
Diet Shift May Trigger LuxS/AI-2 Quorum Sensing in Rumen Bacteria


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sample Collection
2.3. RNA Extraction, LuxS/AI-2 Quorum Sensing, and Biofilm Formation Assay
2.4. Serum Indicators and Rumen Volatile Fatty Acids Determination
2.5. DNA Extraction, Sequencing, and Data Analysis
2.6. Statistical Analysis
3. Results
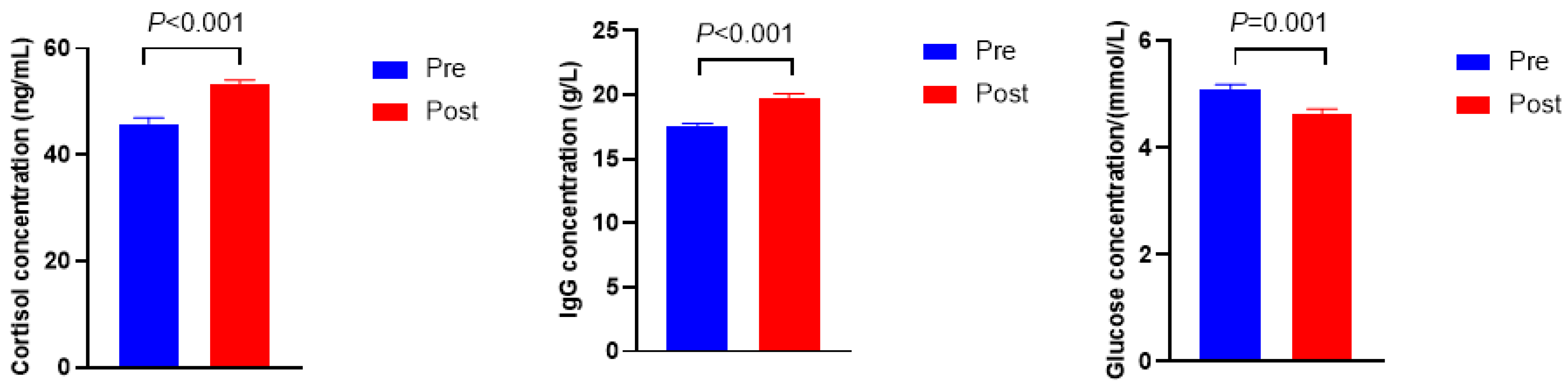
3.1. Serum Biochemical, Immune, and Hormonal Indicators
3.2. AI-2 Concentration and luxS Gene Expression
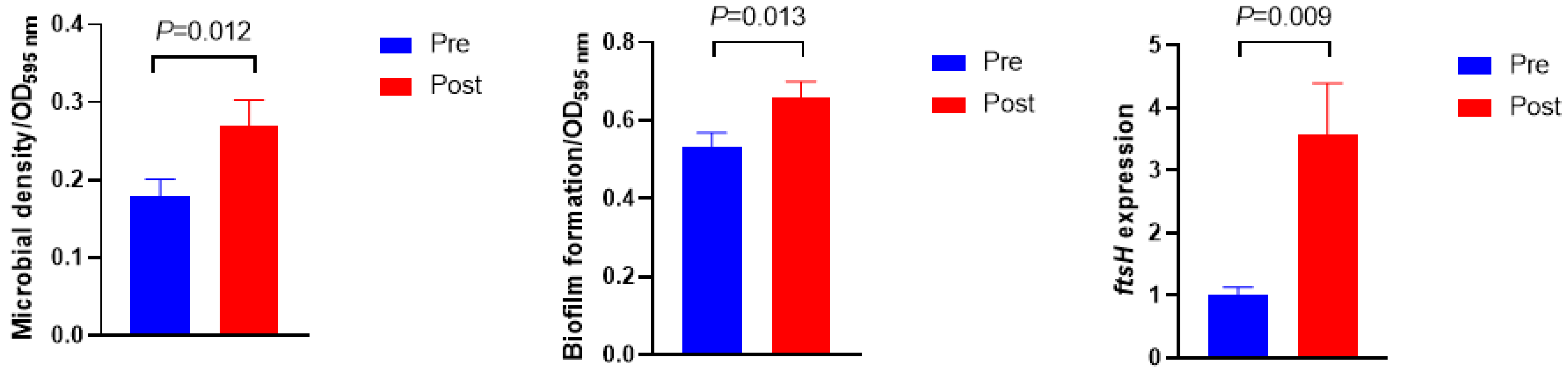
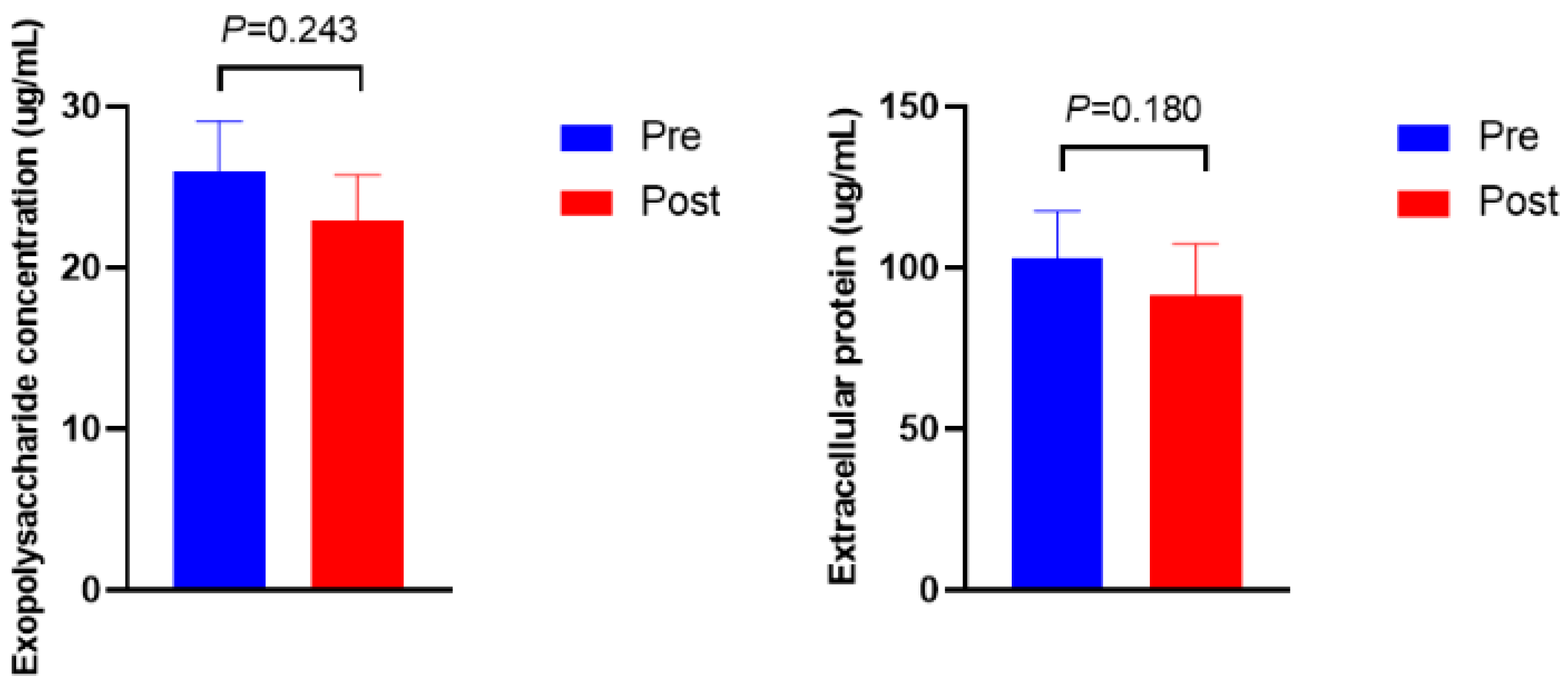
3.3. Microbial Density, Biofilm Formation, ftsH Gene Expression, and Extracellular Polymeric Substances Composition
3.4. Rumen Fermentation Characteristics
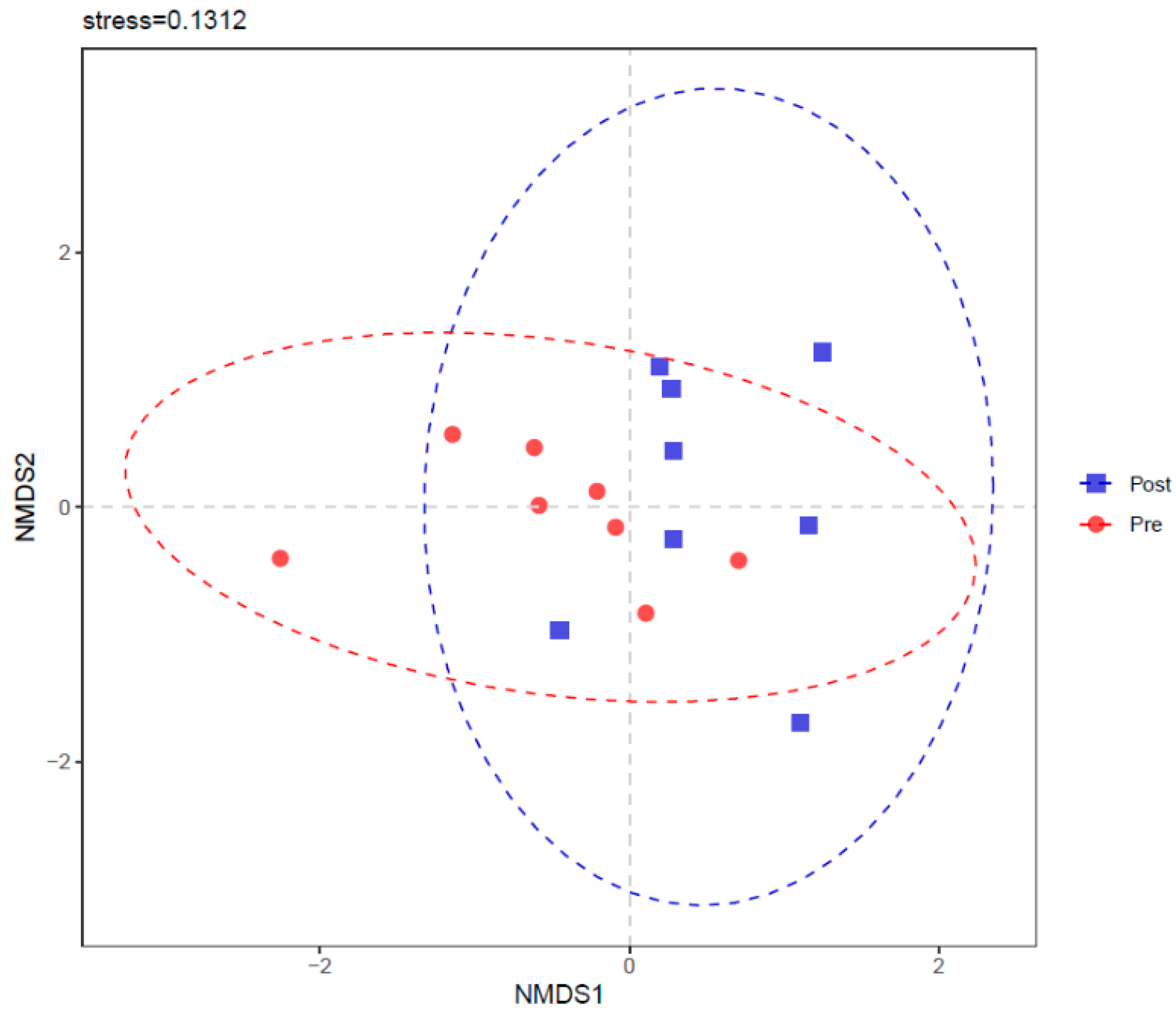
3.5. Rumen Bacterial Diversity and Community Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pitta, D.W.; Indugu, N.; Baker, L.; Vecchiarelli, B.; Attwood, G. Symposium review: Understanding diet-microbe interactions to enhance productivity of dairy cows. J. Dairy Sci. 2018, 101, 7661–7679. [Google Scholar] [CrossRef]
- Brede, M.; Orton, T.; Pinior, B.; Roch, F.-F.; Dzieciol, M.; Zwirzitz, B.; Wagner, M.; Breves, G.; Wetzels, S.U. PacBio and Illumina MiSeq amplicon sequencing confirm full recovery of the bacterial community after subacute ruminal acidosis challenge in the RUSITEC system. Front. Microbiol. 2020, 11, 1813. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, M.; Xue, C.; Zhu, W.; Mao, S. Characterization and comparison of the temporal dynamics of ruminal bacterial microbiota colonizing rice straw and alfalfa hay within ruminants. J. Dairy Sci. 2016, 99, 9668–9681. [Google Scholar] [CrossRef] [PubMed]
- Fernando, S.C.; Purvis, H.T.; Najar, F.Z.; Sukharnikov, L.O.; Krehbiel, C.R.; Nagaraja, T.G.; Roe, B.A.; DeSilva, U. Rumen microbial population dynamics during adaptation to a high-grain diet. Appl. Environ. Microbiol. 2010, 76, 7482–7490. [Google Scholar] [CrossRef] [PubMed]
- Bickhart, D.M.; Weimer, P.J. Symposium review: Host–rumen microbe interactions may be leveraged to improve the productivity of dairy cows. J. Dairy Sci. 2018, 101, 7680–7689. [Google Scholar] [CrossRef] [PubMed]
- Uyeno, Y.; Sekiguchi, Y.; Tajima, K.; Takenaka, A.; Kurihara, M.; Kamagata, Y. An rRNA-based analysis for evaluating the effect of heat stress on the rumen microbial composition of Holstein heifers. Anaerobe 2010, 16, 27–33. [Google Scholar] [CrossRef]
- Huws, S.A.; Edwards, J.E.; Lin, W.; Rubino, F.; Alston, M.; Swarbreck, D.; Caim, S.; Stevens, P.R.; Pachebat, J.; Won, M.Y.; et al. Microbiomes attached to fresh perennial ryegrass are temporally resilient and adapt to changing ecological niches. Microbiome 2021, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Ines, G.; Takumi, S.; Makoto, M. Mining of luxS genes from rumen microbial consortia by metagenomic and metatranscriptomic approaches. Anim. Sci. J. 2015, 87, 666–673. [Google Scholar]
- Vendeville, A.; Winzer, K.; Heurlier, K.; Tang, C.; Hardie, K. Making ‘sense’ of metabolism: Autoinducer-2, LUXS and pathogenic bacteria. Nat. Rev. Microbiol. 2005, 3, 383–396. [Google Scholar] [CrossRef]
- Ran, T.; Zhou, C.S.; Xu, L.W.; Geng, M.M.; Tan, Z.L.; Tang, S.X.; Wang, M.; Han, X.F.; Kang, J.H. Initial detection of the quorum sensing autoinducer activity in the rumen of goats in vivo and in vitro. J. Integr. Agric. 2016, 15, 2343–2352. [Google Scholar] [CrossRef]
- Sharifi, A.; Nayeri Fasaei, B. Selected plant essential oils inhibit biofilm formation and luxS- and pfs-mediated quorum sensing by Escherichia coli O157:H7. Lett. Appl. Microbiol. 2022, 74, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.A.; Ahmer, B.M.M. Detection of acyl-homoserine lactones by Escherichia and Salmonella. Curr. Opin. Microbiol. 2011, 14, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Lenz, D.H.; Mok, K.C.; Lilley, B.N.; Kulkarni, R.V.; Wingreen, N.S.; Bassler, B.L. The small RNA chaperone Hfq and multiple small RNAs control quorum sensing in Vibrio harveyi and Vibrio cholerae. Cell 2004, 118, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Erickson, D.L.; Nsereko, V.L.; Morgavi, D.P.; Selinger, L.B.; Rode, L.M.; Beauchemin, K.A. Evidence of quorum sensing in the rumen ecosystem: Detection of N-acyl homoserine lactone autoinducers in ruminal contents. Can. J. Microbiol. 2002, 48, 374–378. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, M.; Hardwidge, P.R.; Cui, H.; Zhu, G. Isolation and characterization of N-acyl homoserine lactone-producing bacteria from cattle rumen and swine intestines. Front. Cell. Infect. Microbiol. 2018, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, V. SdiA sensing of acyl-homoserine lactones by enterohemorrhagic E. coli (EHEC) serotype O157:H7 in the bovine rumen. Gut Microbes 2010, 1, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Won, M.Y.; Oyama, L.B.; Courtney, S.J.; Creevey, C.J.; Huws, S.A. Can rumen bacteria communicate to each other? Microbiome 2020, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.J.; Liu, H.J.; Weng, C.H.; Lai, C.F.; Ai, L.Y.; Liu, Y.C.; Zhu, H. The response of Serratia marcescens JG to environmental changes by quorum sensing system. Arch. Microbiol. 2016, 198, 585–590. [Google Scholar] [CrossRef] [PubMed]
- National Academies of Sciences, Engineering, and Medicine (NASEM). Nutrient Requirements of Beef Cattle, 8th ed.; The National Academies Press: Washington, DC, USA, 2016; pp. 397–399. [Google Scholar] [CrossRef]
- Clemmons, B.A.; Martino, C.; Schneider, L.G.; Lefler, J.; Embree, M.M.; Myer, P.R. Temporal stability of the ruminal bacterial communities in beef steers. Sci. Rep. 2019, 9, 9522. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Ponce, C.H.; Pulikanti, R. Adaptation of beef cattle to high-concentrate diets: Performance and ruminal metabolism. J. Anim. Sci. 2006, 84, E25–E33. [Google Scholar] [CrossRef] [PubMed]
- Palmonari, A.; Stevenson, D.M.; Mertens, D.R.; Cruywagen, C.W.; Weimer, P.J. pH dynamics and bacterial community composition in the rumen of lactating dairy cows. J. Dairy Sci. 2010, 93, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Paz, H.A.; Anderson, C.L.; Muller, M.J.; Kononoff, P.J.; Fernando, S.C. Rumen bacterial community composition in Holstein and Jersey cows is different under same dietary condition and is not affected by sampling method. Front. Microbiol. 2016, 7, 1206. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Denman, S.E.; Morrison, M.; Yu, Z.; McSweeney, C.S. An efficient RNA extraction method for estimating gut microbial diversity by polymerase chain reaction. Curr. Microbiol. 2009, 58, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Tian, J.; Zhang, Y.; Wu, R.; He, Y. Dissecting signal molecule AI-2 mediated biofilm formation and environmental tolerance in Lactobacillus plantarum. J. Biosci. Bioeng. 2020, 131, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Wattanavanitchakorn, S.; Prakitchaiwattana, C.; Thamyongkit, P. Rapid and simple colorimetric method for the quantification of AI-2 produced from Salmonella Typhimurium. J. Microbiol. Methods 2014, 99, 15–21. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Frølund, B.; Palmgren, R.; Keiding, K.; Nielsen, P.H. Extraction of extracellular polymers from activated sludge using a cation exchange resin. Water Res. 1996, 30, 1749–1758. [Google Scholar] [CrossRef]
- Qiu, Q.; Wei, X.; Zhang, L.; Li, Y.; Qu, M.; Ouyang, K. Effect of dietary inclusion of tea residue and tea leaves on ruminal fermentation characteristics and methane production. Anim. Biotechnol. 2021; in press. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, Q.; Li, W.; Wang, Y.; Zhang, F.; Lv, L.; Li, S.; Yang, H. High-gossypol whole cottonseed exhibited mediocre rumen degradability and less microbial fermentation efficiency than cottonseed hull and cottonseed meal with an in vitro gas production technique. Fermentation 2022, 8, 103. [Google Scholar] [CrossRef]
- Qiu, Q.; Gao, C.; Aziz ur Rahman, M.; Cao, B.; Su, H. Digestive ability, physiological characteristics, and rumen bacterial community of Holstein finishing steers in response to three nutrient density diets as fattening phases advanced. Microorganisms 2020, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: a fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, S.S.; Kemeny, M.E. Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychol. Bull. 2004, 130, 355–391. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, J.M. Manipulating the immune system with immune globulin. N. Engl. J. Med. 1992, 326, 107–116. [Google Scholar]
- Tadahisa, F.; Hiroya, M.; Takahiro, I.; Hiroaki, S. Combined effect of salinomycin and feeding on whole body glucose kinetics in sheep fed a high-concentrate diet. Reprod. Nutr. Dev. 2006, 46, 503–514. [Google Scholar]
- Judson, G.J.; Leng, R.A. Effect of diet on glucose synthesis in sheep. Proc. Aust. Soc. Anim. Prod. 1968, 7, 354–358. [Google Scholar]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef]
- Asanuma, N.; Yoshii, T.; Hino, T. Molecular characterization and transcription of the luxS gene that encodes LuxS autoinducer 2 synthase in Streptococcus bovis. Curr. Microbiol. 2004, 49, 366–371. [Google Scholar] [CrossRef]
- Allison, D.G. The biofilm matrix. Biofouling 2003, 19, 139–150. [Google Scholar] [CrossRef]
- Tan, L.J.; Wang, J.J.; Peng, Z.Y.; Li, Y.F.; Zeng, Q.H.; Zhao, Y. Mixed-species biofilms formation, interaction and novel control strategies in the food industry. Food Science 2022, in press. [Google Scholar]
- Lee, K.W.K.; Periasamy, S.; Mukherjee, M.; Xie, C.; Kjelleberg, S.; Rice, S.A. Biofilm development and enhanced stress resistance of a model, mixed-species community biofilm. ISME J. 2014, 8, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Bove, P.; Capozzi, V.; Garofalo, C.; Rieu, A.; Spano, G.; Fiocco, D. Inactivation of the ftsH gene of Lactobacillus plantarum WCFS1: Effects on growth, stress tolerance, cell surface properties and biofilm formation. Microbiol. Res. 2012, 167, 187–193. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, B.; Grenier, D.; Yi, L. Regulatory mechanisms of the LuxS/AI-2 system and bacterial resistance. Antimicrob. Agents Chemother. 2019, 63, e01186-19. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.J.; Xu, T.W.; Xu, S.X.; Ma, L.; Han, X.P.; Wang, X.G.; Zhang, X.L.; Hu, L.Y.; Zhao, N.; Chen, Y.W.; et al. Effect of dietary concentrate to forage ratio on growth performance, rumen fermentation and bacterial diversity of Tibetan sheep under barn feeding on the Qinghai-Tibetan plateau. PeerJ 2019, 7, e7462. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, J. Production and absorption of volatile fatty acids in the rumen. Livest. Prod. Sci. 1994, 39, 61–69. [Google Scholar] [CrossRef]
- Wang, Q.; Zeng, Y.; Zeng, X.; Wang, X.; Wang, Y.; Dai, C.; Li, J.; Huang, P.; Huang, J.; Hussain, T.; et al. Effects of dietary energy levels on rumen fermentation, gastrointestinal tract histology, and bacterial community diversity in fattening male Hu lambs. Front. Microbiol. 2021, 12, 695445. [Google Scholar] [CrossRef]
- Yang, C.M.J. Response of forage fiber degradation by ruminal microorganisms to branched-chain volatile fatty acids, amino acids, and dipeptides. J. Dairy Sci. 2002, 85, 1183–1190. [Google Scholar] [CrossRef]
- Zhao, D.; Gao, P.; Xu, L.; Qu, L.; Han, Y.; Zheng, L.; Gong, X. Disproportionate responses between free-living and particle-attached bacteria during the transition to oxygen-deficient zones in the Bohai Seawater. Sci. Total Environ. 2021, 791, 148097. [Google Scholar] [CrossRef]
- Sano, H.; Ito, T.; Terashima, Y. Effect of diet forage-to-concentrate ratio on partition of dietary energy and nutrients in fed and fasted sheep. J. Appl. Anim. Res. 2004, 25, 101–108. [Google Scholar] [CrossRef][Green Version]
- Purushe, J.; Fouts, D.E.; Morrison, M.; White, B.A.; Mackie, R.I.; Coutinho, P.M.; Henrissat, B.; Nelson, K.E. Comparative genome analysis of prevotella ruminicola and prevotella bryantii: Insights into their environmental niche. Microb. Ecol. 2010, 60, 721–729. [Google Scholar] [CrossRef]
- Scott, K.P.; Martin, J.C.; Campbell, G.; Mayer, C.; Flint, H.J. Whole-genome transcription profiling reveals genes up-regulated by growth on fucose in the human gut bacterium “Roseburia inulinivorans”. J. Bacteriol. 2006, 188, 4340–4349. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Fina, L.R.; Lassman, B.A.; Bartley, E.E.; Anthony, H.D.; Sapienza, D.A.; Brent, B.E. Characterization of endotoxin from the rumen bacterium Megasphaera elsdenii. Am. J. Vet. Res. 1979, 40, 35–39. [Google Scholar]
- Mazon, G.; Campler, M.R.; Holcomb, C.; Bewley, J.M.; Costa, J.H.C. Effects of a Megasphaera elsdenii oral drench on reticulorumen pH dynamics in lactating dairy cows under subacute ruminal acidosis challenge. Anim. Feed Sci. Technol. 2020, 261, 114404. [Google Scholar] [CrossRef]
- Trabi, E.B.; Seddik, H.; Xie, F.; Lin, L.; Mao, S. Comparison of the rumen bacterial community, rumen fermentation and growth performance of fattening lambs fed low-grain, pelleted or non-pelleted high grain total mixed ration. Anim. Feed Sci. Technol. 2019, 253, 1–12. [Google Scholar] [CrossRef]
- Li, D.; Leahy, S.; Henderson, G.; Kelly, W.; Cookson, A.; Attwood, G.; Moon, C. Atypical bacterial rRNA operon structure is prevalent within the Lachnospiraceae, and use of the 16S-23S rRNA internal transcribed spacer region for the rapid identification of ruminal Butyrivibrio and Pseudobutyrivibrio strains. Ann. Microbiol. 2014, 64, 1623–1631. [Google Scholar] [CrossRef]
- Liu, K.; Xu, Q.; Wang, L.; Wang, J.; Guo, W.; Zhou, M. The impact of diet on the composition and relative abundance of rumen microbes in goat. Asian-Australas. J. Anim. Sci. 2017, 30, 531–537. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Pre-Shift Diet | Post-Shift Diet |
|---|---|---|
| Ingredients, % of dry matter (DM) | ||
| Corn | 36.20 | 20.00 |
| Soybean meal | 19.47 | 19.80 |
| Wheat bran | 13.24 | 3.49 |
| Wheat straw | 25.09 | 50.71 |
| Calcium bicarbonate | 0.50 | 0.50 |
| Calcium hydrophosphate | 0.50 | 0.50 |
| Limestone | 0.50 | 0.50 |
| Salt | 0.50 | 0.50 |
| Premix 1 | 4.00 | 4.00 |
| Total | 100 | 100 |
| Nutritional composition, g/kg of DM | ||
| Metabolic energy (ME), MJ/kg | 10.97 | 9.59 |
| Crude protein (CP) | 161.4 | 141.2 |
| ME to CP ratio, MJ/g | 0.068 | 0.068 |
| Neutral detergent fiber (NDF) | 308.6 | 428.0 |
| Acid detergent fiber (ADF) | 164.8 | 266.4 |
| Accession Number | Gene Name | Primer Sequence (5-3) | Product Size (bp) |
|---|---|---|---|
| SEW05678.1 | 16S rRNA-F | AGAGCCTGAACCAGCCAAGTAG | 128 |
| 16S rRNA-R | GAATTAGCCGGTCCTTATTCATACA | ||
| luxS-F | GGATGATGTAGTGTATGTCGGTCC | 185 | |
| luxS-R | GGAGGTCGTGGAGCAGATAGTT | ||
| SHK82531.1 | 16S rRNA-F | TGCGTCTGATTAGGTAGTAGGCG | 112 |
| 16S rRNA-R | CGTAGGAGTTTGGACCGTGTCT | ||
| ftsH-F | AGATGTATGAGAAGGGTGGTGAGT | 146 | |
| ftsH--R | TCCCTTGGGTATCTTACCTCCC |
| Item 1 | Pre | Post | SEM 2 | p-Value |
|---|---|---|---|---|
| pH value | 6.94 | 6.95 | 0.07 | 0.821 |
| Molar concentration (mM) | ||||
| Acetate | 11.06 | 10.68 | 1.137 | 0.743 |
| Propionate | 5.89 | 4.94 | 0.673 | 0.180 |
| Isobutyrate | 0.11 | 0.10 | 0.022 | 0.448 |
| Butyrate | 1.26 | 1.52 | 0.171 | 0.153 |
| Isovalerate | 0.18 | 0.16 | 0.019 | 0.327 |
| Valerate | 0.56 | 0.47 | 0.044 | 0.049 |
| Total volatile fatty acids | 19.07 | 17.86 | 1.945 | 0.546 |
| Branched-chain volatile fatty acids | 0.86 | 0.72 | 0.073 | 0.092 |
| Acetate to propionate ratio | 1.98 | 2.28 | 0.114 | 0.017 |
| NGR | 2.29 | 2.73 | 0.117 | 0.002 |
| Fermentation efficiency | 0.78 | 0.77 | 0.005 | 0.022 |
| Molar proportion (mol/100 mol) | ||||
| Acetate | 58.62 | 60.17 | 1.137 | 0.194 |
| Propionate | 30.30 | 27.21 | 1.041 | 0.010 |
| Isobutyrate | 0.64 | 0.59 | 0.102 | 0.678 |
| Butyrate | 6.51 | 8.40 | 0.673 | 0.014 |
| Isovalerate | 0.99 | 0.94 | 0.056 | 0.406 |
| Valerate | 2.95 | 2.68 | 0.111 | 0.032 |
| Branched-chain volatile fatty acids | 4.57 | 4.22 | 0.183 | 0.072 |
| Item | Pre | Post | SEM | p-Value |
|---|---|---|---|---|
| Chao 1 | 592.76 | 648.48 | 10.47 | 0.001 |
| Observed species | 506.13 | 560.74 | 9.41 | 0.001 |
| PD whole tree 1 | 30.63 | 33.00 | 0.44 | 0.001 |
| Shannon index | 5.44 | 5.61 | 0.17 | 0.383 |
| Simpson index | 0.94 | 0.94 | 0.02 | 0.807 |
| Item | Pre | Post | SEM | p-Value |
|---|---|---|---|---|
| Bacteroidetes | 51.46 | 46.40 | 2.89 | 0.124 |
| Firmicutes | 27.21 | 25.50 | 4.09 | 0.689 |
| Proteobacteria | 18.56 | 25.19 | 5.38 | 0.257 |
| Cyanobacteria | 0.74 | 1.07 | 0.26 | 0.237 |
| Actinobacteriota | 1.00 | 0.62 | 0.17 | 0.059 |
| Fibrobacterota | 0.53 | 0.64 | 0.46 | 0.822 |
| Desulfobacterota | 0.25 | 0.27 | 0.06 | 0.823 |
| Spirochaetota | 0.12 | 0.19 | 0.04 | 0.121 |
| Item | Pre | Post | SEM | p-Value |
|---|---|---|---|---|
| Prevotella | 49.72 | 42.77 | 2.79 | 0.042 |
| Succinivibrio | 15.50 | 21.95 | 5.15 | 0.251 |
| Roseburia | 1.42 | 5.64 | 1.75 | 0.047 |
| Succinivibrionaceae UCG-001 | 2.78 | 2.70 | 0.81 | 0.922 |
| Selenomonas | 3.44 | 1.94 | 1.02 | 0.185 |
| Erysipelotrichaceae UCG-002 | 4.00 | 0.55 | 2.90 | 0.274 |
| Megasphaera | 2.53 | 0.72 | 0.34 | 0.001 |
| Syntrophococcus | 1.22 | 1.28 | 0.17 | 0.719 |
| Acetitomaculum | 1.85 | 0.52 | 0.63 | 0.072 |
| Dialister | 1.46 | 0.91 | 0.16 | 0.011 |
| Oribacterium | 1.04 | 1.26 | 0.34 | 0.552 |
| Prevotellaceae UCG-001 | 0.20 | 1.41 | 0.31 | 0.006 |
| Lachnospiraceae NK3A20 group | 0.94 | 0.58 | 0.37 | 0.348 |
| Acidaminococcus | 0.87 | 0.58 | 0.17 | 0.126 |
| Ruminococcus | 0.86 | 0.43 | 0.25 | 0.132 |
| Succiniclasticum | 0.49 | 0.80 | 0.18 | 0.123 |
| Fibrobacter | 0.53 | 0.64 | 0.46 | 0.822 |
| Olsenella | 0.69 | 0.47 | 0.12 | 0.119 |
| Lachnospira | 0.26 | 0.77 | 0.13 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.; Long, T.; Li, Y.; Ouyang, K.; Qiu, Q. Diet Shift May Trigger LuxS/AI-2 Quorum Sensing in Rumen Bacteria. Bioengineering 2022, 9, 379. https://doi.org/10.3390/bioengineering9080379
Wei X, Long T, Li Y, Ouyang K, Qiu Q. Diet Shift May Trigger LuxS/AI-2 Quorum Sensing in Rumen Bacteria. Bioengineering. 2022; 9(8):379. https://doi.org/10.3390/bioengineering9080379
Chicago/Turabian StyleWei, Xiao, Tanghui Long, Yanjiao Li, Kehui Ouyang, and Qinghua Qiu. 2022. "Diet Shift May Trigger LuxS/AI-2 Quorum Sensing in Rumen Bacteria" Bioengineering 9, no. 8: 379. https://doi.org/10.3390/bioengineering9080379
APA StyleWei, X., Long, T., Li, Y., Ouyang, K., & Qiu, Q. (2022). Diet Shift May Trigger LuxS/AI-2 Quorum Sensing in Rumen Bacteria. Bioengineering, 9(8), 379. https://doi.org/10.3390/bioengineering9080379