Techno-Functional Role of Exopolysaccharides in Cereal-Based, Yogurt-Like Beverages

Abstract
:1. Introduction
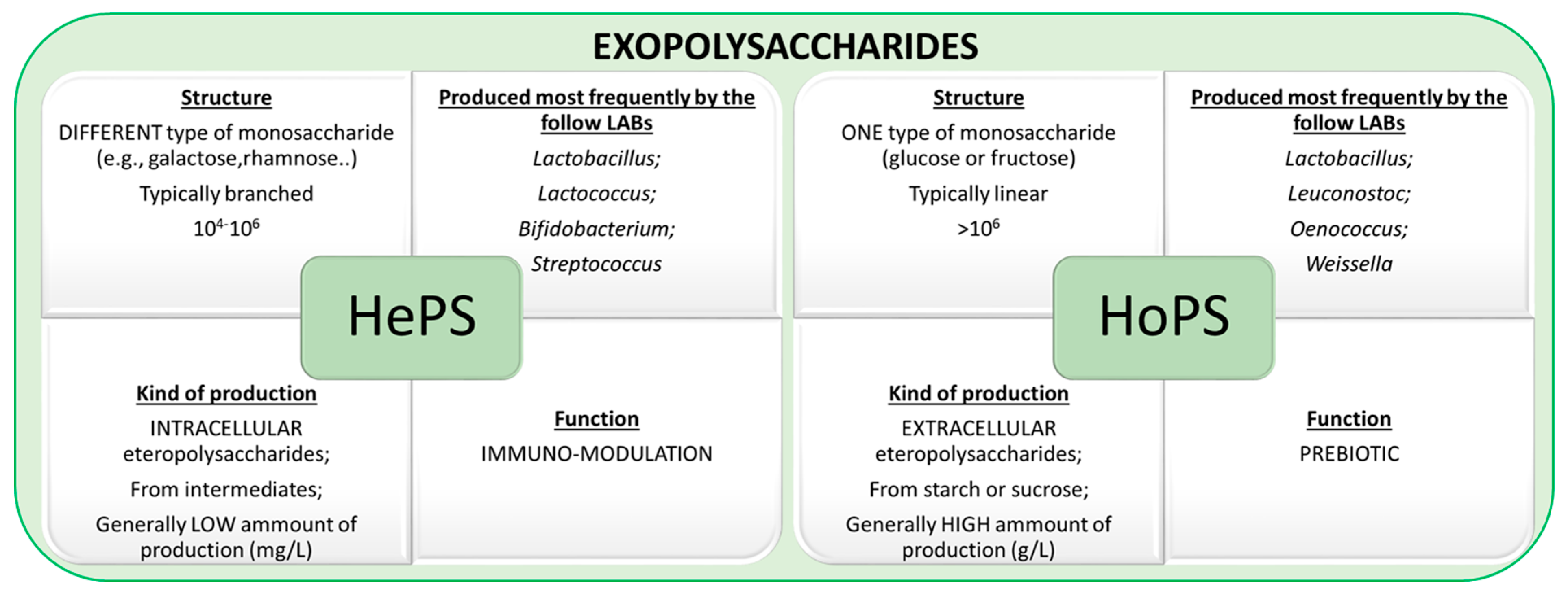
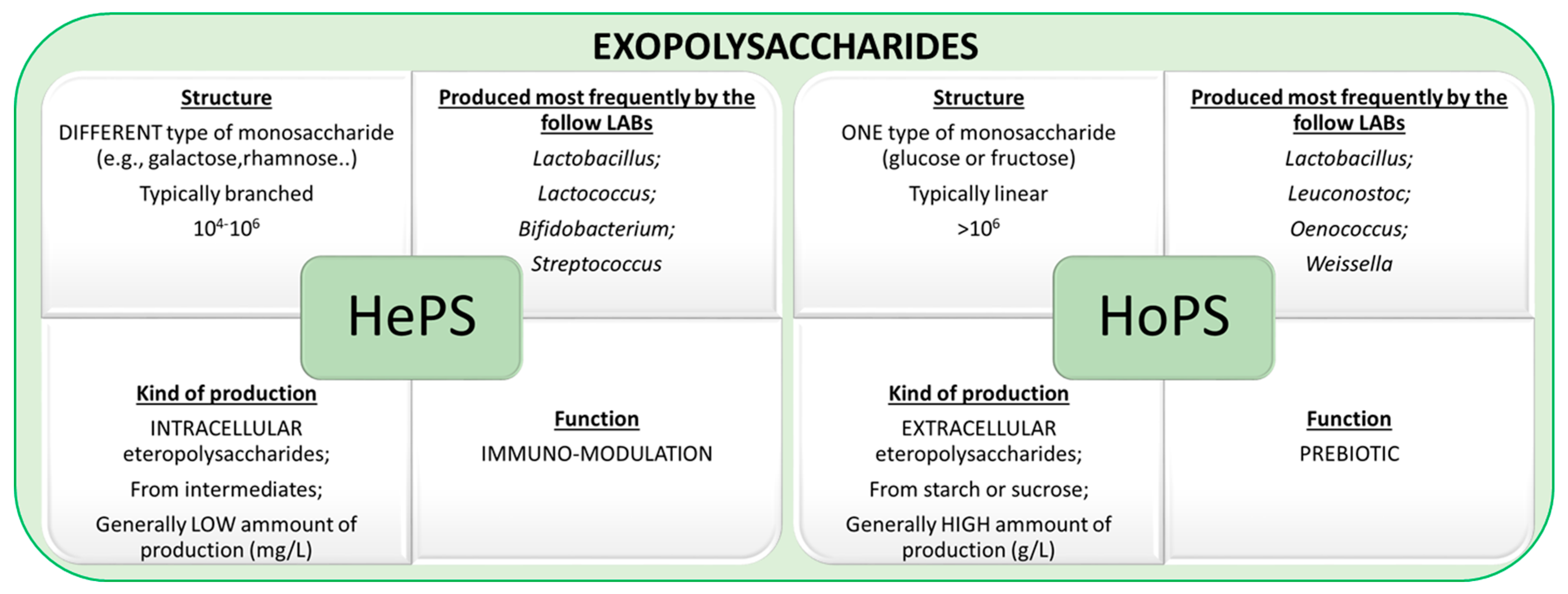
2. Type of Exopolysaccharides
3. Microbial Exopolysaccharides
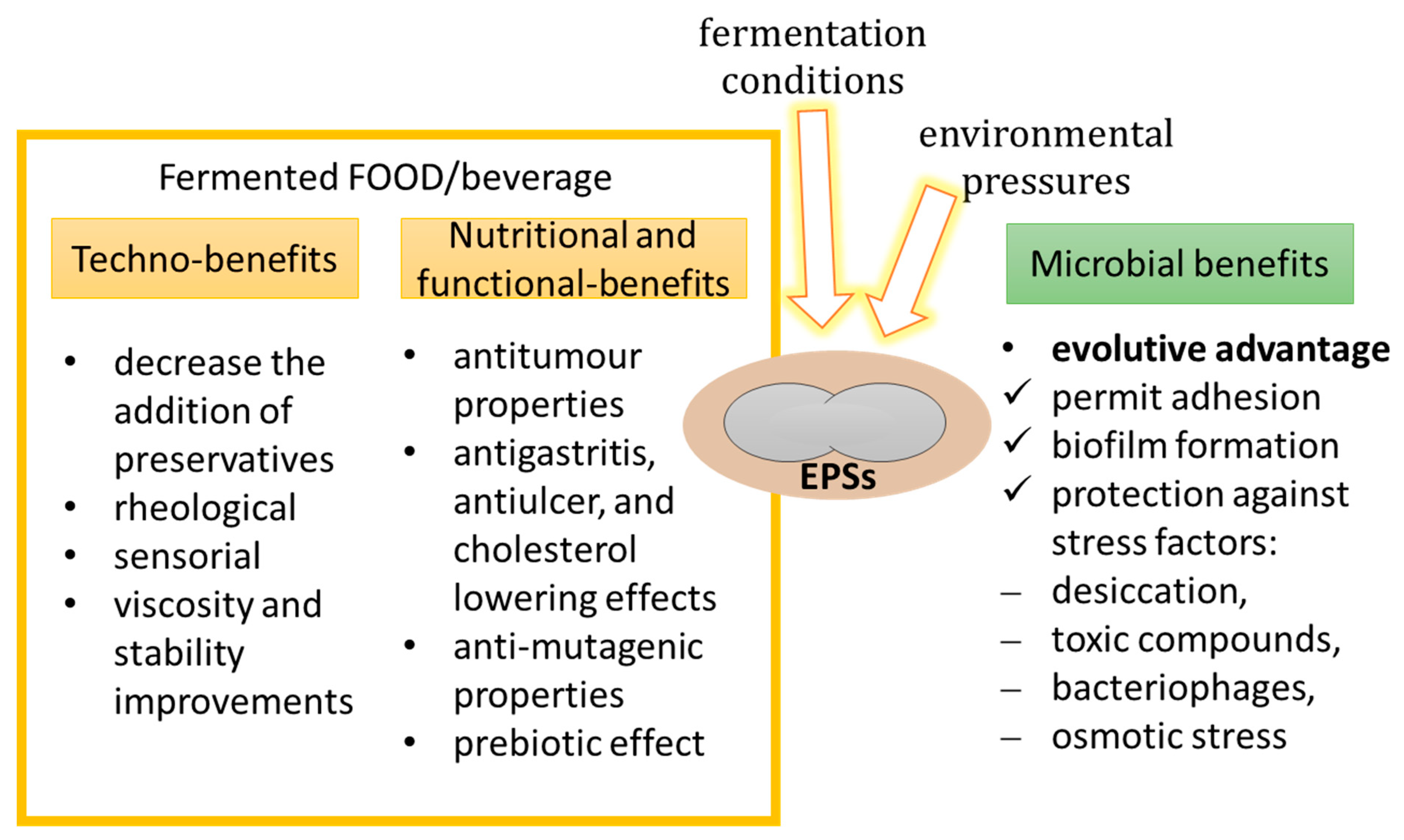
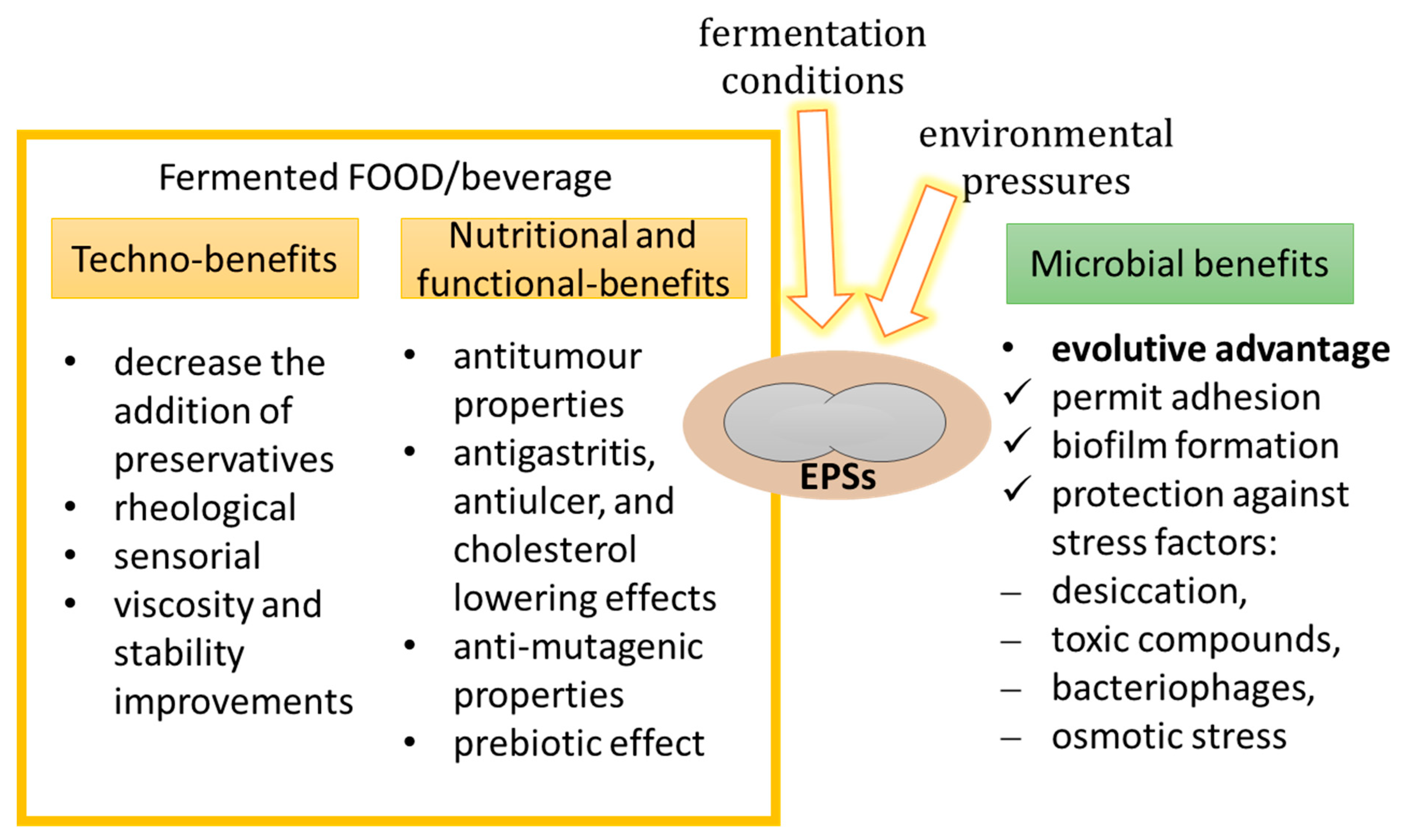
4. EPSs Technical Roles
5. EPSs Functional Roles
6. Cereal-Based Beverages
7. EPS-Producing LABs in Experimental Cereal-Based, Yogurt-Like Beverages
8. Conclusions
Funding
Conflicts of Interest
References
- Coda, R.; Montemurro, M.; Rizzello, C.G. Yogurt-like beverages made with cereals. In Yogurt in Health and Disease Prevention; Shah, N.P., Ed.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: New York, NY, USA, 2017; pp. 183–201. [Google Scholar]
- Greffeuille, V.; Polycarpe Kayode, A.P.; Icard-Vernire, C.; Gnimadi, M.; Rochette, I.; Mouquet-Rivier, C. Changes in iron, zinc and chelating agents during traditional African processing of maize: Effect of iron contamination on bioaccessibility. Food Chem. 2011, 126, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Nionelli, L.; Coda, R.; Curiel, J.A.; Poutanen, K.; Gobbetti, M.; Rizzello, C.G. Manufacture and characterization of a yogurt-like beverage made with oat flakes fermented by selected lactic acid bacteria. Int. J. Food Microbiol. 2014, 185, 17–26. [Google Scholar]
- Hugenholtz, J. Traditional biotechnology for new foods and beverages. Curr. Opin. Biotechnol. 2013, 24, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Deora, N.; Deswal, A. Non-dairy based beverages: An insight. J. Nutr. Food Technol. 2018, 1, 1–4. [Google Scholar] [CrossRef]
- Andhare, P.; Chauhan, K.; Dave, M.; Pathak, H. Microbial Exopolysaccharides: Advances in Applications and Future Prospects. In Biotechnology Volume 3: Microbial Biotechnology; Studium Press LLC: Houston, TX, USA; pp. 1–25. [CrossRef]
- Walling, E.; Gindreau, E.; Lonvaud-Funel, A. A putative glucan synthase gene dps detected in exopolysaccharide-producing Pediococcus damnosus and Oenococcus oeni strains isolated from wine and cider. Int. J. Food Microbiol. 2005, 98, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Garai-Ibabe, G.; Duenas, M.T.; Irastorza, A.; Sierra-Filardi, E.; Werning, M.L.; Lopez, P.; de Palencia, P.F. Naturally occurring 2-substituted (1,3)-β-d-glucan producing Lactobacillus suebicus and Pediococcus parvulus strains with potential utility in the production of functional foods. Bioresour. Technol. 2010, 101, 9254–9263. [Google Scholar] [CrossRef] [PubMed]
- Ripari, V.; Bai, Y.; Gänzle, M.G. Metabolism of phenolic acids in whole wheat and rye malt sourdoughs. Food Microbiol. 2019, 77, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Sanalibaba, P.; Çakmak, G.A. Exopolysaccharides Production by Lactic Acid Bacteria. Appl. Microbiol. Open Access 2016, 2, 1–5. [Google Scholar] [CrossRef]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic Acid Bacteria Exopolysaccharides in Foods and Beverages: Isolation, Properties, Characterization, and Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; De Vin, F.; Vaningelgem, F.; Degeest, B. Recent developments in the biosynthesis and applications of heteropolysaccharides from lactic acid bacteria. Int. Dairy J. 2001, 11, 687–707. [Google Scholar] [CrossRef]
- Kralj, S.; van Geel-Schutten, G.H.; Dondorff, M.M.G.; Kirsanovs, S.; van der Maarel, M.J.E.C.; Dijkhuizen, L. Glucan synthesis in the genus Lactobacillus: Isolation and characterization of glucansucrase genes, enzymes and glucan products from six different strains. Microbiology 2004, 150, 3681–3690. [Google Scholar] [CrossRef] [PubMed]
- Van Hijum, S.A.F.T.; Szalowska, E.; van der Maarel, M.J.E.C.; Dijkhuizen, L. Biochemical and molecular characterization of a levansucrase from Lactobacillus reuteri. Microbiology 2004, 150, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Dave, S.R.; Vaishnav, A.M.; Upadhyay, K.H.; Tipre, D.R. Microbial Exopolysaccharide—An Inevitable Product for Living Beings and Environment. J. Bacteriol. Mycol. Open Access 2016, 2, 00034. [Google Scholar] [CrossRef]
- Poli, A.; Di Donato, P.; Abbamondi, G.R.; Nicolaus, B. Synthesis, Production, and Biotechnological Applications of Exopolysaccharides and Polyhydroxyalkanoates by Archaea. Archaea 2011, 2011, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial exopolysaccharides from extreme marine habitats: Production, characterization and biological activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.A.; Anandapandian, K.T.K.; Parthiban, K. Production and characterization of exopolysaccharides (EPS) from Biofilm forming marine bacterium. Braz. Arch. Biol. Technol. 2011, 54, 259–265. [Google Scholar] [CrossRef]
- Surayot, U.; Wang, J.; Seesuriyachan, P.; Kuntiya, A.; Tabarsa, M.; Lee, Y.; Kim, J.K.; Park, W.; You, S. Exopolysaccharides from lactic acid bacteria: Structural analysis, molecular weight effect on immunomodulation. Int. J. Biol. Macromol. 2014, 68, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Czaczyk, K.; Myszka, K. Biosynthesis of extracellular polymeric substances (EPS) and its role in microbial biofilm formation. Pol. J. Environ. Stud. 2007, 16, 799–806. [Google Scholar]
- Ruas-Madiedo, P.; Medrano, M.; Salazar, N.; de los Losreyes-Gavilán, C.G.; Perez, P.F.; Abraham, A.G. Exopolysaccharides produced by Lactobacillus and Bifidobacterium strains abrogate in vitro the cytotoxic effect of bacterial toxins on eukaryotic cells. J. Appl. Microbiol. 2010, 109, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- Galle, S.; Arendt, E.K. Exopolysaccharides from sourdough lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2014, 54, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Kajala, I.; Mäkelä, J.; Coda, R.; Shukla, S.; Shi, Q.; Maina, N.H.; Juvonen, R.; Ekholm, P.; Goyal, A.; Tenkanen, M.; et al. Rye bran as fermentation matrix boosts in situ dextran production by Weissella confuse compared to wheat bran. Appl. Microbiol. Biotechnol. 2016, 100, 3499–3510. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.R.; Koijam, K. Exopolysaccharide Production by a Lactic Acid Bacteria, Leuconostoc lactis Isolated from Ethnically Fermented Beverage. Natl. Acad. Sci. Lett. 2014, 37, 59–64. [Google Scholar] [CrossRef]
- Tieking, M.; Gänzle, M.G. Exopolysaccharides from cereal-associated lactobacilli. Trends Food Sci. Technol. 2005, 16, 79–84. [Google Scholar] [CrossRef]
- Francieli, F.B.; Ripari, V.; Waszczynskyj, N.; Spier, M.R. Overview of Sourdough Technology: From Production to Marketing. Food Bioprocess Technol. 2018, 11, 242–270. [Google Scholar]
- Lynch, K.M.; Coffey, A.; Arendt, E.K. Exopolysaccharide producing lactic acid bacteria: Their techno-functional role and potential application in gluten-free bread products. Food Res. Int. 2018, 110, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Chien, M.-F.C. Exopolysaccharides production as affected by lactic acid bacteria and fermentation time. Food Chem. 2007, 100, 1419–1423. [Google Scholar] [CrossRef]
- Han, X.; Yang, Z.; Jing, X.; Yu, P.; Zhang, Y.; Yi, H.; Zhang, L. Improvement of the Texture of Yogurt by Use of Exopolysaccharide Producing Lactic Acid Bacteria. BioMed Res. Int. 2016, 2016, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Petel, A.; Prajapati, J.P. Food and Health Applications of Exopolysaccharides produced by Lactic acid Bacteria. Adv. Dairy Res. 2013, 1, 1–7. [Google Scholar]
- Saint-Eve, A.; Martin, N.; Guillemin, H.; Sémon, E.; Guichard, E.; Souchon, I. Flavored yogurt complex viscosity influences real-time aroma release in the mouth and sensory properties. J. Agric. Food Chem. 2006, 54, 7794–7803. [Google Scholar] [CrossRef] [PubMed]
- Juvonen, R.; Honkapää, K.; Maina, N.H.; Shi, Q.; Viljanen, K.; Maaheimo, H.; Virkki, L.; Tenkanen, M.; Lantto, R. The impact of fermentation with exopolysaccharide producing lactic acid bacteria on rheological, chemical and sensory properties of pureed carrots (Daucus carota L.). Int. J. Food Microbiol. 2015, 207, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.A.; Phillips, G.O. Introduction to food hydrocolloids. In Handbook of Hydrocolloids, 2nd ed.; Phillips, G.O., Williams, P.A., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2009; pp. 1–22. [Google Scholar]
- Maina, N.H.; Virkki, L.; Pynnönen, H.; Maaheimo, H.; Tenkanen, M. Structural analysis of enzyme-resistant iso malto oligosaccharides reveals the elongation of α-(1→3)-linked branches in Weissella confusa dextran. Biomacromolecules 2011, 12, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Salazar, N.; Gueimonde, M.; Hernandez-Barranco, A.M.; Ruas-Madiedo, P.; de los Reyes-Gavilan, C.G. Exopolysaccharides produced by intestinal Bifidobacterium strains act as fermentable substrates for human intestinal bacteria. Appl. Environ. Microbiol. 2008, 74, 4737–4745. [Google Scholar] [CrossRef] [PubMed]
- Oda, M.; Hasegawa, H.; Komatsu, S.; Kambe, M.; Tsuchiya, F. Anti-tumor polysaccharide from LactoBacillus spp. Agric. Biol. Chem. 1983, 47, 1623–1625. [Google Scholar] [CrossRef]
- Wang, K.; Li, W.; Rui, X.; Chen, X.; Jiang, M.; Dong, M. Characterization of a novel exopolysaccharide with antitumor activity from Lactobacillus plantarum 70810. Int. J. Biol. Macromol. 2014, 63, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, M.; Hashimoto, S.; Watanabe, T.; Yokokura, T.; Moro, Y. Anti-ulcer effects of lactic acid bacteria and their cell wall polysaccharides. Biol. Pharm. Bull. 1994, 17, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Su, C.A.; Xu, X.Y.; Liu, D.Y.; Wu, M.; Zeng, F.Q.; Zeng, M.Y.; Jiang, N.; Luo, X. Isolation and characterization of exopolysaccharide with immunomodulatory activity from fermentation broth of Morchella conica. J. Pharm. Sci. 2013, 21, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Hirota, T.; Toba, T.; Itoh, T.; Adachi, S. Structure of the extracellular polysaccharide from slime-forming Lactococcus Lactis subsp cremoris SBT 0495. Carbohydr. Res. 1992, 224, 245–253. [Google Scholar] [CrossRef]
- Tok, E.; Aslim, B. Cholesterol removal by some lactic acid bacteria that can be used as probiotic. Microbiol. Immunol. 2010, 54, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Welman, A.D. Exploitation of exopolysaccharides from lactic acid bacteria: Nutritional and functional benefits. In Bacterial Polysaccharides: Current Innovations and Future Trends; Caister Academic Press: Poole, UK, 2009; pp. 331–343. [Google Scholar]
- Rodríguez, C.; Medici, M.; Rodríguez, A.V.; Mozzi, F.; Font de Valdez, G. Prevention of chronic gastritis by fermented milks made with exopolysaccharide producing Streptococcus thermophilus strains. J. Dairy Sci. 2008, 92, 2423–2434. [Google Scholar] [CrossRef] [PubMed]
- Korakli, M.; Gänzle, M.G.; Vogel, R.F. Metabolism by Bifidobacteria and lactic acid bacteria of polysaccharides from wheat and rye, and exopolysaccharides produced by Lactobacillus sanfranciscensis. J. Appl. Microbiol. 2002, 92, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.M.; Ross, R.P.; Fitzgerald, G.F.; Caplice, N.M.; Stanton, C. Sugar-coated: Exopolysaccharide producing lactic acid bacteria for food and human health applications. Food Funct. 2015, 6, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Olano-Martin, E.; Mountzouris, K.C.; Gibson, G.R.; Rastall, R.A. In vitro fermentability of dextran, oligodextran and maltodextrin by human gut bacteria. Br. J. Nutr. 2000, 83, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E.; Sarbu, I.; Nedelcu, I.; Pelinescu, D. Study of PROEXO product influence on infant microbiota in an vitro colonic fermentation system. Ann. Microbiol. 2015, 65, 1189–1193. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory effects triggered by lactic acid bacteria exopolysaccharides: New insights into molecular interactions with host cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, H.; Harata, T.; Uemura, J.; Saito, T.; Kaneko, T.; Itoh, T. Phosphate group requirement for mitogenic activation of lymphocytes by an extracellular phosphopolysaccharide from Lactobacillus delbrueckii ssp. bulgaricus. Int. J. Food Microbiol. 1998, 40, 169–175. [Google Scholar] [CrossRef]
- Tsuda, H.; Hara, K.; Miyamoto, T. Binding of mutagens to exopolysaccharide produced by Lactobacillus plantarum mutant strain 301102S. J. Dairy Sci. 2008, 91, 2960–2966. [Google Scholar] [CrossRef] [PubMed]
- Chabot, S.; Yu, H.L.; De Leseleuc, L.; Cloutier, D.; van Calsteren, M.R.; Lessard, M.; Roy, D.; Lacroix, M.; Oth, D. Exopolysaccharides from Lactobacillus rhamnosus RW-9595M stimulate TNF, IL-6 and IL-12 in human and mouse cultured immunocompetent cells, and IFN- gamma in mouse splenocytes. Lait 2001, 81, 683–697. [Google Scholar] [CrossRef]
- Looijesteijn, P.J.; Trapet, L.; de Vries, E.; Abee, T.; Hugenholtz, J. Physiological function of exopolysaccharides produced by Lactococcus lactis. Int. J. Food Microbiol. 2001, 64, 71–80. [Google Scholar] [CrossRef]
- Yamamoto, S.; Nakashima, Y.; Yoshikawa, J.; Wada, N.; Matsugo, S. Radical scavenging activity of the Japanese traditional food, Amazake. Food Sci. Technol. Res. 2011, 17, 209–218. [Google Scholar] [CrossRef]
- Tamang, J.P.; Thapa, S. Fermentation dynamics during production of bhaati jaanr, a traditional fermented rice beverage of the eastern Himalayas. Food Biotechol. 2006, 20, 251–261. [Google Scholar] [CrossRef]
- Heperkan, D.; Daskaya-Dikmen, C.; Bayram, B. Evaluation of lactic acid bacterial strains of boza for their exopolysaccharide and enzyme production as a potential adjunct culture. Process Biochem. 2014, 49, 1587–1594. [Google Scholar] [CrossRef]
- Muyanja, C.M.B.K.; Narvhus, J.A.; Treimo, J.; Langsrud, T. Isolation, characterization and identification of lactic acid bacteria from bushera: A Ugandan traditional fermented beverage. Int. J. Food Microbiol. 2003, 80, 201–210. [Google Scholar] [CrossRef]
- Thakur, N.; Savitri-Saris, P.E.J.; Bhalla, T.C. Microorganisms associated with amylolytic starters and traditional fermented alcoholic beverages of North Western Himalayas in India. Food Biosci. 2015, 11, 92–96. [Google Scholar] [CrossRef]
- Jargin, S.V. Kvass: A possible contributor to chronic alcoholism in the Former Soviet Union e alcohol content should be indicated on labels and in advertising. Alcohol Alcohol. 2009, 44, 529. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Maity, C.; Adak, A.; Halder, S.K.; Jana, A.; Das, A.; Parua, S.; Mohapatra, P.K.D.; Pati, B.R.; Mondal, K.C. Ethnic preparation of Haria, a rice-based fermented beverage, in the province of Lateritic West Bengal, India. Ethnobot. Res. Appl. 2014, 12, 39–49. [Google Scholar]
- Zuauya, R.; Mygochi, T.; Parawira, W. Microbial and biochemical changes occurring during production of masvusvu and mangisi, traditional Zimbabwean beverages. Plant Food Hum. Nutr. 1997, 51, 43–51. [Google Scholar]
- Das, D.; Goyal, A. Characterization and biocompatibility of glucan: A safe food additive from probiotic Lactobacillus plantarum DM5. J. Sci. Food Agric. 2014, 94, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Sawadogo-Lingani, H.; Diawara, B.; Traore, S.; Jakobsen, M. Technological properties of Lactobacillus fermentum involved in the processing of dolo and pito, West African sorghum beers, for the selection of starter cultures. J. Appl. Microbiol. 2008, 104, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Ji Oh, Y.; Jung, D.S. Evaluation of probiotic properties of Lactobacillus and Pediococcus strains isolated from Omegisool, a traditionally fermented millet alcoholic beverage in Korea. Food Sci. Technol. 2015, 63, 437–444. [Google Scholar]
- Mugula, J.K.; Ninko, S.A.M.; Narvhus, J.A.; Sorhaug, T. Microbiological and fermentation characteristics of togwa, a Tanzanian fermented food. Int. J. Food Microbiol. 2003, 80, 187–199. [Google Scholar] [CrossRef]
- Ray, S.; Bagyaraj, D.J.; Thilagar, G.; Tamang, J.P. Preparation of Chyang, an ethnic fermented beverage of the Himalayas, using different raw cereals. J. Ethn. Foods 2016, 3, 297–299. [Google Scholar] [CrossRef]
- Ripari, V.; Gänzle, M.G.; Berardi, E. Evolution of sourdough microbiota in spontaneous sourdoughs started with different plant materials. Int. J. Food Microbiol. 2016, 232, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Ludena Urquizo, F.E.; Torres, S.M.C.; Tolonen, T.; Jaakkola, M.; Pena-Niebuhr, M.G.; von Wright, A.; Repo-Carrasco-Valencia, R.; Korhonen, H.; Plumed-Ferrer, C. Development of a fermented quinoa-based beverage. Food Sci. Nutr. 2017, 5, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Pelikánová, J.; Liptáková, D.; Valík, L. Suitability of lactic acid bacteria for fermentation of maize and amaranth. J. Food Nutr. Res. 2015, 54, 354–364. [Google Scholar]
- Adebayo-tayo, B.C.; Onilude, A.A. Screening of lactic acid bacteria strains isolated from some Nigerian fermented foods for EPS production. World Appl. Sci. J. 2008, 4, 741–747. [Google Scholar]
- Grobben, G.J.; Van Casteren, W.H.M.; Schols, H.A.; Oosterveld, A.; Sala, G.; Smith, M.R. Analysis of the exopolysaccharides produced by Lactobacillus delbrueckii subsp. bulgaricus NCFB 2772 grown in continuous culture on glucose and fructose. Appl. Microbiol. Biotechnol. 1997, 48, 516–521. [Google Scholar] [CrossRef]
- Magala, M.; Kohajdová, Z.; Karovičová, J.; Greifová, M.; Hojerová, J. Application of Lactic Acid Bacteria for Production of Fermented Beverages Based on Rice Flour. Czech J. Food Sci. 2015, 33, 458–463. [Google Scholar] [CrossRef]
- Russo, P.; de Chiara, M.L.V.; Capozzi, V.; Arena, M.P.; Amodio, M.L.; Rascòn, A.; Dueñas, M.T.; Lòpez, P.; Spano, G. Lactobacillus plantarum strains for multifunctional oat-based foods. Food Sci. Technol. 2016, 68, 288–294. [Google Scholar] [CrossRef]
- Mårtensson, O.; Öste, R.; Holst, O. Lactic acid bacteria in an oat based non-dairy milk substitute: Fermentation characteristics and exopolysaccharide formation. Lebensm. Wiss. Technol. 2000, 33, 525–530. [Google Scholar] [CrossRef]
- Mårtensson, O.; Öste, R.; Holst, O. The effect of yoghurt culture on the survival of probiotic bacteria in oat-based, non-dairy products. Food Res. Int. 2002, 35, 775–784. [Google Scholar] [CrossRef]
- Mårtensson, O.; Dueñas-Chasco, M.; Irastorza, A.; Öste, R.; Holst, O. Comparison of growth characteristics and exopolysaccharide formation of two lactic acid bacteria strains, Pediococcus damnosus 2.6 and Lactobacillus brevis G-77, in an oat-based, nondairy medium. Lebensm. Wiss. Technol. 2003, 36, 353–357. [Google Scholar] [CrossRef]
- Lorusso, A.; Coda, R.; Montemurro, M.; Rizzello, C.G. Use of Selected Lactic Acid Bacteria and Quinoa Flour for Manufacturing Novel Yogurt-Like Beverages. Foods 2018, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Rizzello, C.G.; Trani, A.; Gobbetti, M. Manufacture and characterization of functional emmer beverages fermented by selected lactic acid bacteria. Food Microbiol. 2011, 28, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Lanera, A.; Trani, A.; Gobbetti, M.; Di Cagno, R. Yogurt-like beverages made of a mixture of cereals, soy and grape must: Microbiology, texture, nutritional and sensory properties. Int. J. Food Microbiol. 2012, 155, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.J.; Beer, M.U.; Butler, G. Evaluation of role of concentration and molecular weight of oat b-glucan in determining effect of viscosity on plasma glucose and insulin following an oral glucose load. Br. J. Nutr. 2000, 84, 19–23. [Google Scholar] [PubMed]
- Ai, L.; Guo, B.; Sun, K.; Chen, W.; Zhang, H.; Shao, L.; Wu, Z.; Chen, W.; Hang, F. Strain of Exopolysaccharde-Secreting Lactobacillus plantarum and Application Thereof. U.S. Patent 20140322273A1, 30 October 2014. [Google Scholar]
- Huang, T.-C.; Chan, H.-Y.; Wann, S.-Y.; Lin, F.-M.; Lee, F.-L.; Liao, C.-C. Isolated Pediococcus acidilactici 05b0111 and Method of Producing Exopolysaccharide. U.S. Patent US009873899B2, 23 January 2018. [Google Scholar]
- Gupta, M.; Bajaj, B.K. Development of fermented oat flour beverage as a potential probiotic vehicle. Food Biosci. 2017, 20, 104–109. [Google Scholar] [CrossRef]
- Matejčekovà, Z.; Liptàkovà, D.; Valík, L. Functional probiotic products based on fermented buckwheat with Lactobacillus rhamnosus. LWT Food Sci. Technol. 2017, 81, 35–41. [Google Scholar] [CrossRef]
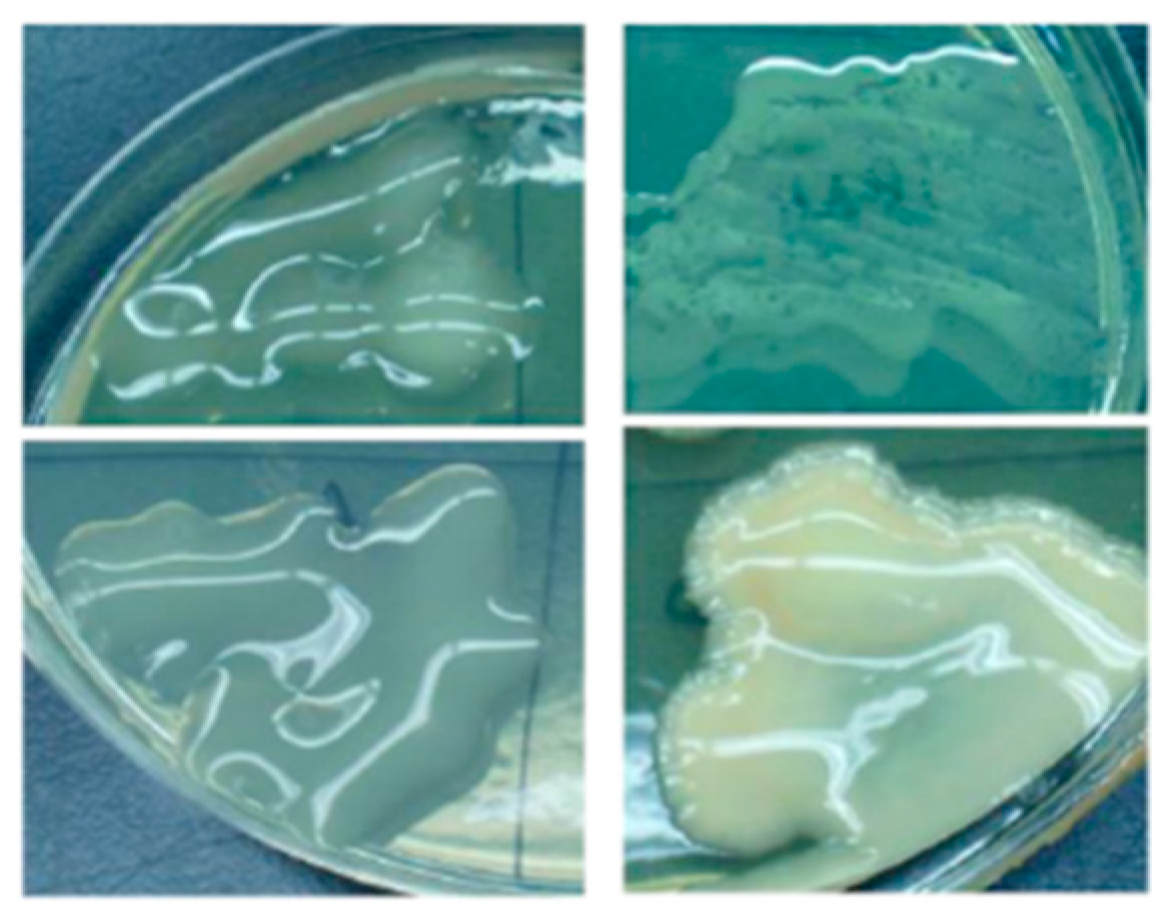

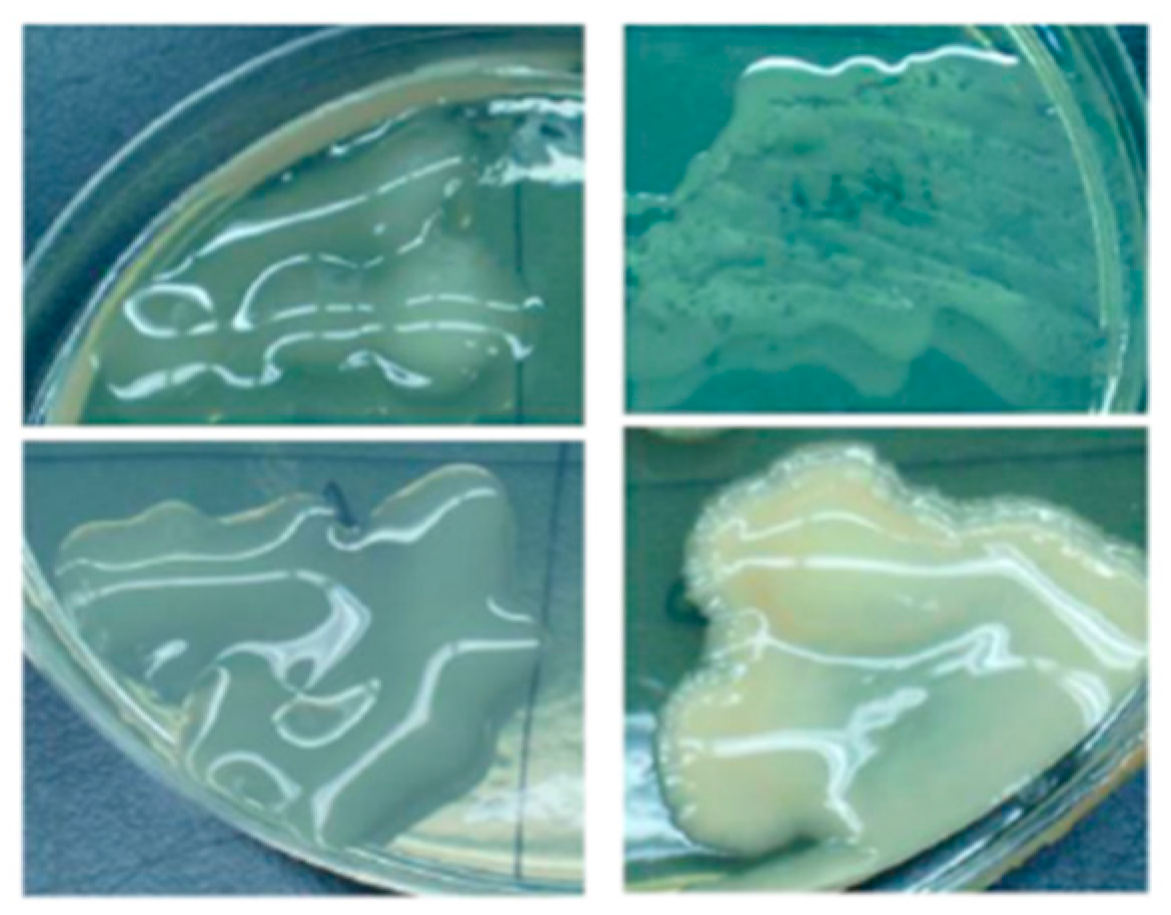{kind=link}
{kind=link}
{kind=link}
| Traditional Beverages Based on Cereals | Microorganism | Cereals | Kind of Beverage | Origins | References |
|---|---|---|---|---|---|
| amazake | Aspergillus sp. | rice | sweet fermented rice drink | Japanese | [54] |
| bhaati jaanr | LABs; yeast (Saccharomycopsis fibuligera, Rhizophus sp.) | rice | staple food beverage | Nepal, India, Bhutan | [55] |
| boza | LABs (Lactobacillus sp., Leuconostoc sp., Lactococcus sp., Pediococcus sp.); yeast (Saccharomyces cerevisiae, Candida sp., Geotrichum sp.) | wheat, rye, millet, maize | sweet colloid beverage | Bulgaria, Albania, Turkey, and Romania | [56] |
| bushera | LAB (species of Lactobacillus, Lactococcus, Leuconostoc, Enterococcus, Streptococcus, Weissella) | sorghum and Millet | non-alcholic drink | Uganda | [57] |
| chhang | LAB (1.7 × 104 such as Lactobacillus sp.) and yeast (3.5 × 104 such as Saccharomyces sp.) | rice, barley, or millet | alcholic drink | Nepal, Tibet | [58] |
| Kvass | LAB (L.casei, Leuc.mesenteroides); yeast (S. cerevisiae) | rye | fermented, non-alcholic drink | Russia | [59] |
| haria | microbial consortia (LAB, Bifidobacterium, and yeast) | rice | beverage | India | [60] |
| mangisi | LAB (9.03 × 1010 CFU/mL); yeast and mould (1.1 × 107 CFU/mL) | millet | sweet, sour, non-alcholic drink | Zimbabwe | [61] |
| Marcha | LABs (L. plantarum); yeast | rice | fermented, non-alcholic drink | India | [62] |
| ogi (or koko) | LABs (such as L. plantarum, L. pantheris, L. vaccinostercus); yeast (such as Candida krusei, Clavispora, S. cerevisiae, Rhodotorula sp.) and mould (such as Aspergillus sp., Penicillium sp.) | maize, sorghum, millet, wheat | non-alcholic drink; porridge | Nigeria, Ghana | [63] |
| omegisool | LAB (Lactobacillus sp., Pediococcus sp.) | millet | alcholic beverage | Korea | [64] |
| togwa | LABs (Lactobacillus sp., Pediococcus sp.); yeast (Candida sp., Issatchenkia orientalis, Saccharomyces cerevisiae) | maize, millet | non-alcholic drink | Africa | [65] |
| LAB strain | EPS | Substrate | Reference |
|---|---|---|---|
| L. lactis ARH74 | EPS-producer strain | quinoa | [69] |
| L. delbrueckii subsp. bulgaricus -NCFB 2772 | EPS-producer strain | oat | [72] |
| L. plantarumCCM 7039 and B. longum CCM 4990 | unknown | rice | [73] |
| L. plantarum Lp90 | EPS-producer strain | oat | [74] |
| P.damnosus | eps | oat | [75] |
| P.damnosus 2.6 | β-glucan | oat based medium | [76] |
| L.brevis G-77 | α- and β-glucan | ||
| W. confusa DSM 2019 | dextran | quinoa | [77] |
| W. cibaria WC4 | EPS-producer strain | emmer flour | [78] |
| L. rhamnosus GG | unknown | buckwheat | [79] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ripari, V. Techno-Functional Role of Exopolysaccharides in Cereal-Based, Yogurt-Like Beverages. Beverages 2019, 5, 16. https://doi.org/10.3390/beverages5010016
Ripari V. Techno-Functional Role of Exopolysaccharides in Cereal-Based, Yogurt-Like Beverages. Beverages. 2019; 5(1):16. https://doi.org/10.3390/beverages5010016
Chicago/Turabian StyleRipari, Valery. 2019. "Techno-Functional Role of Exopolysaccharides in Cereal-Based, Yogurt-Like Beverages" Beverages 5, no. 1: 16. https://doi.org/10.3390/beverages5010016
APA StyleRipari, V. (2019). Techno-Functional Role of Exopolysaccharides in Cereal-Based, Yogurt-Like Beverages. Beverages, 5(1), 16. https://doi.org/10.3390/beverages5010016