
Investigating the Transient Regenerative Potential of Cardiac Muscle Using a Neonatal Pig Partial Apical Resection Model

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Pig Model for Apex Partial Resection and Regeneration
2.2. Surgical Procedures
2.3. In Vivo Analysis of Heart Function: Echocardiogram
2.4. Gross Anatomy and Histology
2.5. Statistical Analysis
3. Results
3.1. Surgical Outcomes
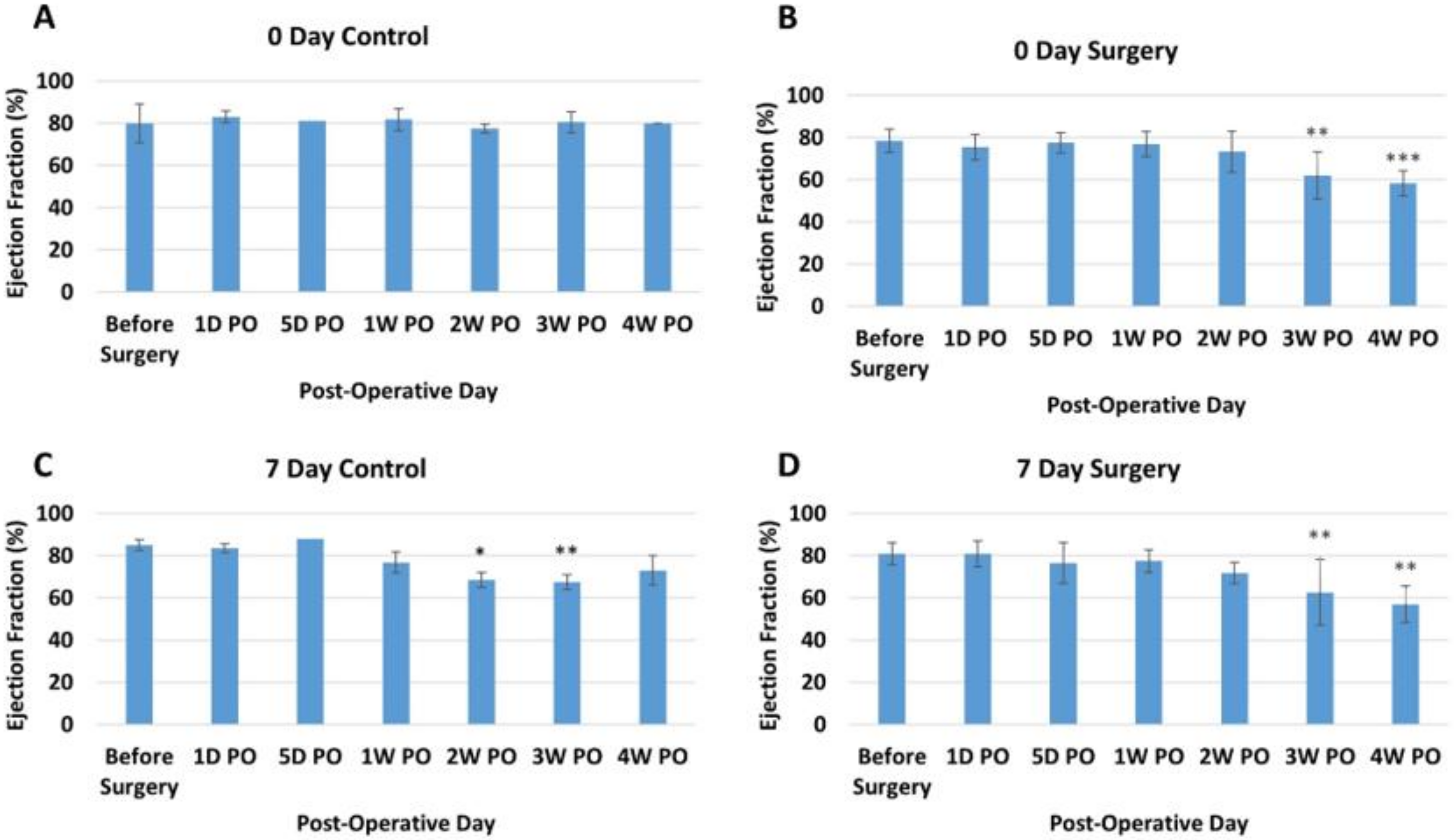
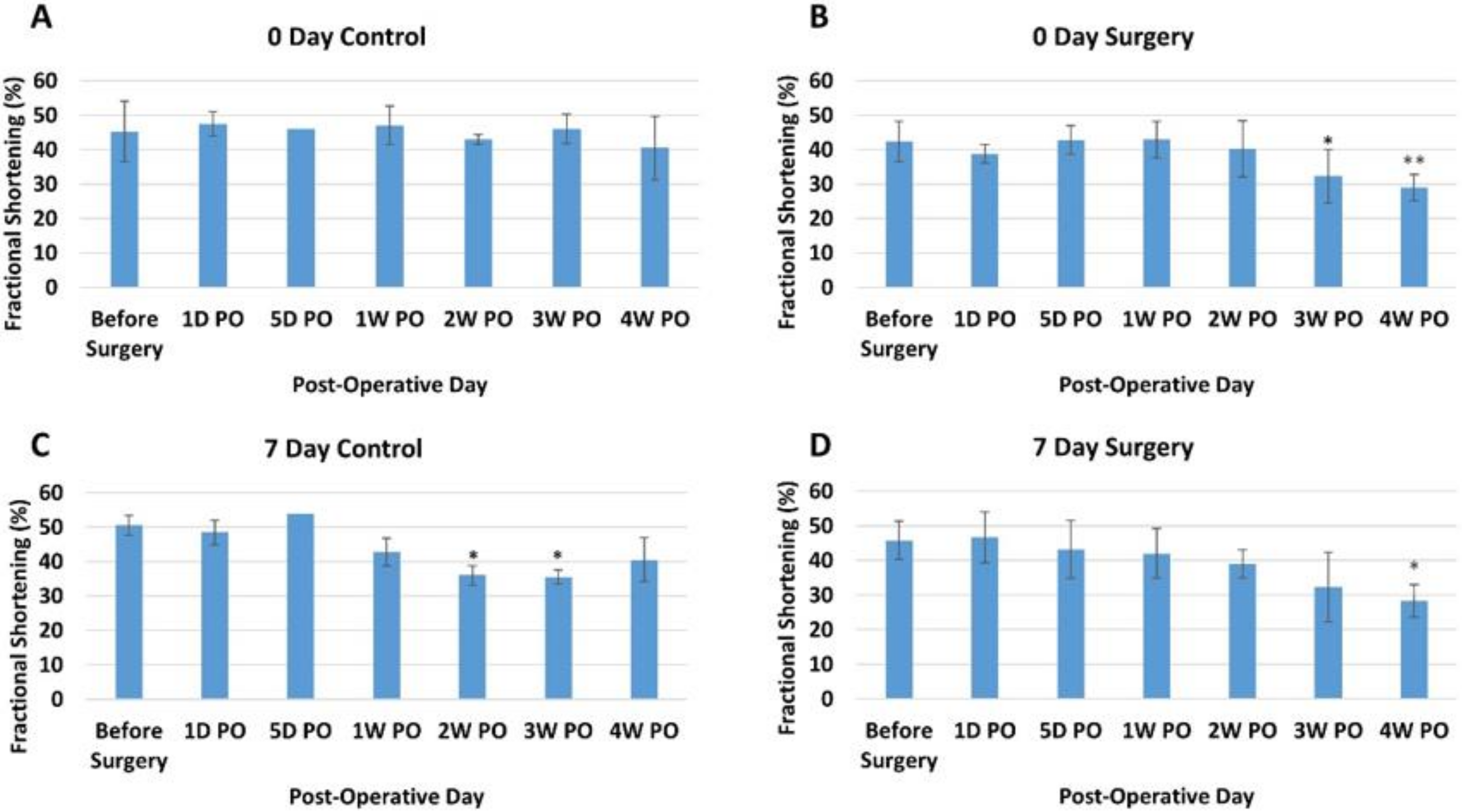
3.2. Echocardiogram Results
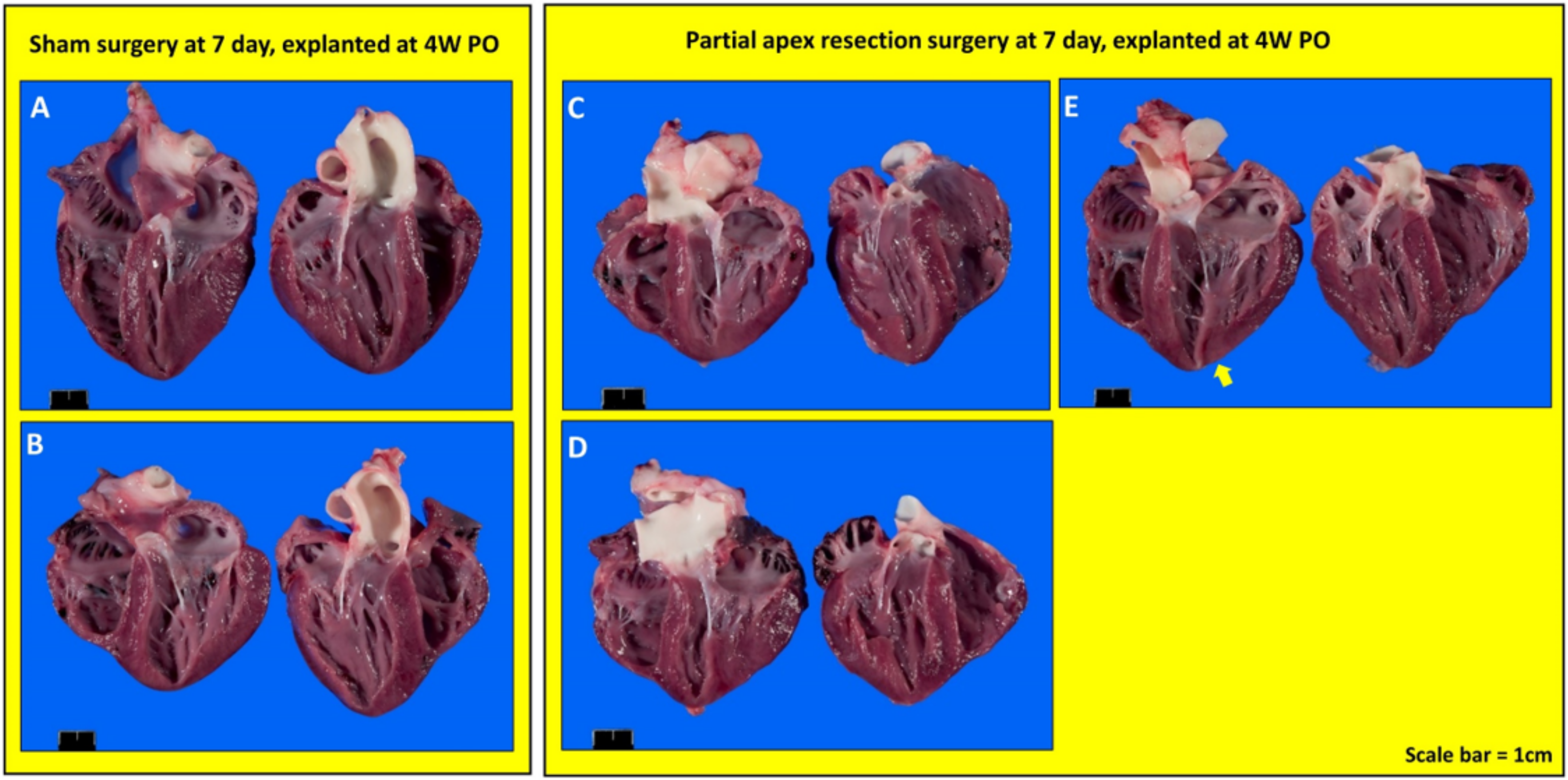
3.3. Gross Anatomy and Histology Results
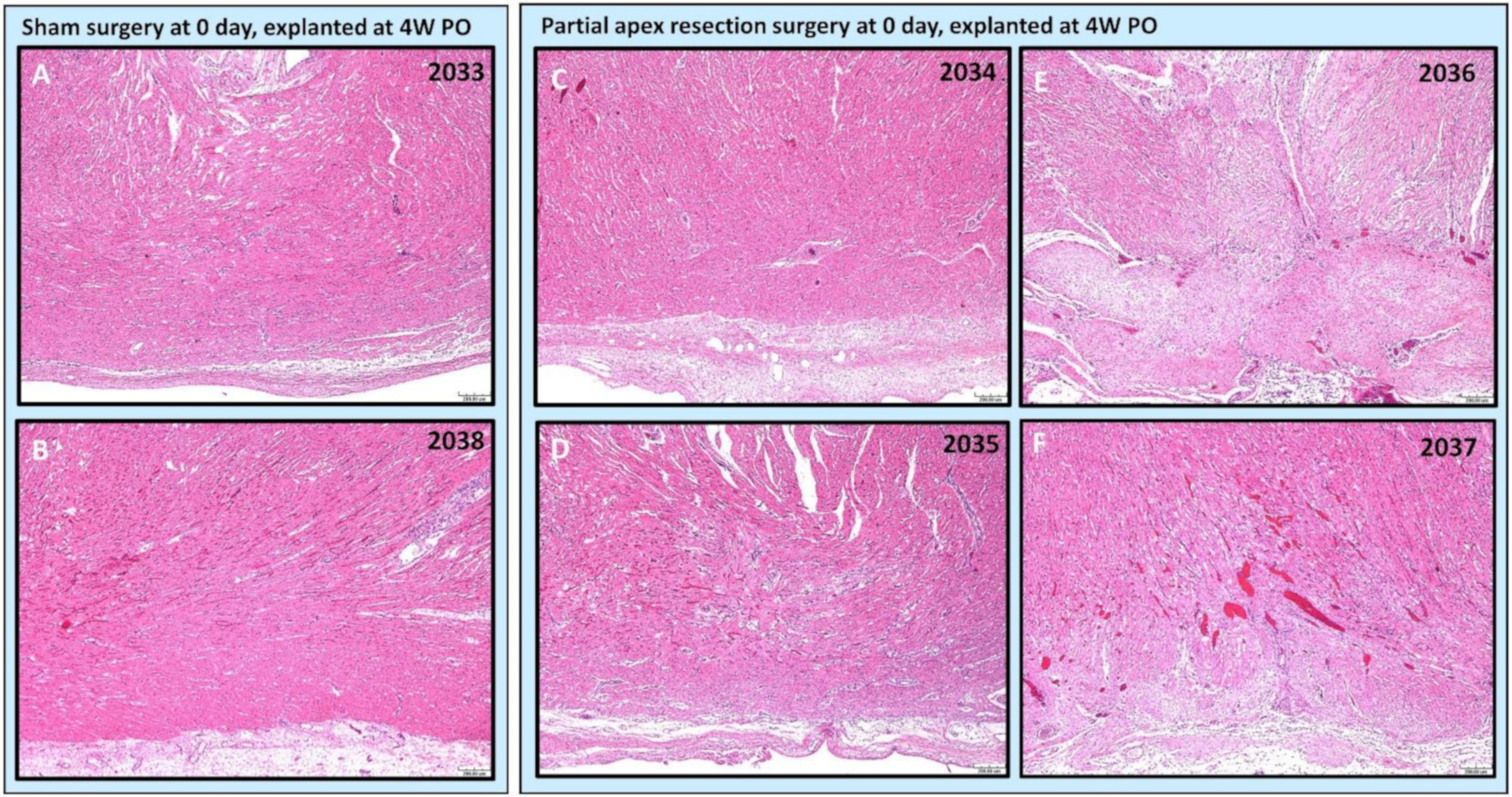
3.4. Histology
3.4.1. Comparing the Outcomes of 0-Day Surgery and Seven-Day Surgery in 4W PO Heart Explants
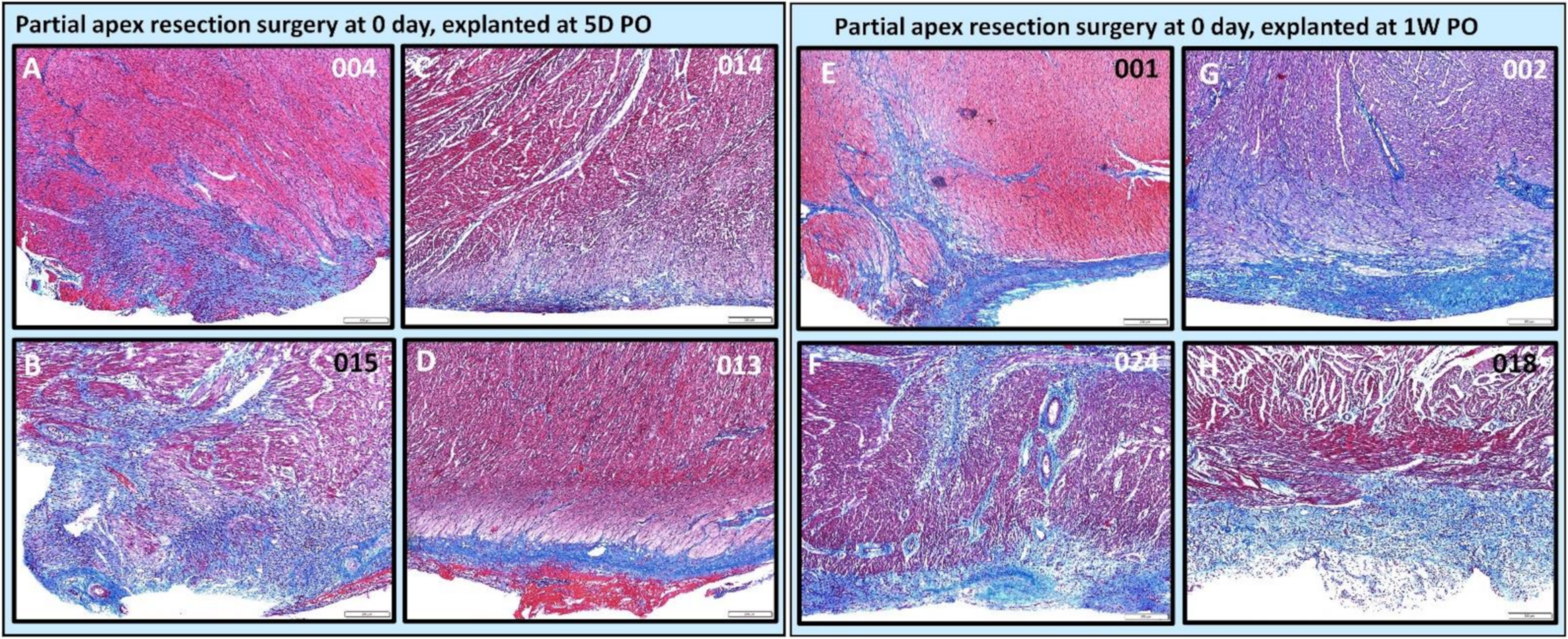
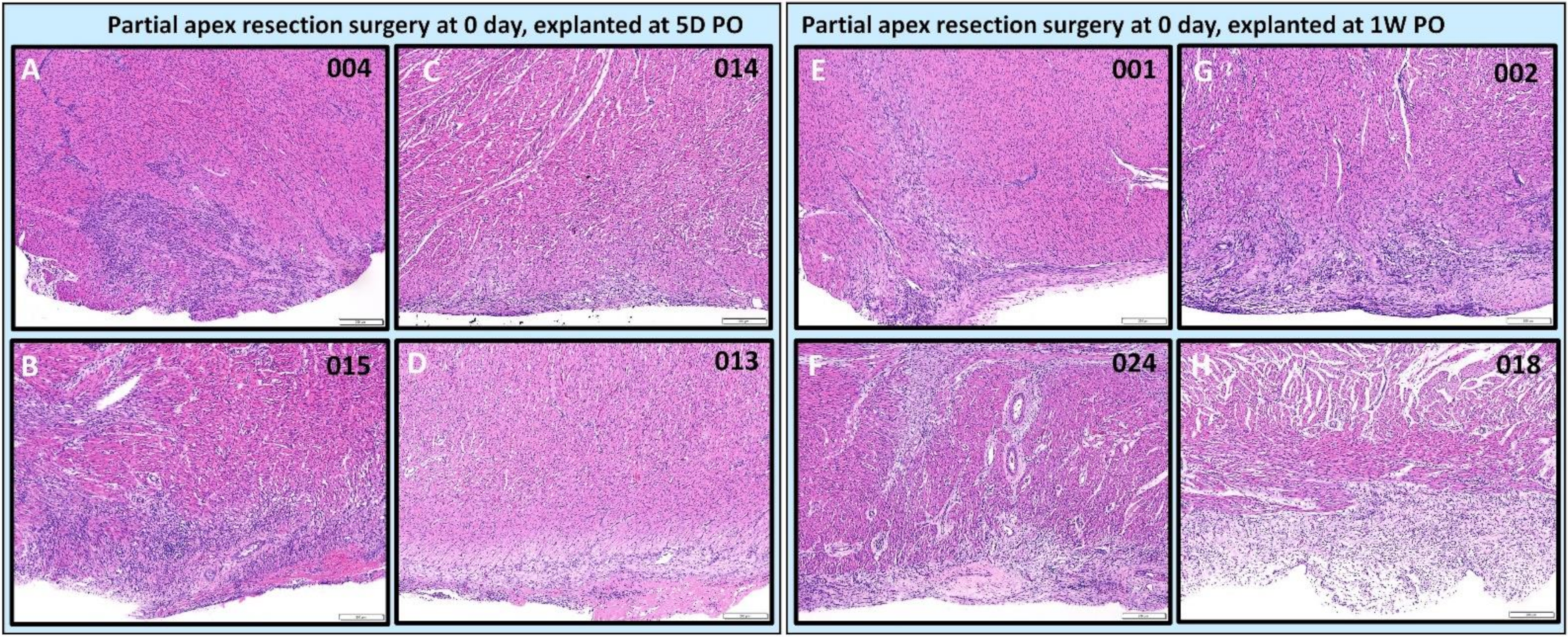
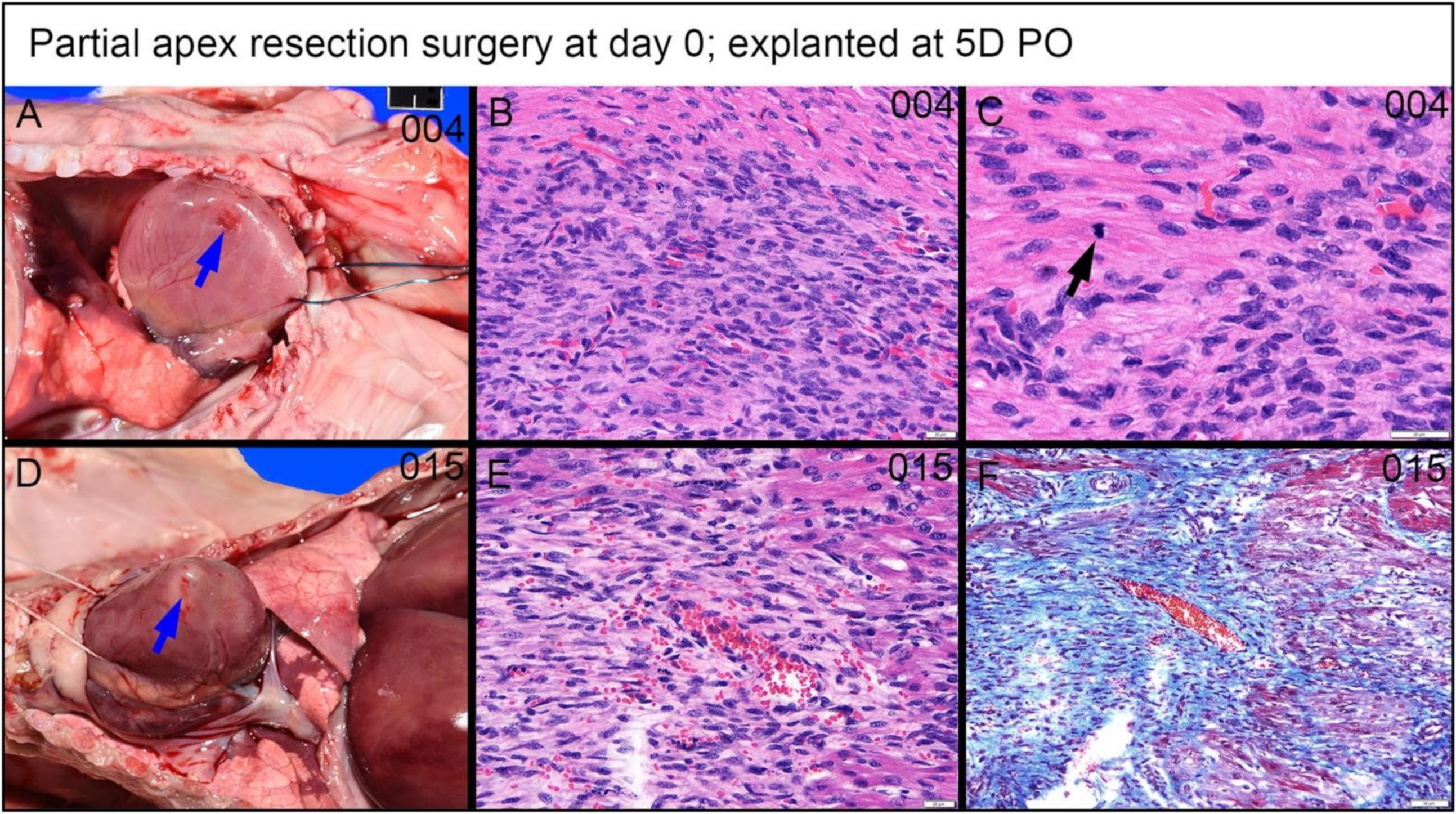
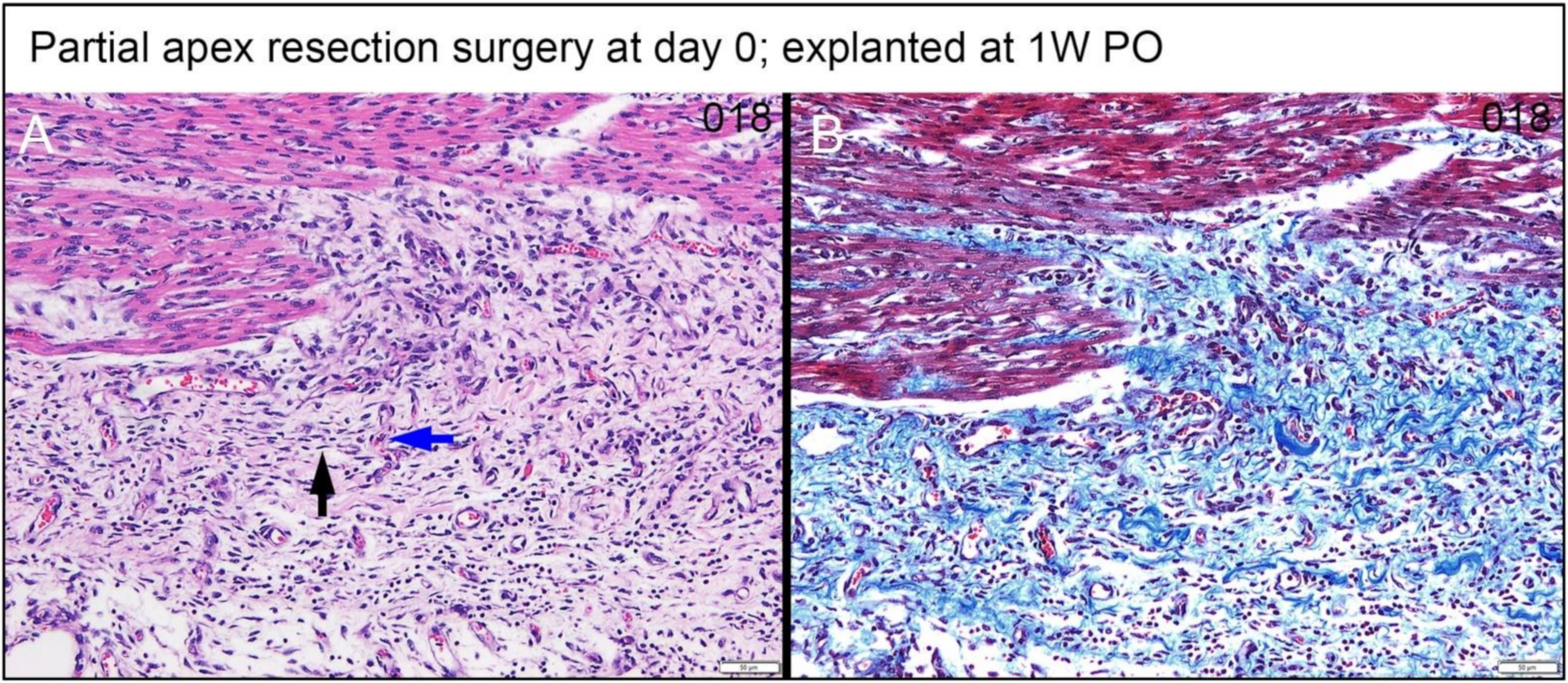
3.4.2. Assessing the Progression of Repair of the Heart Explants at 5D PO and 1W PO
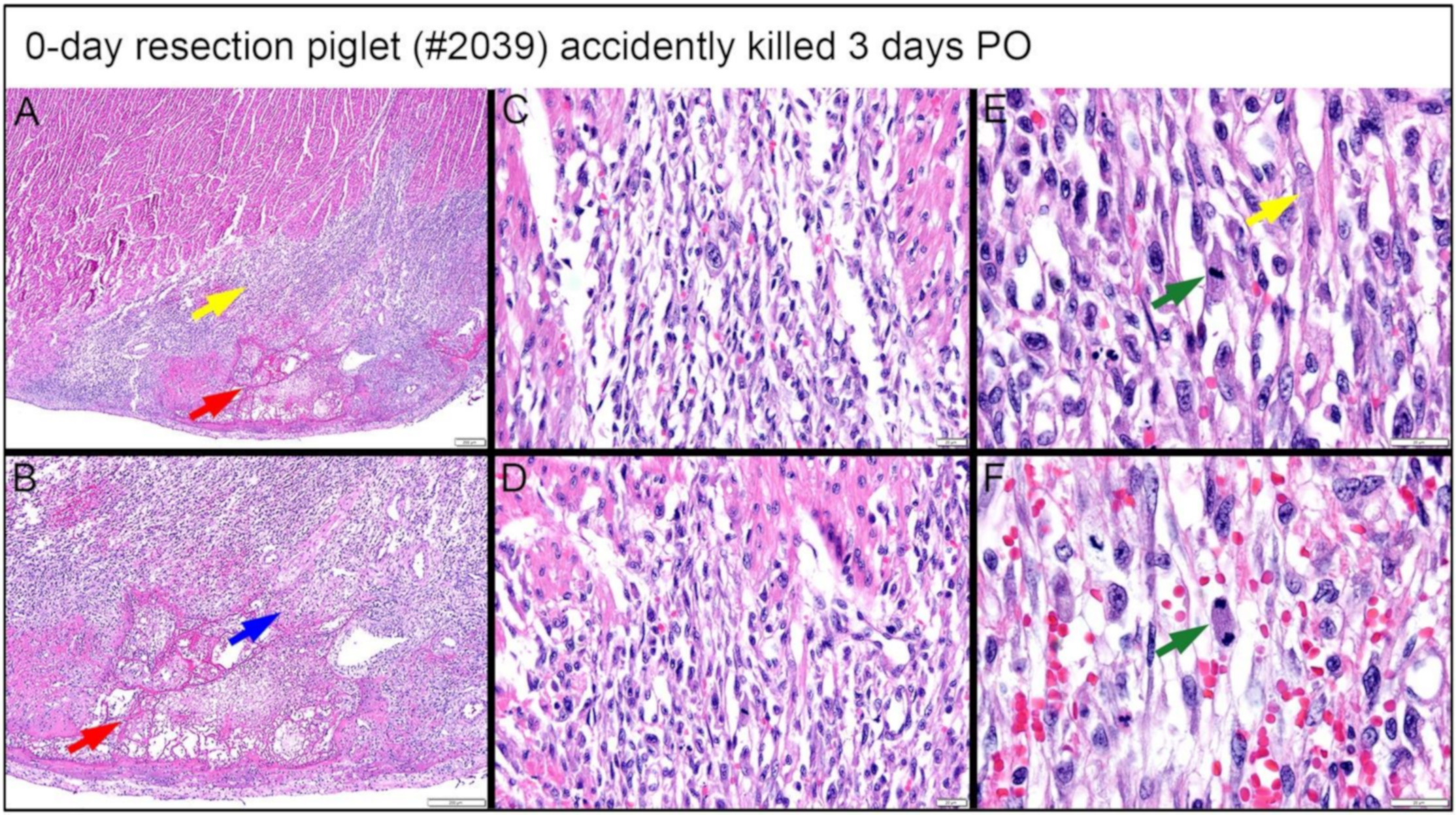
3.4.3. Dense, Proliferative Mesenchymal Cells Bordering the Fibrin and Hemorrhage Zone and Differentiating toward Immature Cardiomyocytes-3D PO Explant after 0-Day Resection Surgery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Blaha, M.J.; Dai, S.; Ford, E.S.; Fox, C.S.; Franco, S.; et al. Turner, Executive summary: Heart disease and stroke statistics—2014 update: A report from the American Heart Association. Circulation 2014, 129, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Adams, R.J.; Berry, J.D.; Brown, T.M.; Carnethon, M.R.; Dai, S.; de Simone, G.; Ford, E.S. Heart disease and stroke statistics—2011 update a report from the American Heart Association. Circulation 2011, 123, e18–e209. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, M.A.; Murry, C.E. Heart regeneration. Nature 2011, 473, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Guan, J. Cellular cardiomyoplasty and cardiac tissue engineering for myocardial therapy. Adv. Drug Deliv. Rev. 2010, 62, 784–797. [Google Scholar] [CrossRef]
- Herrmann, J.L.; Abarbanell, A.M.; Weil, B.R.; Wang, Y.; Wang, M.; Tan, J.; Meldrum, D.R. Cell-based therapy for ischemic heart disease: A clinical update. Ann. Thorac. Surg. 2009, 88, 1714–1722. [Google Scholar] [CrossRef]
- Forrester, J.S.; Makkar, R.R.; Marbán, E. Long-Term Outcome of Stem Cell Therapy for Acute Myocardial Infarction: Right Results, Wrong Reasons. J. Am. Coll. Cardiol. 2009, 53, 2270–2272. [Google Scholar] [CrossRef][Green Version]
- Jameel, M.N.; Zhang, J. Heart failure management: The present and the future. Antioxid. Redox Signal. 2009, 11, 1989–2010. [Google Scholar] [CrossRef]
- Quevedo, H.C.; Hatzistergos, K.E.; Oskouei, B.N.; Feigenbaum, G.S.; Rodriguez, J.E.; Valdes, D.; Pattany, P.M.; Zambrano, J.P.; Hu, Q.; McNiece, I.; et al. Allogeneic mesenchymal stem cells restore cardiac function in chronic ischemic cardiomyopathy via trilineage differentiating capacity. Proc. Natl. Acad. Sci. USA 2009, 106, 14022–14027. [Google Scholar] [CrossRef]
- Malliaras, K.; Marbán, E. Cardiac cell therapy: Where we’ve been, where we are, and where we should be headed. Br. Med. Bull. 2011, 98, 161–185. [Google Scholar] [CrossRef]
- Nguyen, P.K.; Nag, D.; Wu, J.C. Methods to assess stem cell lineage, fate and function. Adv. Drug Deliv. Rev. 2010, 62, 1175–1186. [Google Scholar] [CrossRef]
- Bursac, N. Cardiac tissue engineering using stem cells [Cellular/Tissue Engineering]. IEEE Comput. Graph. Appl. 2009, 28, 80, 82, 84–86; 88–89. [Google Scholar] [CrossRef]
- Vunjak-Novakovic, G.; Tandon, N.; Godier, A.; Maidhof, R.; Marsano, A.; Martens, T.P.; Radisic, M. Challenges in cardiac tissue engineering. Tissue Eng. Part B Rev. 2009, 16, 169–187. [Google Scholar] [CrossRef]
- Reilly, G.C.; Engler, A.J. Intrinsic extracellular matrix properties regulate stem cell differentiation. J. Biomech. 2010, 43, 55–62. [Google Scholar] [CrossRef]
- Conrad, C.; Niess, H.; Huss, R.; Huber, S.; Von Luettichau, I.; Nelson, P.J.; Ott, H.C.; Jauch, K.-W.; Bruns, C.J. Multipotent mesenchymal stem cells acquire a lymphendothelial phenotype and enhance lymphatic regeneration in vivo. Circulation 2009, 119, 281–289. [Google Scholar] [CrossRef]
- Friedland, J.C.; Lee, M.H.; Boettiger, D. Mechanically activated integrin switch controls α5β1 function. Science 2009, 323, 642–644. [Google Scholar] [CrossRef]
- Martino, M.; Mochizuki, M.; Rothenfluh, D.A.; Rempel, S.A.; Hubbell, J.A.; Barker, T.H. Controlling integrin specificity and stem cell differentiation in 2D and 3D environments through regulation of fibronectin domain stability. Biomaterials 2009, 30, 1089–1097. [Google Scholar] [CrossRef]
- Rowlands, A.S.; George, P.A.; Cooper-White, J.J. Directing osteogenic and myogenic differentiation of MSCs: Interplay of stiffness and adhesive ligand presentation. Am. J. Physiol. Cell Physiol. 2008, 295, C1037–C1044. [Google Scholar] [CrossRef]
- Yasunaga, M.; Tada, S.; Torikai-Nishikawa, S.; Nakano, Y.; Okada, M.; Jakt, L.M.; Chiba, T.; Era, T.; Nishikawa, S.-I. Induction and monitoring of definitive and visceral endoderm differentiation of mouse ES cells. Nat. Biotechnol. 2005, 23, 1542–1550. [Google Scholar] [CrossRef]
- Ruiz, S.A.; Chen, C.S. Emergence of patterned stem cell differentiation within multicellular structures. Stem Cells 2008, 26, 2921–2927. [Google Scholar] [CrossRef]
- Stevens, M.M.; George, J.H. Exploring and Engineering the Cell Surface Interface. Science 2005, 310, 1135–1138. [Google Scholar] [CrossRef]
- Berry, M.F.; Engler, A.J.; Woo, Y.J.; Pirolli, T.J.; Bish, L.T.; Jayasankar, V.; Morine, K.J.; Gardner, T.J.; Discher, D.; Sweeney, H.L. Mesenchymal stem cell injection after myocardial infarction improves myocardial compliance. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H2196–H2203. [Google Scholar] [CrossRef]
- Burdick, J.A.; Vunjak-Novakovic, G. Engineered microenvironments for controlled stem cell differentiation. Tissue Eng. Part A 2008, 15, 205–219. [Google Scholar] [CrossRef]
- Discher, D.E.; Mooney, D.J.; Zandstra, P.W. Growth Factors, Matrices, and Forces Combine and Control Stem Cells. Science 2009, 324, 1673–1677. [Google Scholar] [CrossRef]
- Winer, J.P.; Janmey, P.A.; McCormick, M.E.; Funaki, M. Bone Marrow-Derived Human Mesenchymal Stem Cells Become Quiescent on Soft Substrates but Remain Responsive to Chemical or Mechanical Stimuli. Tissue Eng. Part A 2008, 15, 147–154. [Google Scholar] [CrossRef]
- Huebsch, N.; Arany, P.R.; Mao, A.S.; Shvartsman, D.; Ali, O.A.; Bencherif, S.A.; Rivera-Feliciano, J.; Mooney, D.J. Harnessing traction-mediated manipulation of the cell/matrix interface to control stem-cell fate. Nat. Mater. 2010, 9, 518–526. [Google Scholar] [CrossRef]
- Guan, J.; Wang, F.; Li, Z.; Chen, J.; Guo, X.; Liao, J.; Moldovan, N.I. The stimulation of the cardiac differentiation of mesenchymal stem cells in tissue constructs that mimic myocardium structure and biomechanics. Biomaterials 2011, 32, 5568–5580. [Google Scholar] [CrossRef]
- Li, Z.; Guo, X.; Matsushita, S.; Guan, J. Differentiation of cardiosphere-derived cells into a mature cardiac lineage using biodegradable poly(N-isopropylacrylamide) hydrogels. Biomaterials 2011, 32, 3220–3232. [Google Scholar] [CrossRef]
- Li, Z.; Guo, X.; Palmer, A.F.; Das, H.; Guan, J. High-efficiency matrix modulus-induced cardiac differentiation of human mesenchymal stem cells inside a thermosensitive hydrogel. Acta Biomater. 2012, 8, 3586–3595. [Google Scholar] [CrossRef]
- Kraehenbuehl, T.P.; Zammaretti, P.; Van der Vlies, A.J.; Schoenmakers, R.G.; Lutolf, M.P.; Jaconi, M.E.; Hubbell, J.A. Three-dimensional extracellular matrix-directed cardioprogenitor differentiation: Systematic modulation of a synthetic cell-responsive PEG-hydrogel. Biomaterials 2008, 29, 2757–2766. [Google Scholar] [CrossRef]
- Hashimoto, H.; Olson, E.N.; Bassel-Duby, R. Therapeutic approaches for cardiac regeneration and repair. Nat. Rev. Cardiol. 2018, 15, 585–600. [Google Scholar] [CrossRef]
- Tzahor, E.; Poss, K.D. Cardiac regeneration strategies: Staying young at heart. Science 2017, 356, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Sahara, M.; Santoro, F.; Chien, K.R. Programming and reprogramming a human heart cell. EMBO J. 2015, 34, 710–738. [Google Scholar] [CrossRef] [PubMed]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef] [PubMed]
- Raya, Á.; Koth, C.M.; Büscher, D.; Kawakami, Y.; Itoh, T.; Raya, R.M.; Sternik, G.; Tsai, H.-J.; Rodríguez-Esteban, C.; Izpisúa-Belmonte, J.C. Activation of Notch signaling pathway precedes heart regeneration in zebrafish. Proc. Natl. Acad. Sci. USA 2003, 100 (Suppl. 1), 11889–11895. [Google Scholar] [CrossRef]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Hill, J.A.; Richardson, J.A.; Olson, E.N.; Sadek, H.A. Transient Regenerative Potential of the Neonatal Mouse Heart. Science 2011, 331, 1078–1080. [Google Scholar] [CrossRef]
- Andersen, D.C.; Ganesalingam, S.; Jensen, C.H.; Sheikh, S.P. Do Neonatal Mouse Hearts Regenerate following Heart Apex Resection? Stem Cell Rep. 2014, 2, 406–413. [Google Scholar] [CrossRef]
- Forbes, S.J.; Rosenthal, N. Preparing the ground for tissue regeneration: From mechanism to therapy. Nat. Med. 2014, 20, 857–869. [Google Scholar] [CrossRef]
- Mercola, M.; Ruiz-Lozano, P.; Schneider, M.D. Cardiac muscle regeneration: Lessons from development. Genes Dev. 2011, 25, 299–309. [Google Scholar] [CrossRef]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for Cardiomyocyte Renewal in Humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef]
- Kajstura, J.; Urbanek, K.; Perl, S.; Hosoda, T.; Zheng, H.; Ogórek, B.; Ferreira-Martins, J.; Goichberg, P.; Rondon-Clavo, C.; Sanada, F.; et al. Cardiomyogenesis in the Adult Human Heart. Circ. Res. 2010, 107, 305–315. [Google Scholar] [CrossRef]
- Steinhauser, M.L.; Lee, R.T. Regeneration of the heart. EMBO Mol. Med. 2011, 3, 701–712. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, E.; Zhao, M.; Chong, Z.; Fan, C.; Tang, Y.; Hunter, J.D.; Borovjagin, A.V.; Walcott, G.P.; Chen, J.Y.; et al. Regenerative Potential of Neonatal Porcine Hearts. Circulation 2018, 138, 2809–2816. [Google Scholar] [CrossRef]
- Ye, L.; D’Agostino, G.; Loo, S.J.; Wang, C.X.; Su, L.P.; Tan, S.H.; Tee, G.Z.; Pua, C.J.; Pena, E.M.; Cheng, R.B.; et al. Early Regenerative Capacity in the Porcine Heart. Circulation 2018, 138, 2798–2808. [Google Scholar] [CrossRef]
- Malliaras, K.; Polychronopoulou, E.; Poulakida, I.; Sagris, D.; Makaritsis, K. Limited regenerative potential of the neonatal porcine heart after myocardial infarction. Eur. Heart J. 2019, 40, ehz746.0349. [Google Scholar] [CrossRef]
- Weaver, M.E.; Pantely, G.A.; Bristow, J.D.; Ladley, H.D. A quantitative study of the anatomy and distribution of coronary arteries in swine in comparison with other animals and man. Cardiovasc. Res. 1986, 20, 907–917. [Google Scholar] [CrossRef]
- Zaragoza, C.; Gomez-Guerrero, C.; Martin-Ventura, J.L.; Blanco-Colio, L.; Lavin, B.; Mallavia, B.; Tarin, C.; Mas, S.; Ortiz, A.; Egido, J. Animal models of cardiovascular diseases. BioMed Res. Int. 2011, 2011, 497841. [Google Scholar] [CrossRef]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef]
- Crick, S.J.; Sheppard, M.N.; Ho, S.Y.; Gebstein, L.; Anderson, R.H. Anatomy of the pig heart: Comparisons with normal human cardiac structure. J. Anat. 1998, 193, 105–119. [Google Scholar] [CrossRef]
- Fugelseth, D.; Satas, S.; Runde, M.; Hågå, P.; Thoresen, M. Cardiac function and morphology studied by two-dimensional Doppler echocardiography in unsedated newborn pigs. Exp. Physiol. 1999, 84, 69–78. [Google Scholar] [CrossRef]
- Wang, B.; Tedder, M.E.; Perez, C.E.; Wang, G.; Curry, A.L.D.J.; To, F.; Elder, S.H.; Williams, L.N.; Simionescu, D.T.; Liao, J. Structural and biomechanical characterizations of porcine myocardial extracellular matrix. J. Mater. Sci. Mater. Med. 2012, 23, 1835–1847. [Google Scholar] [CrossRef]
- Wang, B.; Williams, L.N.; Curry, A.L.D.J.; Liao, J. Preparation of acellular myocardial scaffolds with well-preserved cardiomyocyte lacunae, and method for applying mechanical and electrical simulation to tissue construct. In Cardiac Tissue Engineering; Humana Press: New York, NY, USA, 2014; Volume 1181, pp. 189–202. [Google Scholar]
- Zhang, S.; Crow, J.A.; Yang, X.; Chen, J.; Borazjani, A.; Mullins, K.B.; Chen, W.; Cooper, R.C.; McLaughlin, R.M.; Liao, J. The Correlation of 3D DT-MRI Fiber Disruption with Structural and Mechanical Degeneration in Porcine Myocardium. Ann. Biomed. Eng. 2010, 38, 3084–3095. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Numbers and Time Points for Heart Harvest | |||
|---|---|---|---|
| Total N = 36 | Heart Harvest 5 Day Post-Surgery | Heart Harvest 1 Week Post-Surgery | Heart Harvest 4 Weeks Post-Surgery |
| Apex Removal Surgery on 0-day-old Piglets | N = 4 | N = 4 | N = 5 |
| Apex Removal Surgery on 7-day-old Piglets | N = 5 | N = 5 | N = 5 |
| Sham Surgery on 0-day-old Piglets | NA | N = 2 | N = 2 |
| Sham Surgery on 7-day-old Piglets | NA | N = 2 | N = 2 |
| 0 Day Control and 0 Day Surgery | ||||
| Piglet ID | Sex of Piglet | Body Weight at Surgery (lbs) | Tissue Mass Removed from Apical Region (mg) | Age at Time of Apical Resection |
| 2033 | M | 3.31 | 0 day control | NA |
| 2038 | F | 2.81 | 0 day control | NA |
| 005 | F | 3.53 | 0 day control | NA |
| 017 | M | 2.20 | 0 day control | NA |
| 2034 | M | 4.31 | *, sample lost in pig (cut similar to 2035) | 0 days |
| 2035 | F | 3.31 | 5.1 | 0 days |
| 2036 | M | 3.69 | 22.6 | 0 days |
| 2039 | M | 3.79 | 34.5 (accidently killed by the mother sow 3 days PO) | 0 days |
| 2037 | F | 3.56 | *, sample lost in pig (cut similar to 2039) | 0 days |
| 004 | M | 3.09 | 2.3 | 0 days |
| 013 | F | 3.00 | 2.2 | 0 days |
| 014 | M | 4.37 | 16.8 | 0 days |
| 015 | F | 4.10 | 3.1 | 0 days |
| 001 | F | 3.60 | 2.9 | 0 days |
| 002 | F | 3.75 | 2.6 | 0 days |
| 018 | F | 3.53 | 0.8 | 0 days |
| 024 | M | 3.09 | 0.3 | 0 days |
| AVG ± STD | 3.64 ± 0.48 | 8.47 ± 11.20 | ||
| 7 Day Control and 7 Day Surgery | ||||
| Piglet ID | Sex of Piglet | Body Weight at Surgery (lbs) | Tissue Mass Removed from Apical Region (mg) | Age at Time of Apical Resection |
| 2071 | F | 6.31 | 7 day control | NA |
| 2077 | M | 3.42 | 7 day control | NA |
| 012 | F | 5.51 | 7 day control | NA |
| 027 | M | 6.59 | 7 day control | NA |
| 2073 | M | 3.09 | 13.0 | 7 days |
| 2075 | M | 4.41 | 6.0 | 7 days |
| 2076 | M | 7.48 | 8.4 | 7 days |
| 2078 * | M | 4.86 | *, heart tore during surgery; euthanized-massive blood loss | 7 days |
| 2079 * | F | 5.29 | *, resected into ventricle; euthanized-massive blood loss | 7 days |
| 007 | F | 5.07 | 6.7 | 7 days |
| 008 | M | 5.07 | 5.2 | 7 days |
| 029 | F | 7.74 | 12.2 | 7 days |
| 031 | M | 7.61 | 2.3 | 7 days |
| 009 | M | 5.07 | 9.9 | 7 days |
| 010 | F | 6.17 | 7.3 | 7 days |
| 011 | M | 5.07 | 2.4 | 7 days |
| 016 | F | 6.72 | 11.5 | 7 days |
| 026 | M | 7.34 | 6.5 | 7 days |
| 028 | M | 8.07 | 9.0 | 7 days |
| AVG ± STD | 6.39 ± 1.19 | 5.62 ± 1.75 | ||
| Piglet ID and Group | Body Weight at Euthanasia (lbs) | Heart Mass at Euthanasia (g) | Age at Euthanasia |
|---|---|---|---|
| 2033 (0 day control) | 13.28 | 35.73 | 4 weeks |
| 2038 (0 day control) | 13.88 | 56.70 | 4 weeks |
| AVG ± STD | 13.58 ± 0.30 | 46.22 ± 10.49 | ---------- |
| 2034 (0 day surgery) | 14.52 | 42.52 | 4 weeks |
| 2035 (0 day surgery) | 15.40 | 39.69 | 4 weeks |
| 2036 (0 day surgery) | 15.66 | 42.52 | 4 weeks |
| 2037 (0 day surgery) | 16.24 | 48.19 | 4 weeks |
| AVG ± STD | 15.46 ± 0.62 | 43.23 ± 3.09 | ---------- |
| 2039 * (0 day surgery) | ----------------------- | ------------------ | Lost at 3 days |
| 2071 (7 day control) | 16.78 | 51.03 | 5 weeks |
| 2077 (7 day control) | 16.82 | 48.19 | 5 weeks |
| AVG ± STD | 16.80 ± 0.02 | 49.61 ± 1.42 | ---------- |
| 2073 (7 day surgery) | 9.48 | 25.51 | 5 weeks |
| 2075 (7 day surgery) | 11.16 | 31.18 | 5 weeks |
| 2076 (7 day surgery) | 21.52 | 56.70 | 5 weeks |
| AVG ± STD | 14.05 ± 5.32 | 37.80 ± 13.57 | ---------- |
| 2078 * (7 day surgery) | ----------------------- | ------------------ | Lost during surgery |
| 2079 * (7 day surgery) | ----------------------- | ------------------ | Lost during surgery |
| 005 (0 day control) | 5.82 | 26.48 | 1 week |
| 017 (0 day control) | 4.08 | 18.92 | 1 week |
| AVG ± STD | 4.95 ± 0.87 | 22.70 ± 3.78 | ---------- |
| 004 (0 day surgery) | 4.02 | 14.83 | 5 days |
| 013 (0 day surgery) | 4.23 | 17.67 | 5 days |
| 014 (0 day surgery) | 6.48 | 26.73 | 5 days |
| 015 (0 day surgery) | 5.86 | 28.00 | 5 days |
| AVG ± STD | 5.15 ± 1.05 | 21.81 ± 5.67 | ---------- |
| 001 (0 day surgery) | 5.46 | 20.01 | 1 week |
| 002 (0 day surgery) | 5.30 | 22.82 | 1 week |
| 018 (0 day surgery) | 5.86 | 34.91 | 1 week |
| 024 (0 day surgery) | 5.20 | 28.57 | 1 week |
| AVG ± STD | 5.46 ± 0.25 | 26.57 ± 5.72 | ---------- |
| 012 (7 day control) | 10.10 | 31.16 | 2 week |
| 027 (7 day control) | 9.88 | 31.52 | 2 week |
| AVG ± STD | 9.99 ± 0.11 | 31.34 ± 0.18 | ---------- |
| 007 (7 day surgery) | 7.64 | 24.71 | 12 days |
| 008 (7 day surgery) | 6.92 | 22.22 | 12 days |
| 011 (7 day surgery) | 9.06 | 39.72 | 12 days |
| 029 (7 day surgery) | 7.74 | 34.27 | 12 days |
| 031 (7 day surgery) | 7.61 | 34.14 | 12 days |
| AVG ± STD | 7.79 ± 0.70 | 31.01 ± 6.53 | ---------- |
| 009 (7 day surgery) | 8.30 | 34.54 | 2 week |
| 010 (7 day surgery) | 10.34 | 34.36 | 2 week |
| 016 (7 day surgery) | 6.72 | 37.80 | 2 week |
| 026 (7 day surgery) | 7.34 | 42.16 | 2 week |
| 028 (7 day surgery) | 8.07 | 44.11 | 2 week |
| AVG ± STD | 8.15 ± 1.23 | 38.59 ± 3.95 | ---------- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copeland, K.M.; Brazile, B.L.; Butler, J.R.; Cooley, J.; Brinkman-Ferguson, E.; Claude, A.; Lin, S.; Rais-Rohani, S.; Welch, B.; McMahan, S.R.; et al. Investigating the Transient Regenerative Potential of Cardiac Muscle Using a Neonatal Pig Partial Apical Resection Model. Bioengineering 2022, 9, 401. https://doi.org/10.3390/bioengineering9080401
Copeland KM, Brazile BL, Butler JR, Cooley J, Brinkman-Ferguson E, Claude A, Lin S, Rais-Rohani S, Welch B, McMahan SR, et al. Investigating the Transient Regenerative Potential of Cardiac Muscle Using a Neonatal Pig Partial Apical Resection Model. Bioengineering. 2022; 9(8):401. https://doi.org/10.3390/bioengineering9080401
Chicago/Turabian StyleCopeland, Katherine M., Bryn L. Brazile, J. Ryan Butler, Jim Cooley, Erin Brinkman-Ferguson, Andrew Claude, Sallie Lin, Sammira Rais-Rohani, Bradley Welch, Sara R. McMahan, and et al. 2022. "Investigating the Transient Regenerative Potential of Cardiac Muscle Using a Neonatal Pig Partial Apical Resection Model" Bioengineering 9, no. 8: 401. https://doi.org/10.3390/bioengineering9080401
APA StyleCopeland, K. M., Brazile, B. L., Butler, J. R., Cooley, J., Brinkman-Ferguson, E., Claude, A., Lin, S., Rais-Rohani, S., Welch, B., McMahan, S. R., Nguyen, K. T., Hong, Y., Ramaswamy, S., Liu, Z.-P., Bajona, P., Peltz, M., & Liao, J. (2022). Investigating the Transient Regenerative Potential of Cardiac Muscle Using a Neonatal Pig Partial Apical Resection Model. Bioengineering, 9(8), 401. https://doi.org/10.3390/bioengineering9080401