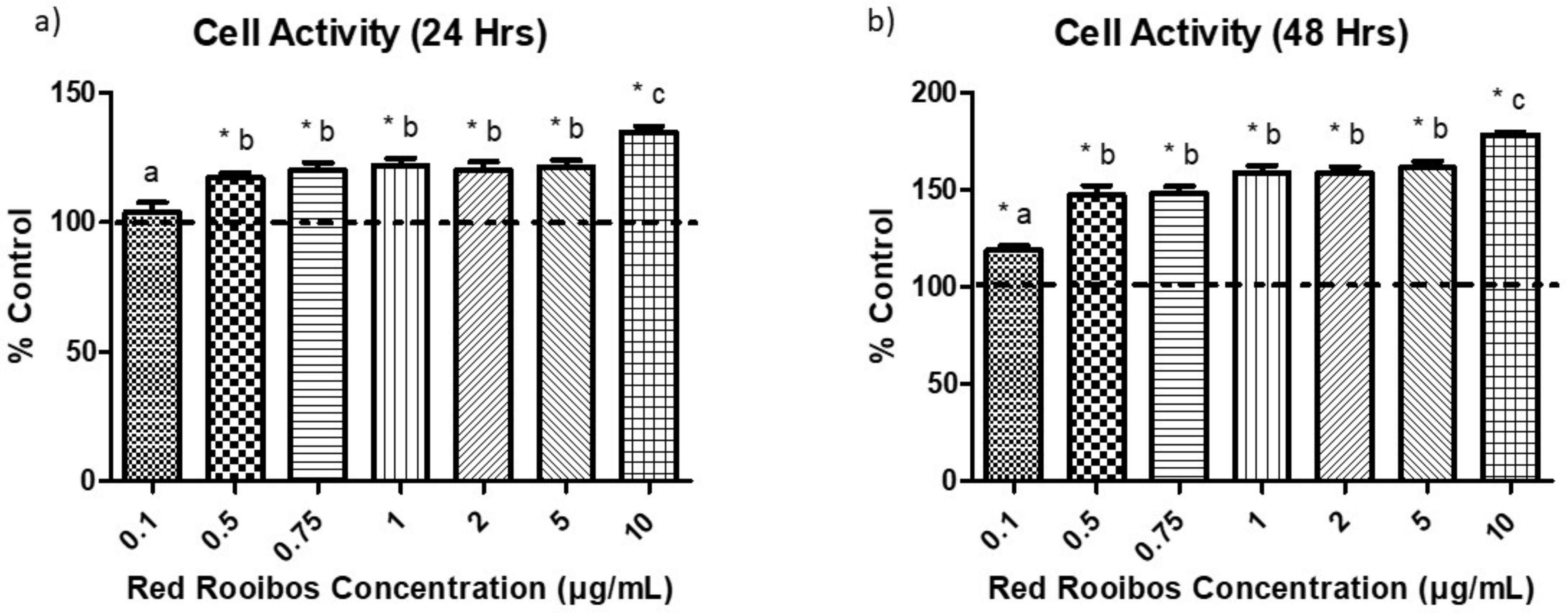
3.3. Cell Activity
3.4. Expression of ALP, NPP1, SOST, and OPN Genes
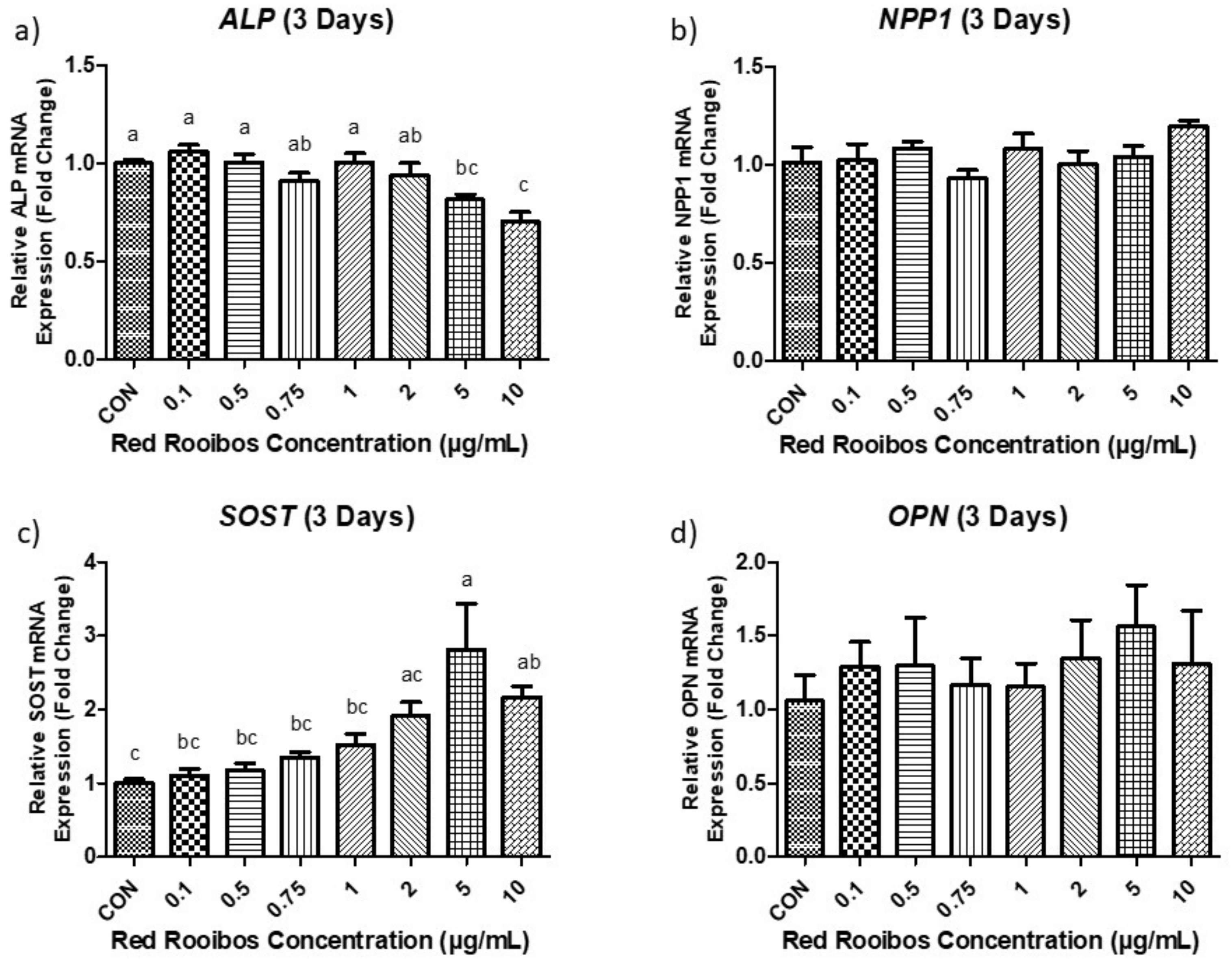
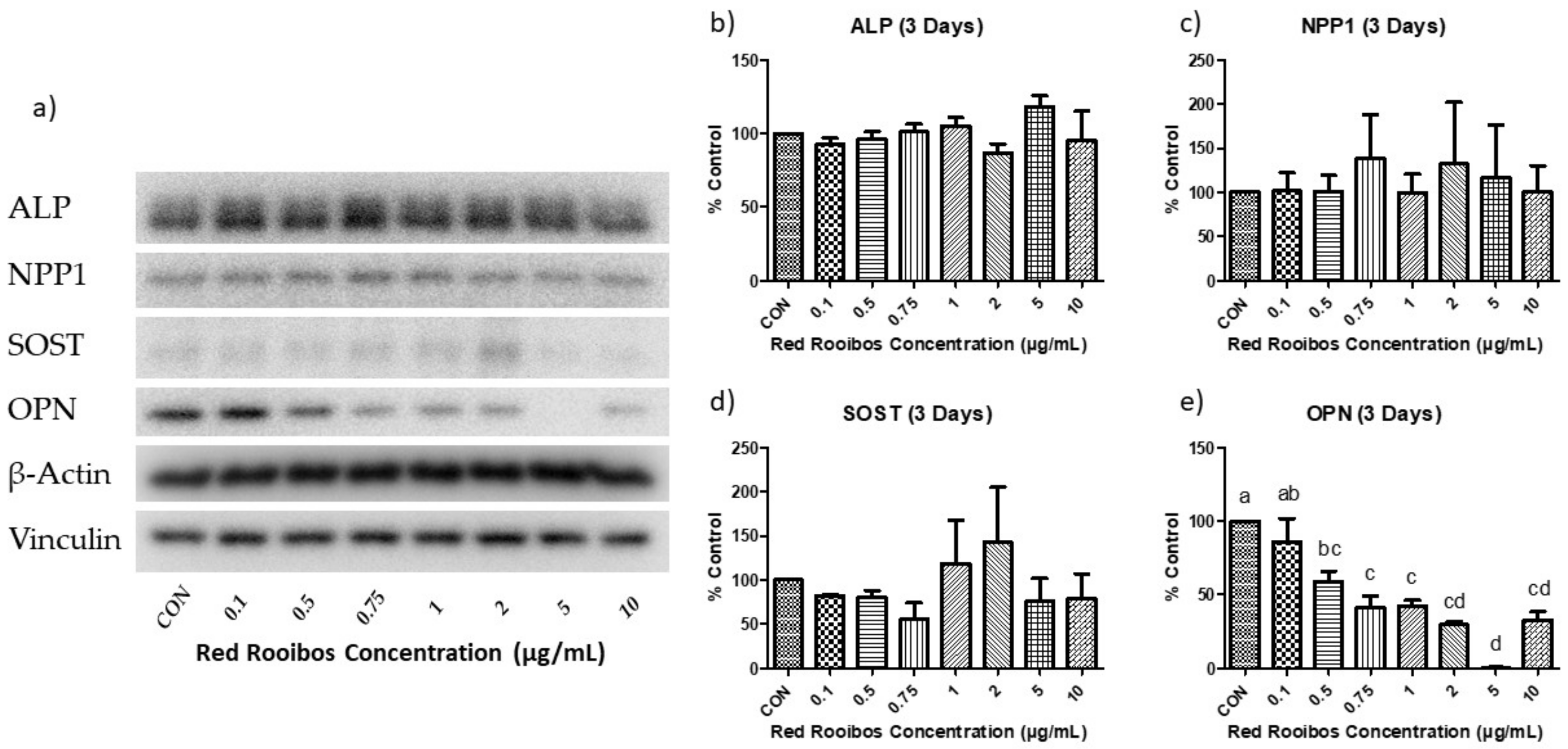
3.4.1. Mineralization Day 3 (Gene Expression)
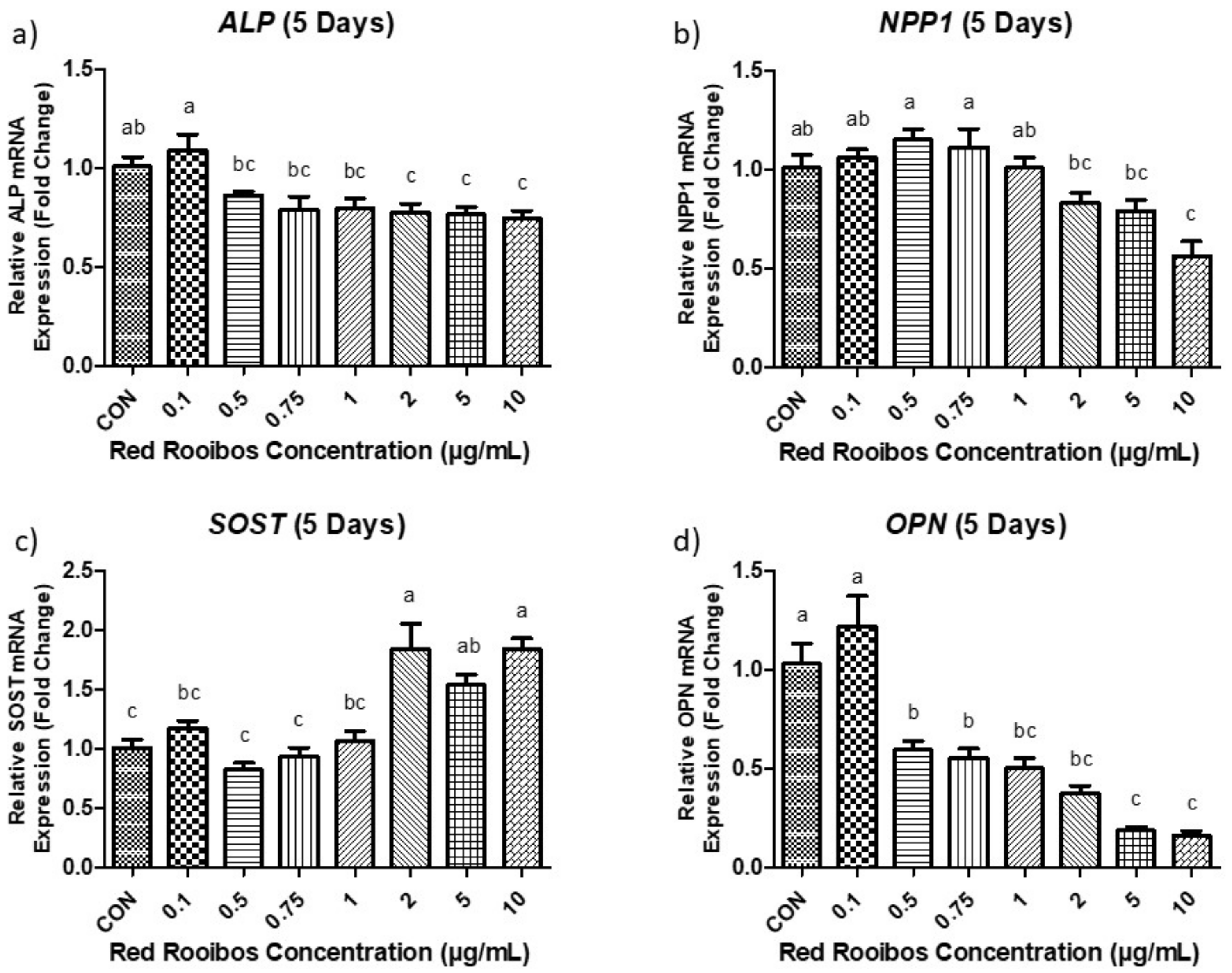
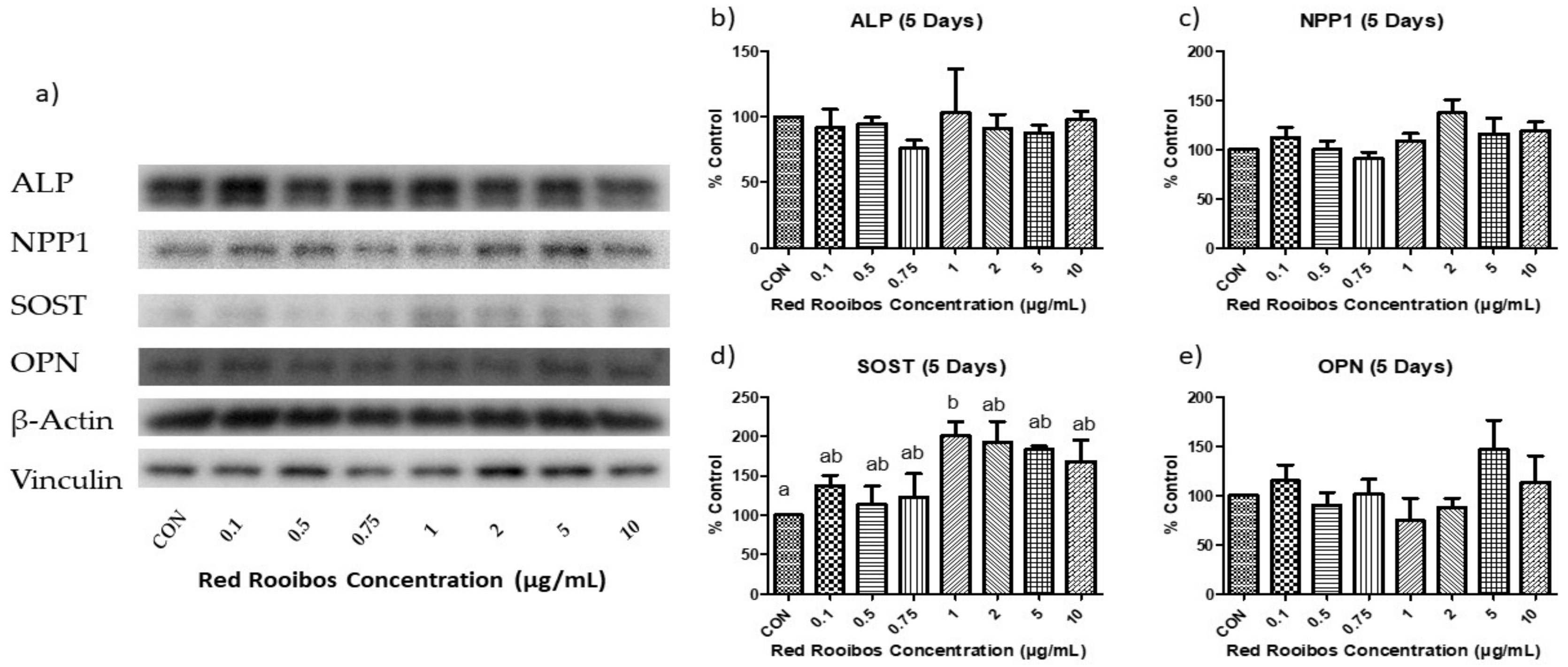
3.4.2. Mineralization Day 5 (Gene Expression)
3.5. Protein Expression of ALP, NPP1, SOST, and OPN
3.5.1. Mineralization Day 3 (Protein Expression)
3.5.2. Mineralization Day 5 (Protein Expression)
3.6. ALP and NPP1 Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chan, G.K.; Duque, G. Age-related bone loss: Old bone, new facts. Gerontology 2002, 48, 62–71. [Google Scholar] [CrossRef]
- Tella, S.H.; Gallagher, J.C. Prevention and treatment of postmenopausal osteoporosis. J. Steroid Biochem. Mol. Biol. 2014, 142, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Cauley, J.A. Public health impact of osteoporosis. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Bliuc, D.; Nguyen, N.D.; Nguyen, T.V.; Eisman, J.A.; Center, J.R. Compound risk of high mortality following osteoporotic fracture and refracture in elderly women and men. J. Bone Miner. Res. 2013, 28, 2317–2324. [Google Scholar] [CrossRef]
- Magaziner, J.; Fredman, L.; Hawkes, W.; Hebel, J.R.; Zimmerman, S.; Orwig, D.L.; Wehren, L. Changes in functional status attributable to hip fracture: A comparison of hip fracture patients to community-dwelling aged. Am. J. Epidemiol. 2003, 157, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Cashman, K. Prebiotics and calcium bioavailability. Curr. Issues Intest. Microbiol. 2003, 4, 21–32. [Google Scholar]
- Cotter, A.; Cashman, K.D. Genistein appears to prevent early postmenopausal bone loss as effectively as hormone replacement therapy. Nutr. Rev. 2003, 61, 346–351. [Google Scholar] [CrossRef]
- Weaver, C.M.; Alekel, D.L.; Ward, W.E.; Ronis, M.J. Flavonoid intake and bone health. J. Nutr. Gerontol. Geriatr. 2012, 31, 239–253. [Google Scholar] [CrossRef]
- New, S.A. Intake of fruit and vegetables: Implications for bone health. Proc. Nutr. Soc. 2003, 62, 889–899. [Google Scholar] [CrossRef]
- Kruger, M.C.; Tousen, Y.; Katsumata, S.; Tadaishi, M.; Kasonga, A.E.; Deepak, V.; Coetzee, M.; Ishimi, Y. Effects of soy phytoestrogens and New Zealand functional foods on bone health. J. Nutr. Sci. Vitaminol. (Tokyo) 2015, 61, S142–S144. [Google Scholar] [CrossRef]
- Nash, L.A.; Ward, W.E. Tea and bone health: Findings from human studies, potential mechanisms, and identification of knowledge gaps. Crit. Rev. Food Sci. Nutr. 2017, 57, 1603–1617. [Google Scholar] [CrossRef]
- Yang, C.S.; Ju, J.; Lu, G.; Xiao, H.; Hao, X.; Sang, S.; Lambert, J.D. Cancer prevention by tea and tea polyphenols. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 245–248. [Google Scholar]
- Deka, A.; Vita, J.A. Tea and cardiovascular disease. Pharmacol. Res. 2011, 64, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Breiter, T.; Laue, C.; Kressel, G.; Groll, S.; Engelhardt, U.H.; Hahn, A. Bioavailability and antioxidant potential of rooibos flavonoids in humans following the consumption of different rooibos formulations. Food Chem. 2011, 128, 338–347. [Google Scholar] [CrossRef]
- McAlpine, M.D.; Ward, W.E. Influence of steep time on polyphenol content and antioxidant capacity of black, green, rooibos, and herbal teas. Beverages 2016, 2, 17. [Google Scholar] [CrossRef]
- Hegarty, V.M.; May, H.M.; Khaw, K.T. Tea drinking and bone mineral density in older women. Am. J. Clin. Nutr. 2000, 71, 1003–1007. [Google Scholar] [CrossRef]
- Wu, C.H.; Yang, Y.C.; Yao, W.J.; Lu, F.H.; Wu, J.S.; Chang, C.J. Epidemiological evidence of increased bone mineral density in habitual tea drinkers. Arch. Intern. Med. 2002, 162, 1001–1006. [Google Scholar] [CrossRef]
- Devine, A.; Hodgson, J.M.; Dick, I.M.; Prince, R.L. Tea drinking is associated with benefits on bone density in older women. Am. J. Clin. Nutr. 2007, 86, 1243–1247. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Yang, J.L.; Jiang, H.C.; Lai, Z.; Wu, F.; Liu, Z.X. Updated association of tea consumption and bone mineral density: A meta-analysis. Medicine (Baltim.) 2017, 96, e6437. [Google Scholar] [CrossRef]
- Sheng, J.; Qu, X.; Zhang, X.; Zhai, Z.; Li, H.; Liu, X.; Li, H.; Liu, G.; Zhu, Z.; Hao, Y.; et al. Coffee, tea, and the risk of hip fracture: A meta-analysis. Osteoporosis Int. 2014, 25, 141–150. [Google Scholar] [CrossRef]
- Chen, B.; Shi, H.F.; Wu, S.C. Tea consumption didn′t modify the risk of fracture: A dose-response meta-analysis of observational studies. Diagn. Pathol. 2014, 9, 44. [Google Scholar] [CrossRef]
- Das, A.S.; Das, D.; Mukherjee, M.; Mukherjee, S.; Mitra, C. Phytoestrogenic effects of black tea extract (Camellia sinensis) in an oophorectomized rat (Rattus norvegicus) model of osteoporosis. Life Sci. 2005, 77, 3049–3057. [Google Scholar] [CrossRef]
- Shen, C.L.; Wang, P.; Guerrieri, J.; Yeh, J.K.; Wang, J.S. Protective effect of green tea polyphenols on bone loss in middle-aged female rats. Osteoporos Int. 2008, 19, 979–990. [Google Scholar] [CrossRef]
- Chen, C.H.; Kang, L.; Lin, R.W.; Fu, Y.C.; Lin, Y.S.; Chang, J.K.; Chen, H.T.; Chen, C.H.; Lin, S.Y.; Wang, G.J.; et al. (-)-Epigallocatechin-3-gallate improves bone microarchitecture in ovariectomized rats. Menopause 2013, 20, 687–694. [Google Scholar] [CrossRef]
- Shen, C.L.; Yeh, J.K.; Stoecker, B.J.; Chyu, M.C.; Wang, J.S. Green tea polyphenols mitigate deterioration of bone microarchitecture in middle-aged female rats. Bone 2009, 44, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Addison, W.N.; Azari, F.; Sorensen, E.S.; Kaartinen, M.T.; McKee, M.D. Pyrophosphate inhibits mineralization of osteoblast cultures by binding to mineral, up-regulating osteopontin, and inhibiting alkaline phosphatase activity. J. Biol. Chem. 2007, 282, 15872–15883. [Google Scholar] [CrossRef] [PubMed]
- Sodek, J.; Ganss, B.; McKee, M.D. Osteopontin. Crit. Rev. Oral. Biol. Med. 2000, 11, 279–303. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. The amazing osteocyte. J. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef]
- Bramati, L.; Minoggio, M.; Gardana, C.; Simonetti, P.; Mauri, P.; Pietta, P. Quantitative characterization of flavonoid compounds in Rooibos tea (Aspalathus linearis) by LC-UV/DAD. J. Agric. Food Chem. 2002, 50, 5513–5519. [Google Scholar] [CrossRef]
- Chen, C.H.; Ho, M.L.; Chang, J.K.; Hung, S.H.; Wang, G.J. Green tea catechin enhances osteogenesis in a bone marrow mesenchymal stem cell line. Osteoporos Int. 2005, 16, 2039–2045. [Google Scholar] [CrossRef]
- Vali, B.; Rao, L.G.; El-Sohemy, A. Epigallocatechin-3-gallate increases the formation of mineralized bone nodules by human osteoblast-like cells. J. Nutr. Biochem. 2007, 18, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Nash, L.A.; Ward, W.E. Comparison of black, green and rooibos tea on osteoblast activity. Food Funct. 2016, 7, 1166–1175. [Google Scholar] [CrossRef]
- Liang, Q.; Lv, M.; Zhang, X.; Hu, J.; Wu, Y.; Huang, Y.; Wang, X.; Sheng, J. Effect of black tea extract and thearubigins on osteoporosis in rats and osteoclast formation in vitro. Front. Physiol. 2018, 9, 1225. [Google Scholar] [CrossRef]
- Oka, Y.; Iwai, S.; Amano, H.; Irie, Y.; Yatomi, K.; Ryu, K.; Yamada, S.; Inagaki, K.; Oguchi, K. Tea polyphenols inhibit rat osteoclast formation and differentiation. J. Pharmacol. Sci. 2012, 118, 55–64. [Google Scholar] [CrossRef]
- Nash, L.A.; Sullivan, P.J.; Peters, S.J.; Ward, W.E. Rooibos flavonoids, orientin and luteolin, stimulate mineralization in human osteoblasts through the Wnt pathway. Mol. Nutr. Food Res. 2015, 59, 443–453. [Google Scholar] [CrossRef]
- Moosa, S.; Kasonga, A.E.; Deepak, V.; Marais, S.; Magoshi, I.B.; Bester, M.J.; Kruger, M.C.; Coetzee, M. Rooibos tea extracts inhibit osteoclast formation and activity through the attenuation of NF-kappaB activity in RAW264.7 murine macrophages. Food Funct. 2018, 9, 3301–3312. [Google Scholar] [CrossRef]
- Kurahashi, N.; Sasazuki, S.; Iwasaki, M.; Inoue, M.; Tsugane, S.; Group, J.S. Green tea consumption and prostate cancer risk in Japanese men: A prospective study. Am. J. Epidemiol. 2008, 167, 71–77. [Google Scholar] [CrossRef]
- Neyestani, T.R.; Shariatzade, N.; Kalayi, A.; Gharavi, A.; Khalaji, N.; Dadkhah, M.; Zowghi, T.; Haidari, H.; Shab-bidar, S. Regular daily intake of black tea improves oxidative stress biomarkers and decreases serum C-reactive protein levels in type 2 diabetic patients. Ann. Nutr. Metab. 2010, 57, 40–49. [Google Scholar] [CrossRef]
- Tan, E.K.; Tan, C.; Fook-Chong, S.M.; Lum, S.Y.; Chai, A.; Chung, H.; Shen, H.; Zhao, Y.; Teoh, M.L.; Yih, Y.; et al. Dose-dependent protective effect of coffee, tea, and smoking in Parkinson’s disease: A study in ethnic Chinese. J. Neurol. Sci. 2003, 216, 163–167. [Google Scholar] [CrossRef]
- Fogh, J.; Fogh, J.M.; Orfeo, T. One hundred and twenty-seven cultured human tumor cell lines producing tumors in nude mice. J. Natl. Cancer Inst. 1977, 59, 221–226. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef]
- Lazcano, O.; Li, C.Y.; Pierre, R.V.; O’Duffy, J.D.; Beissner, R.S.; Abell-Aleff, P.C. Clinical utility of the alizarin red S stain on permanent preparations to detect calcium-containing compounds in synovial fluid. Am. J. Clin. Pathol. 1993, 99, 90–96. [Google Scholar] [CrossRef]
- Tada, H.; Shiho, O.; Kuroshima, K.; Koyama, M.; Tsukamoto, K. An improved colorimetric assay for interleukin 2. J. Immunol. Methods 1986, 93, 157–165. [Google Scholar] [CrossRef]
- MacNeil, A.J.; Jiao, S.C.; McEachern, L.A.; Yang, Y.J.; Dennis, A.; Yu, H.; Xu, Z.; Marshall, J.S.; Lin, T.J. MAPK kinase 3 is a tumor suppressor with reduced copy number in breast cancer. Cancer Res. 2014, 74, 162–172. [Google Scholar] [CrossRef][Green Version]
- Sabokbar, A.; Millett, P.J.; Myer, B.; Rushton, N. A rapid, quantitative assay for measuring alkaline phosphatase activity in osteoblastic cells in vitro. Bone Miner. 1994, 27, 57–67. [Google Scholar] [CrossRef]
- Pochmann, D.; Innocente, A.M.; Buffon, A.; Freitas Sarkis, J.J.; Porciuncula, L.O. Biochemical characterization of an ectonucleotide pyrophosphatase/phosphodiesterase (E-NPP, E.C. 3.1.4.1) from rat cardiac soluble and microsomal fractions. J. Enzyme Inhib. Med. Chem. 2012, 27, 29–36. [Google Scholar] [CrossRef][Green Version]
- Romagnoli, C.; Marcucci, G.; Favilli, F.; Zonefrati, R.; Mavilia, C.; Galli, G.; Tanini, A.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Role of GSH/GSSG redox couple in osteogenic activity and osteoclastogenic markers of human osteoblast-like SaOS-2 cells. FEBS J. 2013, 280, 867–879. [Google Scholar] [CrossRef]
- Arai, M.; Shibata, Y.; Pugdee, K.; Abiko, Y.; Ogata, Y. Effects of reactive oxygen species (ROS) on antioxidant system and osteoblastic differentiation in MC3T3-E1 cells. IUBMB Life 2007, 59, 27–33. [Google Scholar] [CrossRef]
- Ueno, T.; Yamada, M.; Igarashi, Y.; Ogawa, T. N-acetyl cysteine protects osteoblastic function from oxidative stress. J. Biomed. Mater. Res. A 2011, 99, 523–531. [Google Scholar] [CrossRef]
- Almeida, M.; Han, L.; Martin-Millan, M.; Plotkin, L.I.; Stewart, S.A.; Roberson, P.K.; Kousteni, S.; O’Brien, C.A.; Bellido, T.; Parfitt, A.M.; et al. Skeletal involution by age-associated oxidative stress and its acceleration by loss of sex steroids. J. Biol. Chem. 2007, 282, 27285–27297. [Google Scholar] [CrossRef]
- Bai, X.C.; Lu, D.; Bai, J.; Zheng, H.; Ke, Z.Y.; Li, X.M.; Luo, S.Q. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-kappaB. Biochem. Biophys. Res. Commun. 2004, 314, 197–207. [Google Scholar] [CrossRef]
- Yamada, M.; Ueno, T.; Minamikawa, H.; Sato, N.; Iwasa, F.; Hori, N.; Ogawa, T. N-acetyl cysteine alleviates cytotoxicity of bone substitute. J. Dent. Res. 2010, 89, 411–416. [Google Scholar] [CrossRef]
- Jun, J.H.; Lee, S.H.; Kwak, H.B.; Lee, Z.H.; Seo, S.B.; Woo, K.M.; Ryoo, H.M.; Kim, G.S.; Baek, J.H. N-acetylcysteine stimulates osteoblastic differentiation of mouse calvarial cells. J. Cell. Biochem. 2008, 103, 1246–1255. [Google Scholar] [CrossRef]
- Huh, Y.J.; Kim, J.M.; Kim, H.; Song, H.; So, H.; Lee, S.Y.; Kwon, S.B.; Kim, H.J.; Kim, H.H.; Lee, S.H.; et al. Regulation of osteoclast differentiation by the redox-dependent modulation of nuclear import of transcription factors. Cell Death Differ. 2006, 13, 1138–1146. [Google Scholar] [CrossRef]
- Jilka, R.L.; Noble, B.; Weinstein, R.S. Osteocyte apoptosis. Bone 2013, 54, 264–271. [Google Scholar] [CrossRef]
- Shen, C.L.; Chyu, M.C.; Yeh, J.K.; Zhang, Y.; Pence, B.C.; Felton, C.K.; Brismee, J.M.; Arjmandi, B.H.; Doctolero, S.; Wang, J.S. Effect of green tea and Tai Chi on bone health in postmenopausal osteopenic women: A 6-month randomized placebo-controlled trial. Osteoporos Int. 2012, 23, 1541–1552. [Google Scholar] [CrossRef]
- Lyle, A.N.; Joseph, G.; Fan, A.E.; Weiss, D.; Landazuri, N.; Taylor, W.R. Reactive oxygen species regulate osteopontin expression in a murine model of postischemic neovascularization. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1383–1391. [Google Scholar] [CrossRef]
- Prideaux, M.; Wijenayaka, A.R.; Kumarasinghe, D.D.; Ormsby, R.T.; Evdokiou, A.; Findlay, D.M.; Atkins, G.J. SaOS2 Osteosarcoma cells as an in vitro model for studying the transition of human osteoblasts to osteocytes. Calcif. Tissue Int. 2014, 95, 183–193. [Google Scholar] [CrossRef]
- Burgers, T.A.; Williams, B.O. Regulation of Wnt/beta-catenin signaling within and from osteocytes. Bone 2013, 54, 244–249. [Google Scholar] [CrossRef]
- Nash, L.A.; Peters, S.J.; Sullivan, P.J.; Ward, W.E. Supraphysiological levels of quercetin glycosides are required to alter mineralization in Saos2 cells. Int. J. Environ. Res. Public Health 2016, 13, 460. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McAlpine, M.D.; Gittings, W.; MacNeil, A.J.; Ward, W.E. Red Rooibos Tea Stimulates Osteoblast Mineralization in a Dose-Dependent Manner. Beverages 2019, 5, 69. https://doi.org/10.3390/beverages5040069
McAlpine MD, Gittings W, MacNeil AJ, Ward WE. Red Rooibos Tea Stimulates Osteoblast Mineralization in a Dose-Dependent Manner. Beverages. 2019; 5(4):69. https://doi.org/10.3390/beverages5040069
Chicago/Turabian StyleMcAlpine, Michael D., William Gittings, Adam J. MacNeil, and Wendy E. Ward. 2019. "Red Rooibos Tea Stimulates Osteoblast Mineralization in a Dose-Dependent Manner" Beverages 5, no. 4: 69. https://doi.org/10.3390/beverages5040069
APA StyleMcAlpine, M. D., Gittings, W., MacNeil, A. J., & Ward, W. E. (2019). Red Rooibos Tea Stimulates Osteoblast Mineralization in a Dose-Dependent Manner. Beverages, 5(4), 69. https://doi.org/10.3390/beverages5040069