Depletion of Boric Acid and Cobalt from Cultivation Media: Impact on Recombinant Protein Production with Komagataella phaffii

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Strains and Proteins
2.3. Leakage of Elements from Cultivation Vessels
2.4. Preculture and Cultivation Media
2.4.1. Precultures
2.4.2. Bioreactor Cultivation Media
2.5. Bioreactor Cultivations
2.5.1. Batch, Fed-Batch and Induction PHASE
2.5.2. Description of Single Cultivations
2.5.3. Cultivation Sample Preparation
2.6. Data Analysis
2.7. High Pressure Liquid Chromatography
2.8. Flow Cytometry
2.9. Protein Purification
2.9.1. GalOx
2.9.2. HRP
2.9.3. FC
2.10. Protein Concentration and Activity
2.10.1. Protein Concentration
2.10.2. Enzyme Activity
2.11. Enzyme Kinetics and Thermal Stability
2.12. ICP-OES Measurements
2.13. N-Glycosylation Profiling
3. Results
3.1. Leakage of Elements, Elemental Media Composition and Elemental Consumption
3.1.1. Elemental Leakage from Vessels
3.1.2. Elemental Media Composition
3.1.3. Elemental Consumption Analysis
3.2. Impact of Co and BA Depletion on Recombinant Protein Production in Glass Bioreactors
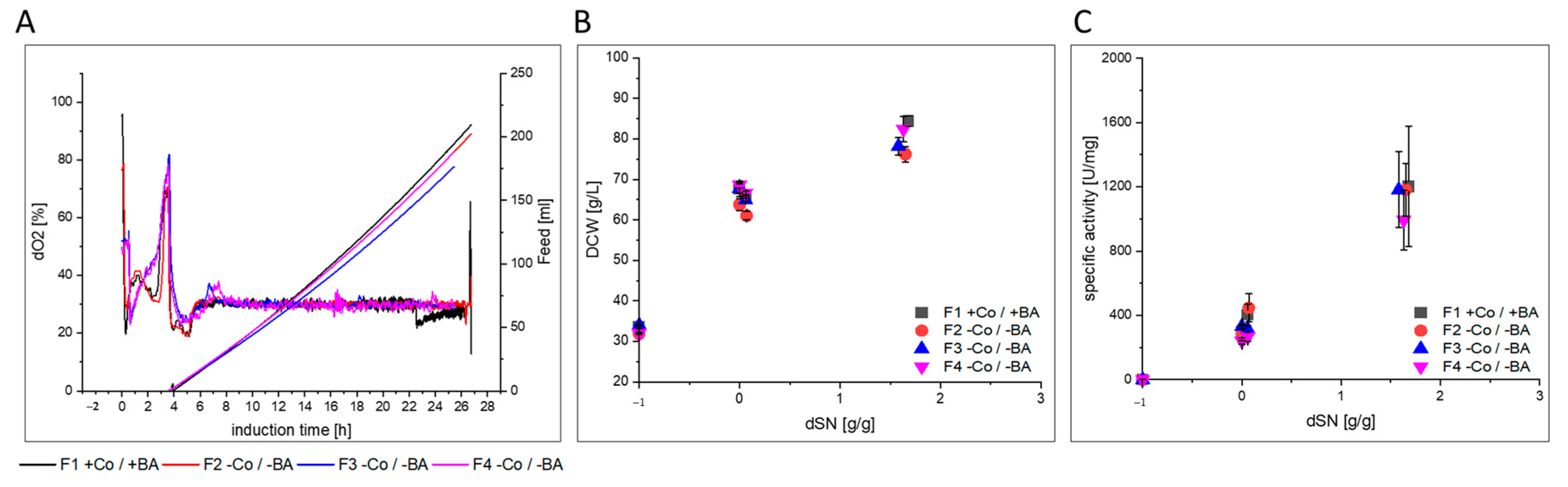
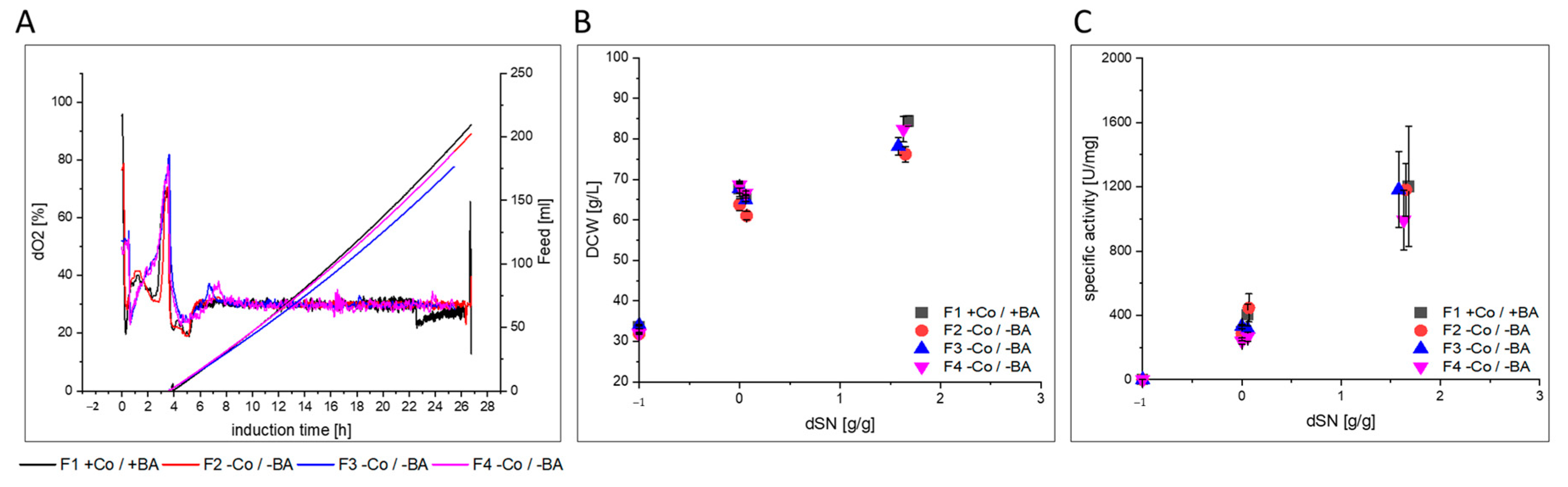
3.2.1. GalOx
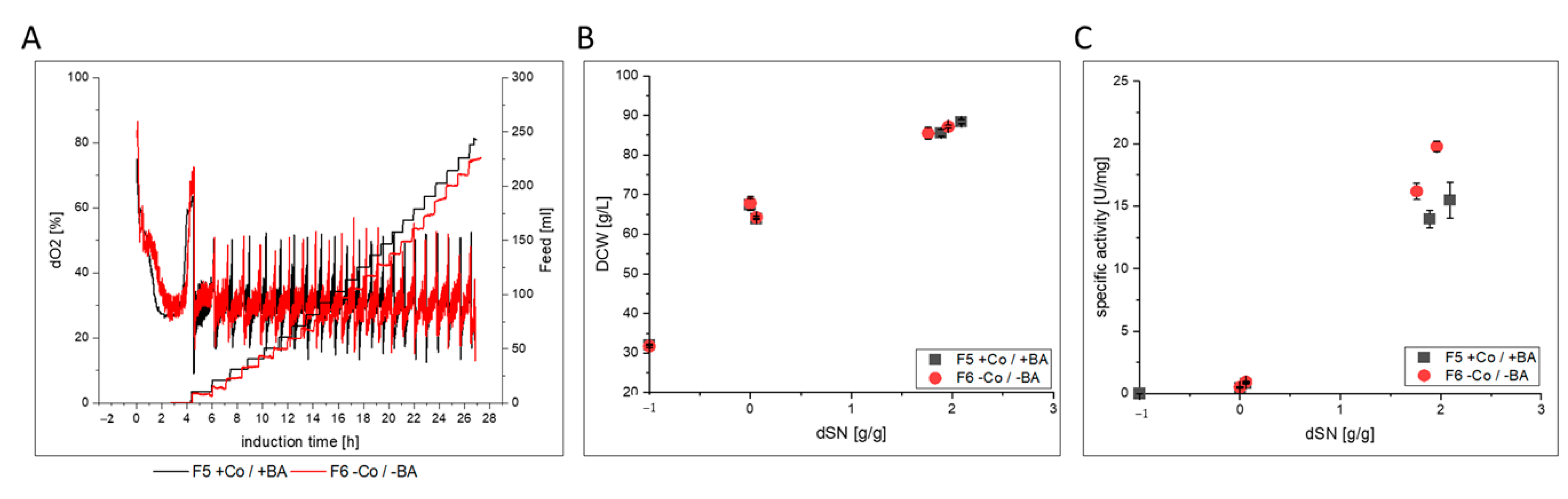
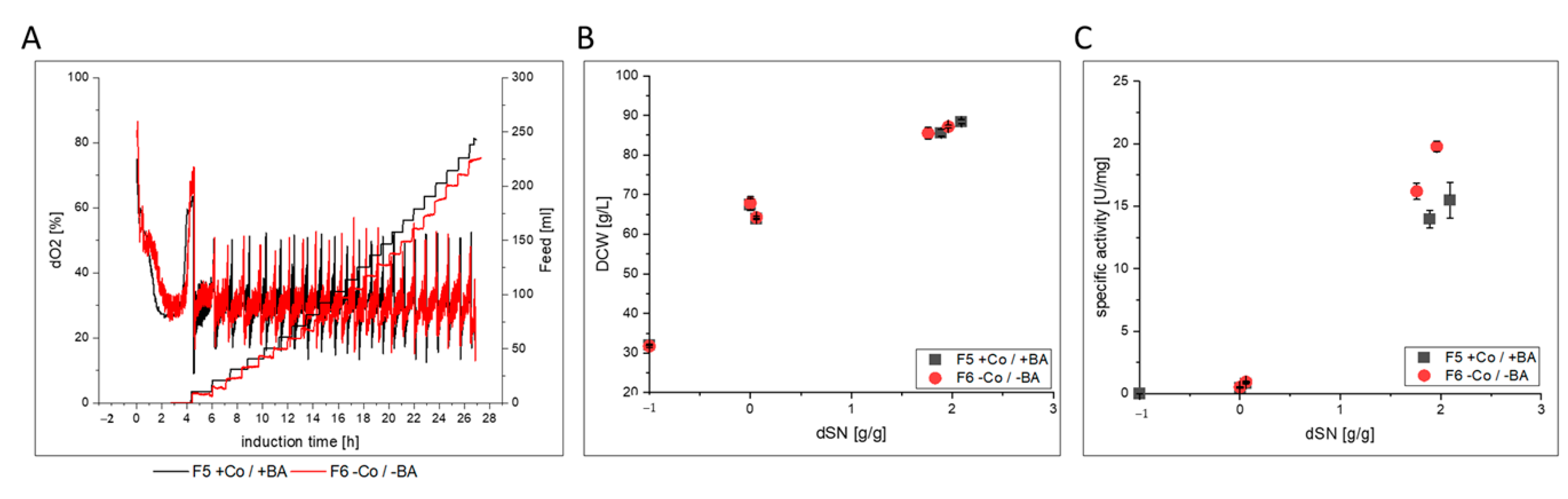
3.2.2. HRP
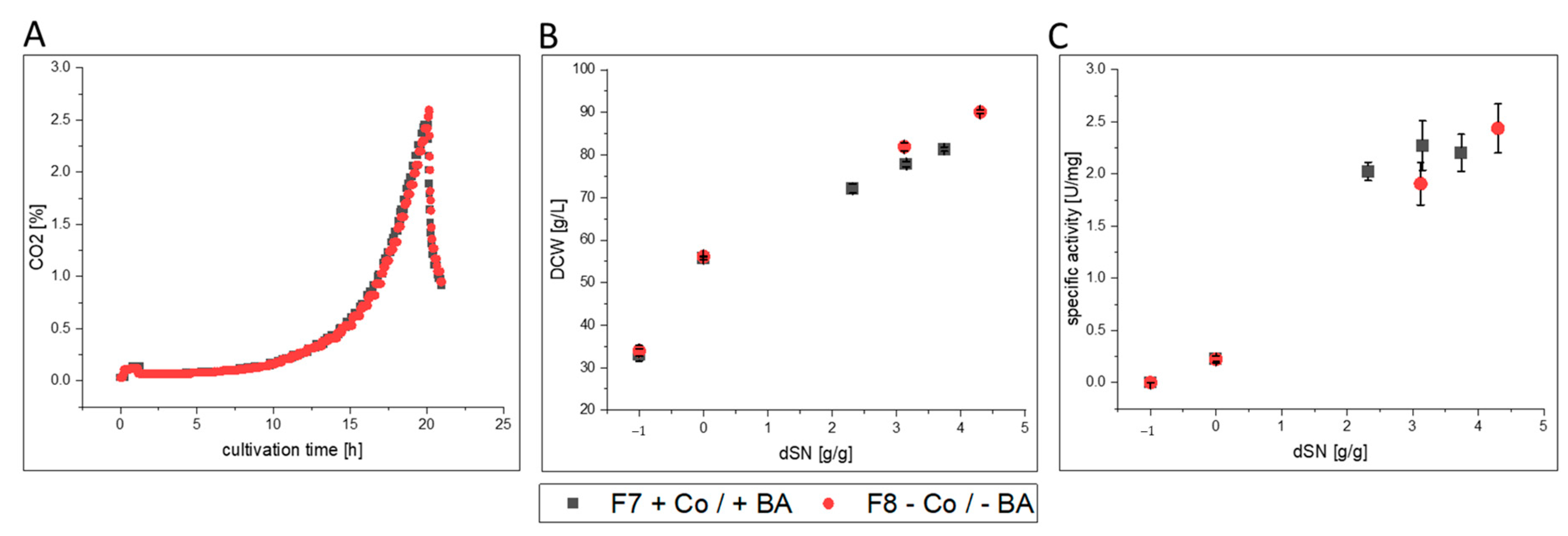
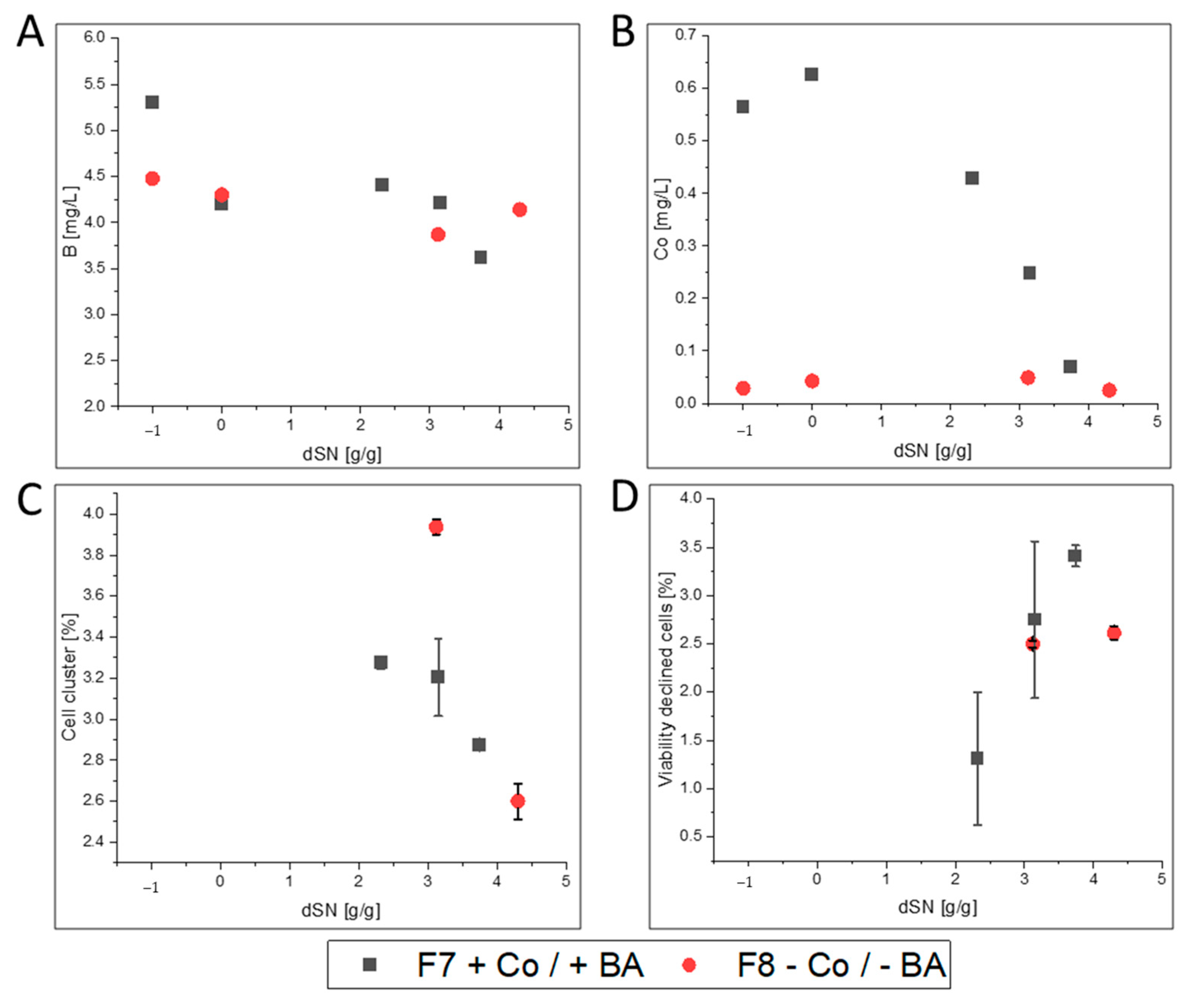
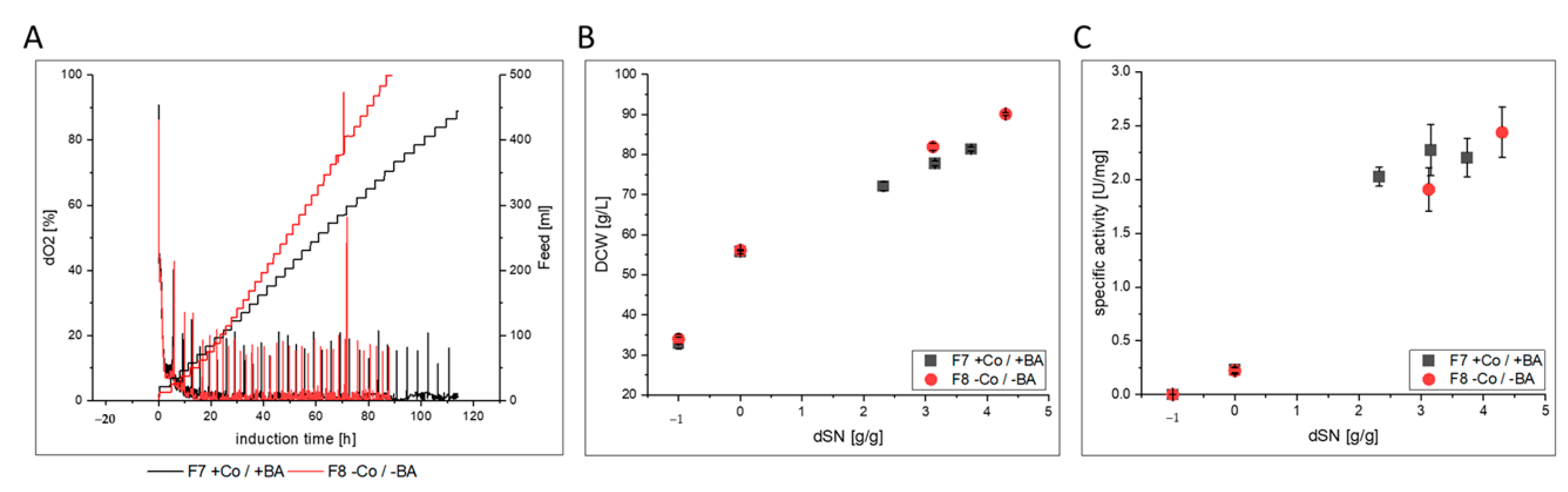
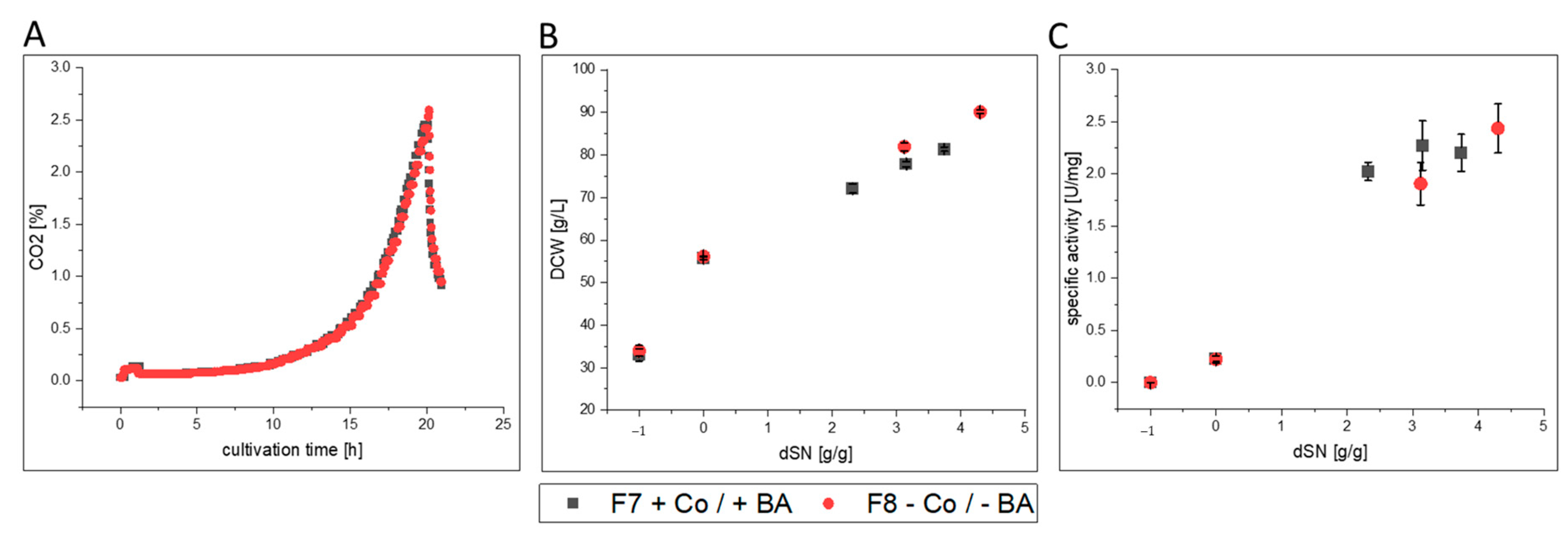
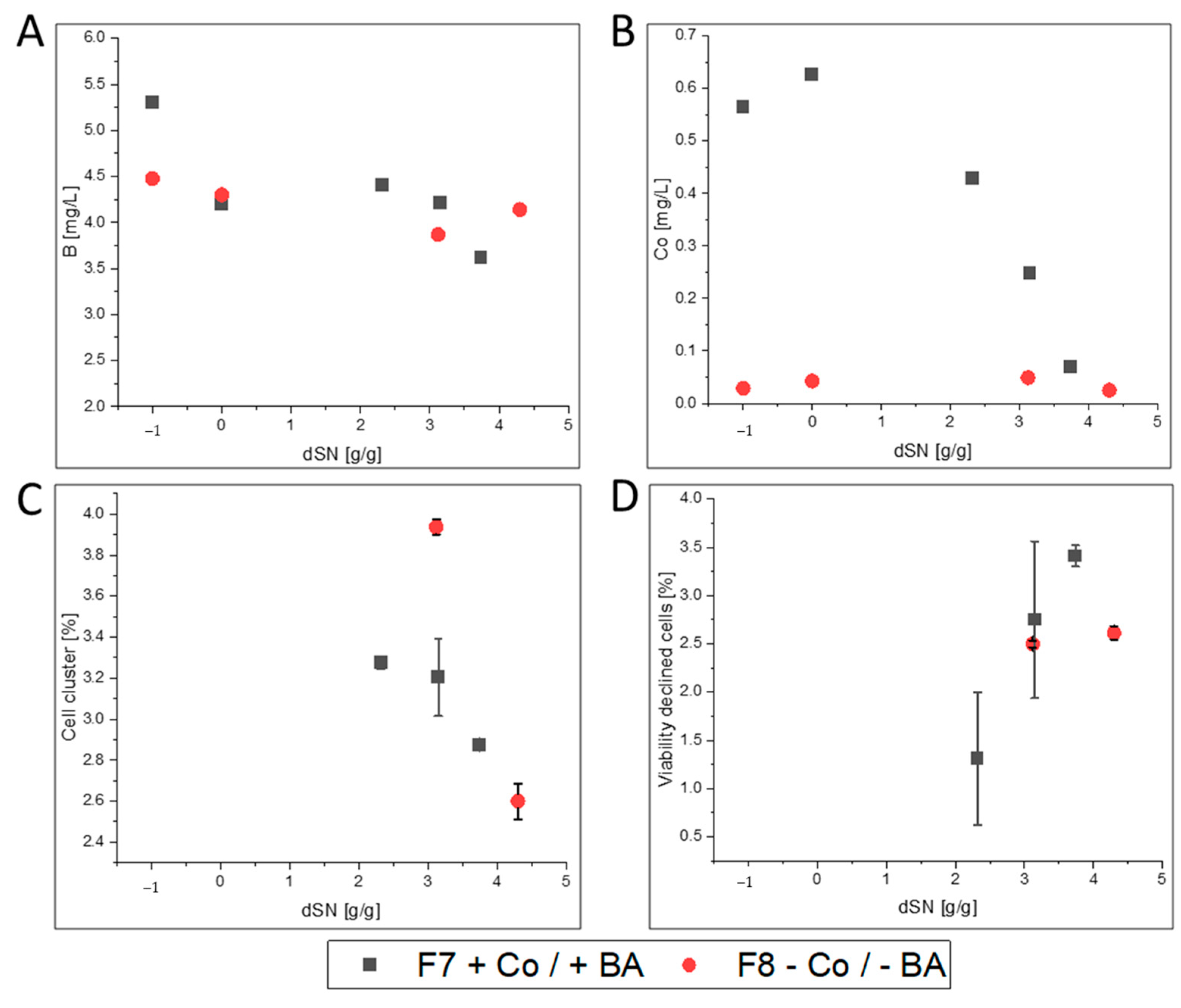
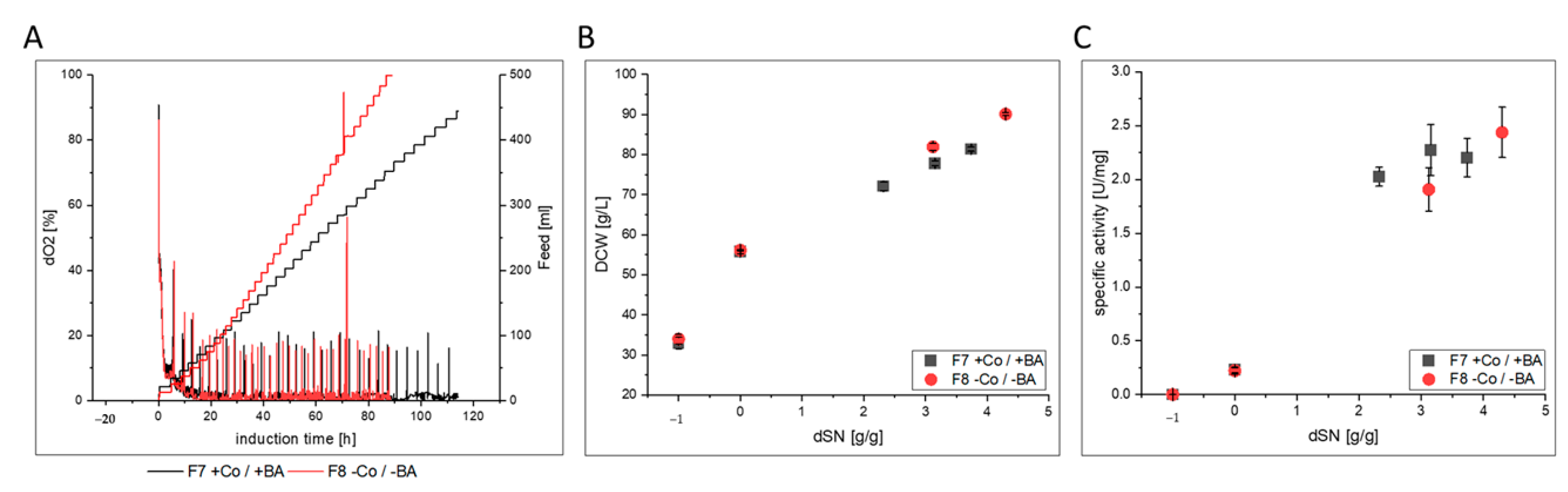
3.2.3. FC
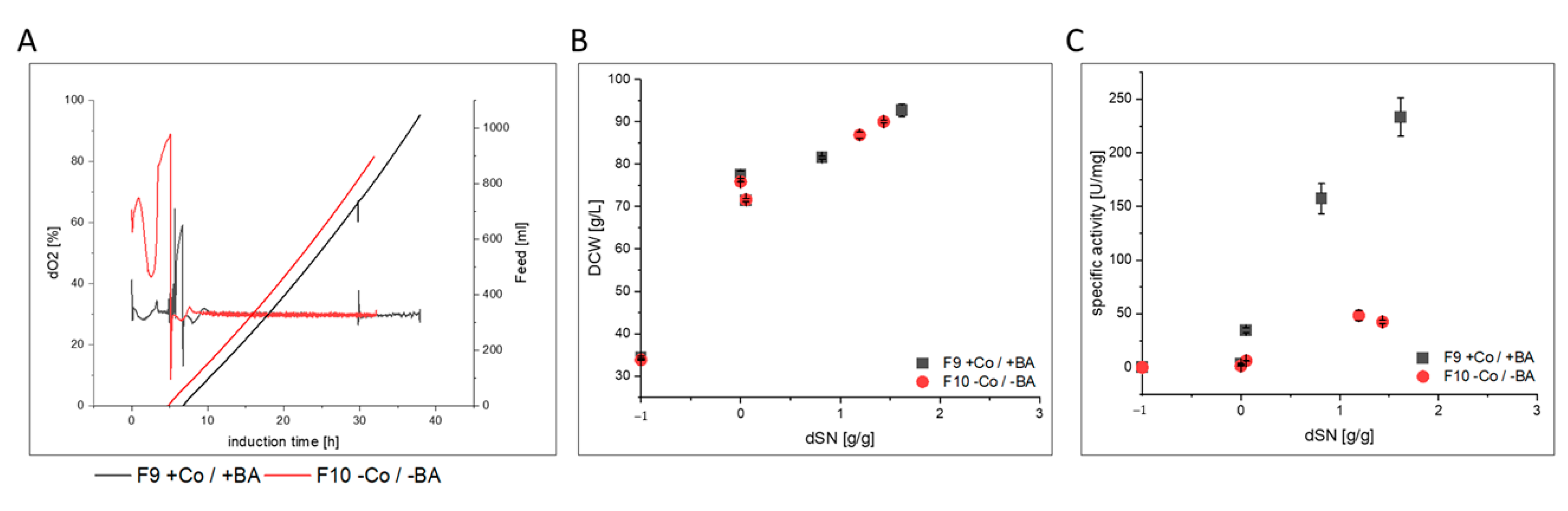
3.3. Impact of Co and BA Depletion on HRP Production in a Stainless Steel Bioreactors
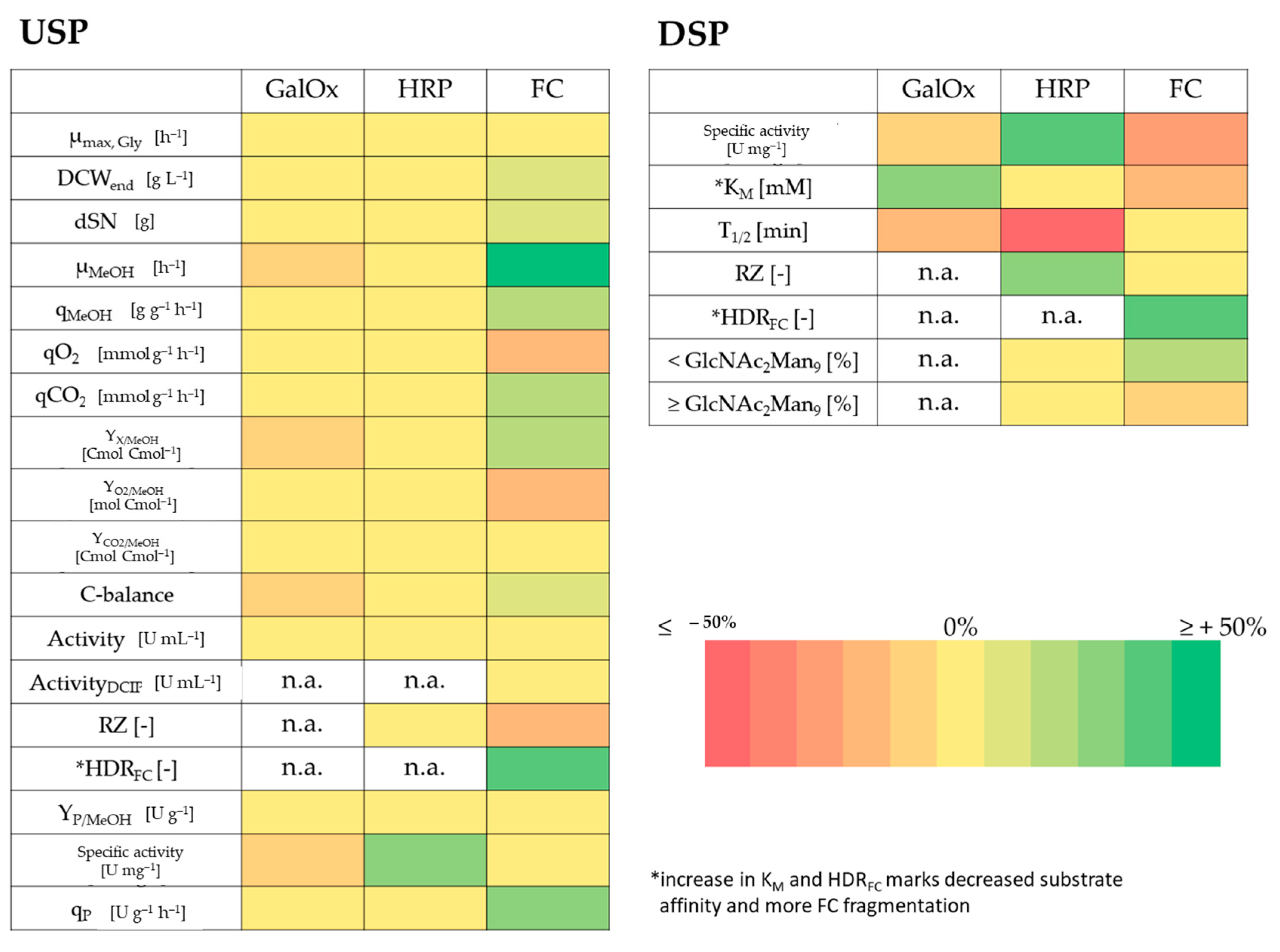
4. Discussion
- GalOx producing cells in F2–F4 showed a slightly lower µMeOH and purified GalOx had a decreased substrate affinity.
- HRP producing cells in F6 showed a slightly decreased host cell protein secretion that led to increased specific activity in the broth. After purification, the enzyme kinetics and N-glycosylation patterns were not affected, but the thermal stability was 3-fold reduced. However, when cells were cultivated under exponential rather than pulsed methanol feeding in a stainless steel bioreactor, HRP-producing cells in F10 had an extensively decreased productivity. N-glycosylation was affected and surprisingly, HRP from F10 showed a 3-fold decrease in thermal stability, similar to HRP from F6.
- FC-producing cells in F8 showed ethanol production, faster growth and an increased productivity. The purified enzyme had increased substrate affinity and decreased glycosylation.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations of Variables
References
- Vieira Gomes, A.M.; Souza Carmo, T.; Silva Carvalho, L.; Mendonça Bahia, F.; Parachin, N.S. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spohner, S.C.; Schaum, V.; Quitmann, H.; Czermak, P. Kluyveromyces lactis: An emerging tool in biotechnology. J. Biotechnol. 2016, 222, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Bankar, A.V.; Kumar, A.R.; Zinjarde, S.S. Environmental and industrial applications of Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2009, 84, 847–865. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J. Production of biopharmaceutical proteins by yeast: Advances through metabolic engineering. Bioengineered 2013, 4, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Anumanthan, A.; Gao, X.-G.; Ilangovan, K.; Suzara, V.V.; Düzgüneş, N.; Renugopalakrishnan, V. Expression of Recombinant Proteins in Pichia Pastoris. Appl. Biochem. Biotechnol. 2007, 142, 105–124. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, Y.; Miao, H.; Tang, X.; Xu, B.; Wu, Q.; Mu, Y.; Huang, Z. Transcriptomic Analysis of Pichia pastoris (Komagataella phaffii) GS115 During Heterologous Protein Production Using a High-Cell-Density Fed-Batch Cultivation Strategy. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Kumar, R.; Kumar, P. Yeast-based vaccines: New perspective in vaccine development and application. Fems Yeast Res. 2019, 19. [Google Scholar] [CrossRef] [Green Version]
- Bretthauer, R.K.; Castellino, F.J. Glycosylation of Pichia pastoris-derived proteins. Biotechnol. Appl. Biochem. 1999, 30, 193–200. [Google Scholar]
- Looser, V.; Bruhlmann, B.; Bumbak, F.; Stenger, C.; Costa, M.; Camattari, A.; Fotiadis, D.; Kovar, K. Cultivation strategies to enhance productivity of Pichia pastoris: A review. Biotechnol. Adv. 2015, 33, 1177–1193. [Google Scholar] [CrossRef] [Green Version]
- Laukens, B.; Visscher, C.D.; Callewaert, N. Engineering yeast for producing human glycoproteins: Where are we now? Future Microbiol. 2015, 10, 21–34. [Google Scholar] [CrossRef]
- EuropeanChemicalsAgency. Understanding REACH. Available online: https://echa.europa.eu/regulations/reach/understanding-reach (accessed on 14 September 2020).
- Williams, E.S.; Panko, J.; Paustenbach, D.J. The European Union’s REACH regulation: A review of its history and requirements. Crit. Rev. Toxicol. 2009, 39, 553–575. [Google Scholar] [CrossRef] [PubMed]
- EuropeanChemicalsAgency. List of Restricted Substances. Available online: https://echa.europa.eu/de/substances-restricted-under-reach (accessed on 16 November 2020).
- Hitzman, D.O.; Wegner, E.H. Methanol Foam Fermentation to Single Cell Protein by Microorganisms. U.S. Patent 3,982,998; filed 6 December 1974, and issued 28 September 1976,
- Stratton, J.; Chiruvolu, V.; Meagher, M.M. High Cell-Density Fermentation. In Pichia Protocols; Higgins, D.R., Cregg, J.M., Eds.; Humana Press: Totowa, NJ, USA, 1998; pp. 107–120. [Google Scholar] [CrossRef]
- Invitrogen-Corporation. Pichia Fermentation Process Guidelines. Version B. Available online: http://tools.thermofisher.com/content/sfs/manuals/pichiaferm_prot.pdf (accessed on 14 September 2020).
- Min, C.-K.; Lee, J.-W.; Chung, K.-H.; Park, H.-W. Control of specific growth rate to enhance the production of a novel disintegrin, saxatilin, in recombinant Pichia pastoris. J. Biosci. Bioeng. 2010, 110, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Bhatacharya, P.; Pandey, G.; Mukherjee, K.J. Production and purification of recombinant human granulocyte–macrophage colony stimulating factor (GM-CSF) from high cell density cultures of Pichia pastoris. Bioprocess Biosyst. Eng. 2007, 30, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Hang, H.-F.; Chen, W.; Guo, M.-J.; Chu, J.; Zhuang, Y.-P.; Zhang, S. A simple unstructured model-based control for efficient expression of recombinant porcine insulin precursor by Pichia pastoris. Korean J. Chem. Eng. 2008, 25, 1065–1069. [Google Scholar] [CrossRef]
- Yamawaki, S.; Matsumoto, T.; Ohnishi, Y.; Kumada, Y.; Shiomi, N.; Katsuda, T.; Lee, E.K.; Katoh, S. Production of Single-Chain Variable Fragment Antibody (scFv) in Fed-Batch and Continuous Culture of Pichia pastoris by Two Different Methanol Feeding Methods. J. Biosci. Bioeng. 2007, 104, 403–407. [Google Scholar] [CrossRef]
- Paulova, L.; Hyka, P.; Branska, B.; Melzoch, K.; Kovar, K. Use of a mixture of glucose and methanol as substrates for the production of recombinant trypsinogen in continuous cultures with Pichia pastoris Mut+. J. Biotechnol. 2012, 157, 180–188. [Google Scholar] [CrossRef]
- Dietzsch, C.; Spadiut, O.; Herwig, C. A fast approach to determine a fed batch feeding profile for recombinant Pichia pastoris strains. Microb. Cell Factories 2011, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Krainer, F.W.; Capone, S.; Jäger, M.; Vogl, T.; Gerstmann, M.; Glieder, A.; Herwig, C.; Spadiut, O. Optimizing cofactor availability for the production of recombinant heme peroxidase in Pichia pastoris. Microb. Cell Factories 2015, 14, 4. [Google Scholar] [CrossRef] [Green Version]
- Krainer, F.W.; Gmeiner, C.; Neutsch, L.; Windwarder, M.; Pletzenauer, R.; Herwig, C.; Altmann, F.; Glieder, A.; Spadiut, O. Knockout of an endogenous mannosyltransferase increases the homogeneity of glycoproteins produced in Pichia pastoris. Sci. Rep. 2013, 3, 3279. [Google Scholar] [CrossRef] [Green Version]
- Maurer, M.; Kühleitner, M.; Gasser, B.; Mattanovich, D. Versatile modeling and optimization of fed batch processes for the production of secreted heterologous proteins with Pichia pastoris. Microb. Cell Factories 2006, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Matthews, C.B.; Kuo, A.; Love, K.R.; Love, J.C. Development of a general defined medium for Pichia pastoris. Biotechnol. Bioeng. 2018, 115, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Prielhofer, R.; Maurer, M.; Klein, J.; Wenger, J.; Kiziak, C.; Gasser, B.; Mattanovich, D. Induction without methanol: Novel regulated promoters enable high-level expression in Pichia pastoris. Microb. Cell Factories 2013, 12, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laroche, Y.; Storme, V.; De Meutter, J.; Messens, J.; Lauwereys, M. High-level secretion and very efficient isotopic labeling of tick anticoagulant peptide (TAP) expressed in the methylotrophic yeast, Pichia pastoris. Bio/Technol. 1994, 12, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- EuropeanChemicalsAgency. Boric Acid. Available online: https://echa.europa.eu/substance-information/-/substanceinfo/100.030.114 (accessed on 14 September 2020).
- EuropeanChemicalsAgency. Cobalt. Available online: https://echa.europa.eu/substance-information/-/substanceinfo/100.028.325 (accessed on 14 September 2020).
- EuropeanChemicalsAgency. Cobalt Dichloride. Available online: https://echa.europa.eu/substance-information/-/substanceinfo/100.028.718 (accessed on 14 September 2020).
- EuropeanChemicalsAgency. Authorisation. Available online: https://echa.europa.eu/substances-of-very-high-concern-identification-explained (accessed on 17 November 2020).
- Uluisik, I.; Kaya, A.; Unlu, E.S.; Avsar, K.; Karakaya, H.C.; Yalcin, T.; Koc, A. Genome-wide identification of genes that play a role in boron stress response in yeast. Genomics 2011, 97, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, F.H.; Eckhert, C.D. Boron. Adv. Nutr. 2020, 11, 461–462. [Google Scholar] [CrossRef]
- O’Neill, M.A.; Ishii, T.; Albersheim, P.; Darvill, A.G. Rhamnogalacturonan II: Structure and Function of a Borate Cross-Linked Cell Wall Pectic Polysaccharide. Annu. Rev. Plant Biol. 2004, 55, 109–139. [Google Scholar] [CrossRef] [Green Version]
- Rowe, R.I.; Eckhert, C.D. Boron is required for zebrafish embryogenesis. J. Exp. Biol. 1999, 202, 1649–1654. [Google Scholar]
- Fort, D.J.; Propst, T.L.; Stover, E.L.; Strong, P.L.; Murray, F.J. Adverse reproductive and developmental effects in Xenopus from insufficient boron. Biol. Trace Elem. Res. 1998, 66, 237–259. [Google Scholar] [CrossRef]
- Lanoue, L.; Taubeneck, M.W.; Muniz, J.; Hanna, L.A.; Strong, P.L.; Murray, F.J.; Nielsen, F.H.; Hunt, C.D.; Keen, C.L. Assessing the effects of low boron diets on embryonic and fetal development in rodents using in vitro and in vivo model systems. Biol. Trace Elem. Res. 1998, 66, 271–298. [Google Scholar] [CrossRef]
- Nozawa, A.; Takano, J.; Kobayashi, M.; Von Wirén, N.; Fujiwara, T. Roles of BOR1, DUR3, and FPS1 in boron transport and tolerance in Saccharomyces cerevisiae. Fems Microbiol. Lett. 2006, 262, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Takano, J.; Kobayashi, M.; Noda, Y.; Fujiwara, T. Saccharomyces cerevisiae Bor1p is a boron exporter and a key determinant of boron tolerance. Fems Microbiol. Lett. 2007, 267, 230–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, A.; Karakaya, H.C.; Fomenko, D.E.; Gladyshev, V.N.; Koc, A. Identification of a novel system for boron transport: Atr1 is a main boron exporter in yeast. Mol. Cell Biol. 2009, 29, 3665–3674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uluisik, I.; Karakaya, H.C.; Koc, A. The importance of boron in biological systems. J. Trace Elem. Med. Biol. 2018, 45, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.; Rowe, R.I.; Soch, N.; Eckhert, C.D. Boron Stimulates Yeast (Saccharomyces cerevisiae) Growth. J. Nutr. 1999, 129, 2236–2238. [Google Scholar] [CrossRef]
- Plantz, B.A.; Nickerson, K.; Kachman, S.D.; Schlegel, V.L. Evaluation of metals in a defined medium for Pichia pastoris expressing recombinant beta-galactosidase. Biotechnol. Prog. 2007, 23, 687–692. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Shimizu, S. Cobalt proteins. Eur. J. Biochem. 1999, 261, 1–9. [Google Scholar] [CrossRef]
- Bleackley, M.R.; MacGillivray, R.T.A. Transition metal homeostasis: From yeast to human disease. BioMetals 2011, 24, 785–809. [Google Scholar] [CrossRef]
- Zhang, Y.; Rodionov, D.A.; Gelfand, M.S.; Gladyshev, V.N. Comparative genomic analyses of nickel, cobalt and vitamin B12 utilization. BMC Genom. 2009, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Eide, D.J.; Clark, S.; Nair, T.M.; Gehl, M.; Gribskov, M.; Guerinot, M.L.; Harper, J.F. Characterization of the yeast ionome: A genome-wide analysis of nutrient mineral and trace element homeostasis in Saccharomyces cerevisiae. Genome Biol. 2005, 6, R77. [Google Scholar] [CrossRef] [Green Version]
- Conklin, D.S.; McMaster, J.A.; Culbertson, M.R.; Kung, C. COT1, a gene involved in cobalt accumulation in Saccharomyces cerevisiae. Mol. Cell Biol. 1992, 12, 3678–3688. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, R.A.; Brickey, W.W.; Walker, K.W. N-Terminal processing: The methionine aminopeptidase and Nα-acetyl transferase families. Trends Biochem. Sci. 1998, 23, 263–267. [Google Scholar] [CrossRef]
- Herrera-Camacho, I.; Morales-Monterrosas, R.; Quiróz-Alvarez, R. Aminopeptidase yscCo-II: A new cobalt-dependent aminopeptidase from yeast—purification and biochemical characterization. Yeast 2000, 16, 219–229. [Google Scholar] [CrossRef]
- Schoderboeck, L.; Mühlegger, S.; Losert, A.; Gausterer, C.; Hornek, R. Effects assessment: Boron compounds in the aquatic environment. Chemosphere 2011, 82, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Jensen, W.B. The Origin of Pyrex. J. Chem. Educ. 2006, 83, 692. [Google Scholar] [CrossRef]
- Green, G.H.; Blincoe, C.; Weeth, H.J. Boron contamination from borosilicate glass. J. Agric. Food Chem. 1976, 24, 1245–1246. [Google Scholar] [CrossRef] [PubMed]
- EuropeanChemicalsAgency. Methanol. Available online: https://echa.europa.eu/de/information-on-chemicals/cl-inventory-database/-/discli/details/37212 (accessed on 17 November 2020).
- Daly, R.; Hearn, M.T.W. Expression of heterologous proteins in Pichia pastoris: A useful experimental tool in protein engineering and production. J. Mol. Recognit. 2005, 18, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Spadiut, O.; Olsson, L.; Brumer, H. A comparative summary of expression systems for the recombinant production of galactose oxidase. Microb. Cell Factories 2010, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Spadiut, O.; Herwig, C. Production and purification of the multifunctional enzyme horseradish peroxidase. Pharm. Bioprocess 2013, 1, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Rajamanickam, V.; Winkler, M.; Flotz, P.; Meyer, L.; Herwig, C.; Spadiut, O. Comparison of Purification Strategies of Three Horseradish PeroxidaseIsoenzymes Recombinantly Produced in Pichia pastoris. J. Chromatogr. Sep. Tech. 2016, 7, 1–6. [Google Scholar]
- Capone, S.; Horvat, J.; Herwig, C.; Spadiut, O. Development of a mixed feed strategy for a recombinant Pichia pastoris strain producing with a de-repression promoter. Microb. Cell Factories 2015, 14, 101. [Google Scholar] [CrossRef] [Green Version]
- Gmeiner, C.; Saadati, A.; Maresch, D.; Krasteva, S.; Frank, M.; Altmann, F.; Herwig, C.; Spadiut, O. Development of a fed-batch process for a recombinant Pichia pastoris Deltaoch1 strain expressing a plant peroxidase. Microb. Cell Fact 2015, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Dubelaar, G.B.J.; Gerritzen, P.L. CytoBuoy: A step forward towards using flow cytometry in operational oceanography. Sci. Mar. 2000, 64, 255–265. [Google Scholar] [CrossRef]
- Pekarsky, A.; Veiter, L.; Rajamanickam, V.; Herwig, C.; Grünwald-Gruber, C.; Altmann, F.; Spadiut, O. Production of a recombinant peroxidase in different glyco-engineered Pichia pastoris strains: A morphological and physiological comparison. Microb. Cell Factories 2018, 17, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehgartner, D.; Herwig, C.; Neutsch, L. At-line determination of spore inoculum quality in Penicillium chrysogenum bioprocesses. Appl. Microbiol. Biotechnol. 2016, 100, 5363–5373. [Google Scholar] [CrossRef] [PubMed]
- Gondry, M.; Dubois, J.; Terrier, M.; Lederer, F. The catalytic role of tyrosine 254 in flavocytochrome b2 (l-lactate dehydrogenase from baker’s yeast). Eur. J. Biochem. 2001, 268, 4918–4927. [Google Scholar] [CrossRef]
- Capone, S.; Ćorajević, L.; Bonifert, G.; Murth, P.; Maresch, D.; Altmann, F.; Herwig, C.; Spadiut, O. Combining Protein and Strain Engineering for the Production of Glyco-Engineered Horseradish Peroxidase C1A in Pichia pastoris. Int. J. Mol. Sci. 2015, 16, 23127–23142. [Google Scholar] [CrossRef] [Green Version]
- Schenk, J.; Balazs, K.; Jungo, C.; Urfer, J.; Wegmann, C.; Zocchi, A.; Marison, I.W.; von Stockar, U. Influence of specific growth rate on specific productivity and glycosylation of a recombinant avidin produced by a Pichia pastoris Mut+ strain. Biotechnol. Bioeng. 2008, 99, 368–377. [Google Scholar] [CrossRef]
- Holleman, A.F. Bor. In Lehrbuch der Anorganischen Chemie; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2007; p. 1104. [Google Scholar]
- Peng, T.; Xu, Y.; Zhang, Y. Comparative genomics of molybdenum utilization in prokaryotes and eukaryotes. BMC Genom. 2018, 19, 691. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Zhang, J.; Liu, B.; Du, G.; Chen, J. High-Level Extracellular Production of Glucose Oxidase by Recombinant Pichia Pastoris Using a Combined Strategy. Appl. Biochem. Biotechnol. 2015, 175, 1429–1447. [Google Scholar] [CrossRef]
- Chen, S.; Vetro, J.A.; Chang, Y.-H. The Specificity in Vivo of Two Distinct Methionine Aminopeptidases in Saccharomyces cerevisiae. Arch. Biochem. Biophys. 2002, 398, 87–93. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivation | Bioreactor Type/Feeding | Strain | Target Protein | Co and BA Added | Comments |
|---|---|---|---|---|---|
| F1 | glass/exp. | SMD1168H Mut+ | GalOx | yes | |
| F2 | glass/exp. | no | triplicates | ||
| F3 | glass/exp. | no | |||
| F4 | glass/exp. | no | |||
| F5 | glass/pulsed | BSYBG11 MutS | HRP | yes | |
| F6 | glass/pulsed | no | |||
| F7 | glass/pulsed | ATUM PPS 9011 MutS | FC | yes | ICP-OES; flow cytometry |
| F8 | glass/pulsed | no | |||
| F9 | steel/exp. | BSYBG11 MutS | HRP | yes | |
| F10 | steel/exp. | no |
| B | Fe | Mn | Co | Cu | Zn | Mo | Mg | Ca | K | |
|---|---|---|---|---|---|---|---|---|---|---|
| [mg·L−1] | ||||||||||
| Added | 0.02 | 56.6 | 4.2 | 1.0 | 6.6 | 41.6 | 0.3 | 441.8 | 41.9 | 5933.3 |
| Measured | 4.90 ± 0.57 | <L | 2.3 ± 0.2 | 0.6 | 1.8 ± 0.3 | 5.5 ± 0.8 | 0.3 ± 0.1 | 468.1 ± 25.8 | 48.2 ± 6.8 | 8333.2 ± 535.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pekarsky, A.; Mihalyi, S.; Weiss, M.; Limbeck, A.; Spadiut, O. Depletion of Boric Acid and Cobalt from Cultivation Media: Impact on Recombinant Protein Production with Komagataella phaffii. Bioengineering 2020, 7, 161. https://doi.org/10.3390/bioengineering7040161
Pekarsky A, Mihalyi S, Weiss M, Limbeck A, Spadiut O. Depletion of Boric Acid and Cobalt from Cultivation Media: Impact on Recombinant Protein Production with Komagataella phaffii. Bioengineering. 2020; 7(4):161. https://doi.org/10.3390/bioengineering7040161
Chicago/Turabian StylePekarsky, Alexander, Sophia Mihalyi, Maximilian Weiss, Andreas Limbeck, and Oliver Spadiut. 2020. "Depletion of Boric Acid and Cobalt from Cultivation Media: Impact on Recombinant Protein Production with Komagataella phaffii" Bioengineering 7, no. 4: 161. https://doi.org/10.3390/bioengineering7040161
APA StylePekarsky, A., Mihalyi, S., Weiss, M., Limbeck, A., & Spadiut, O. (2020). Depletion of Boric Acid and Cobalt from Cultivation Media: Impact on Recombinant Protein Production with Komagataella phaffii. Bioengineering, 7(4), 161. https://doi.org/10.3390/bioengineering7040161