

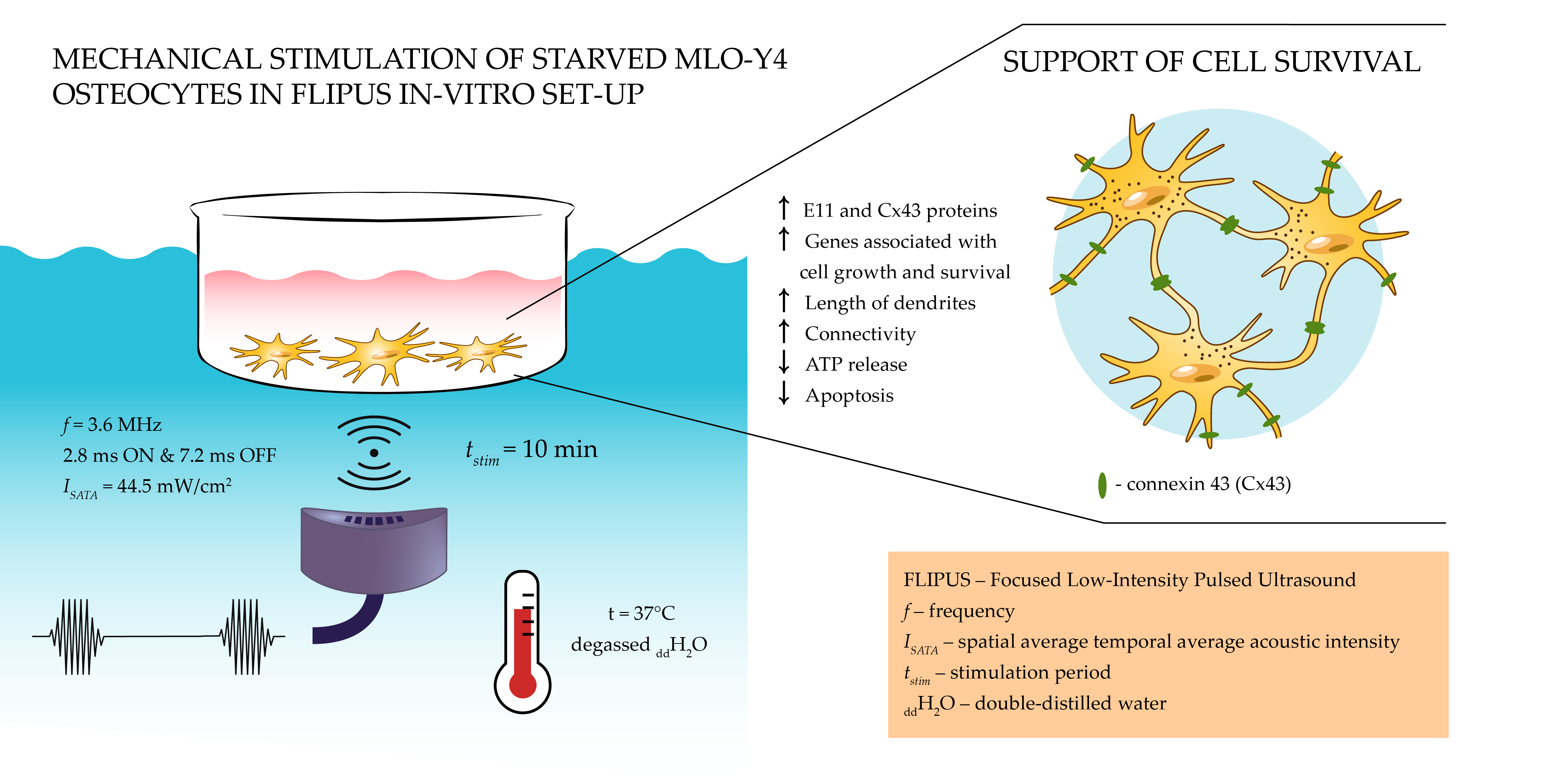
Focused Low-Intensity Pulsed Ultrasound (FLIPUS) Mitigates Apoptosis of MLO-Y4 Osteocyte-like Cells

Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Focused Low-Intensity Pulsed Ultrasound In Vitro Set-Up
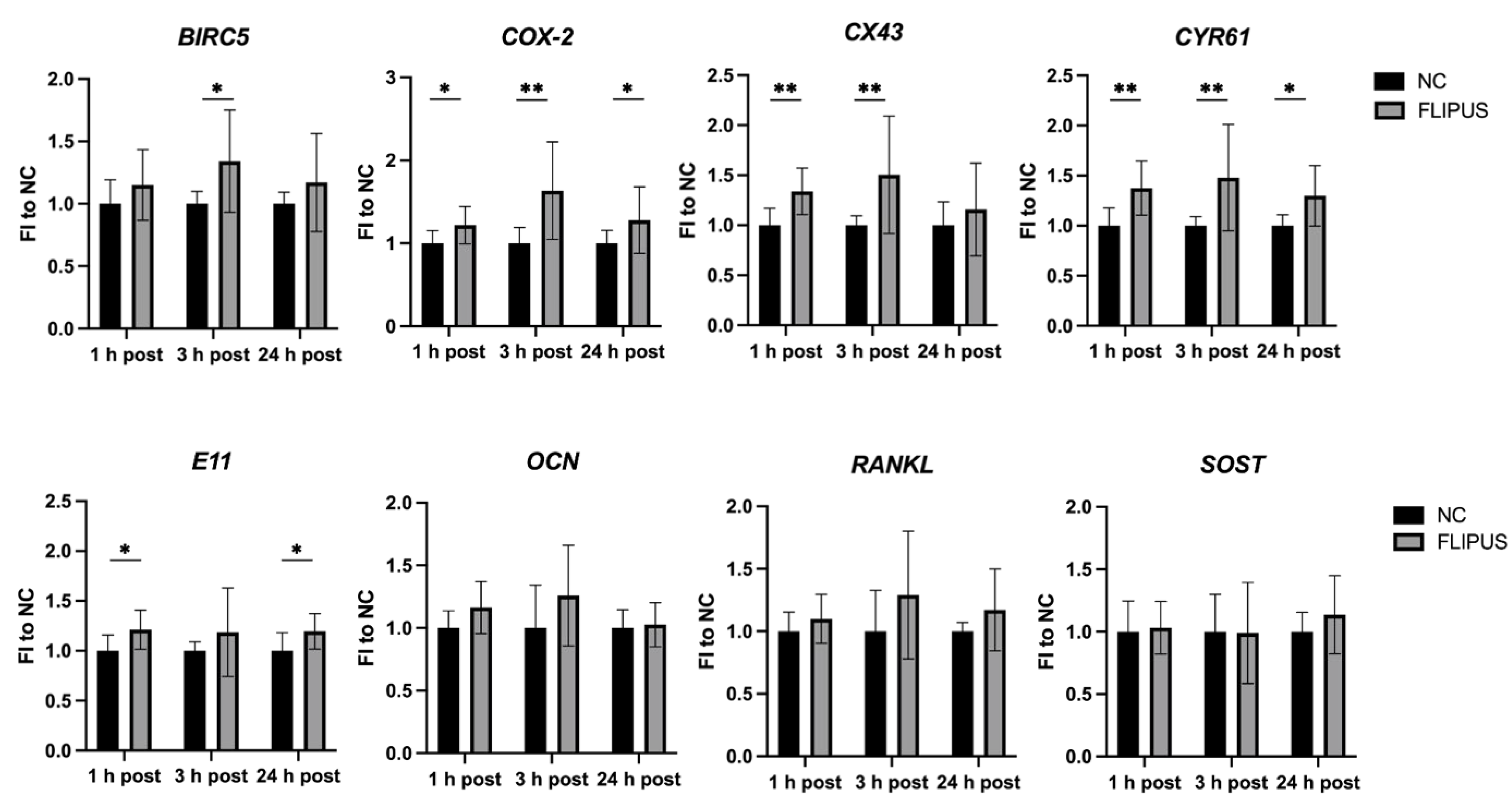
2.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. WST-8 Viability Assay
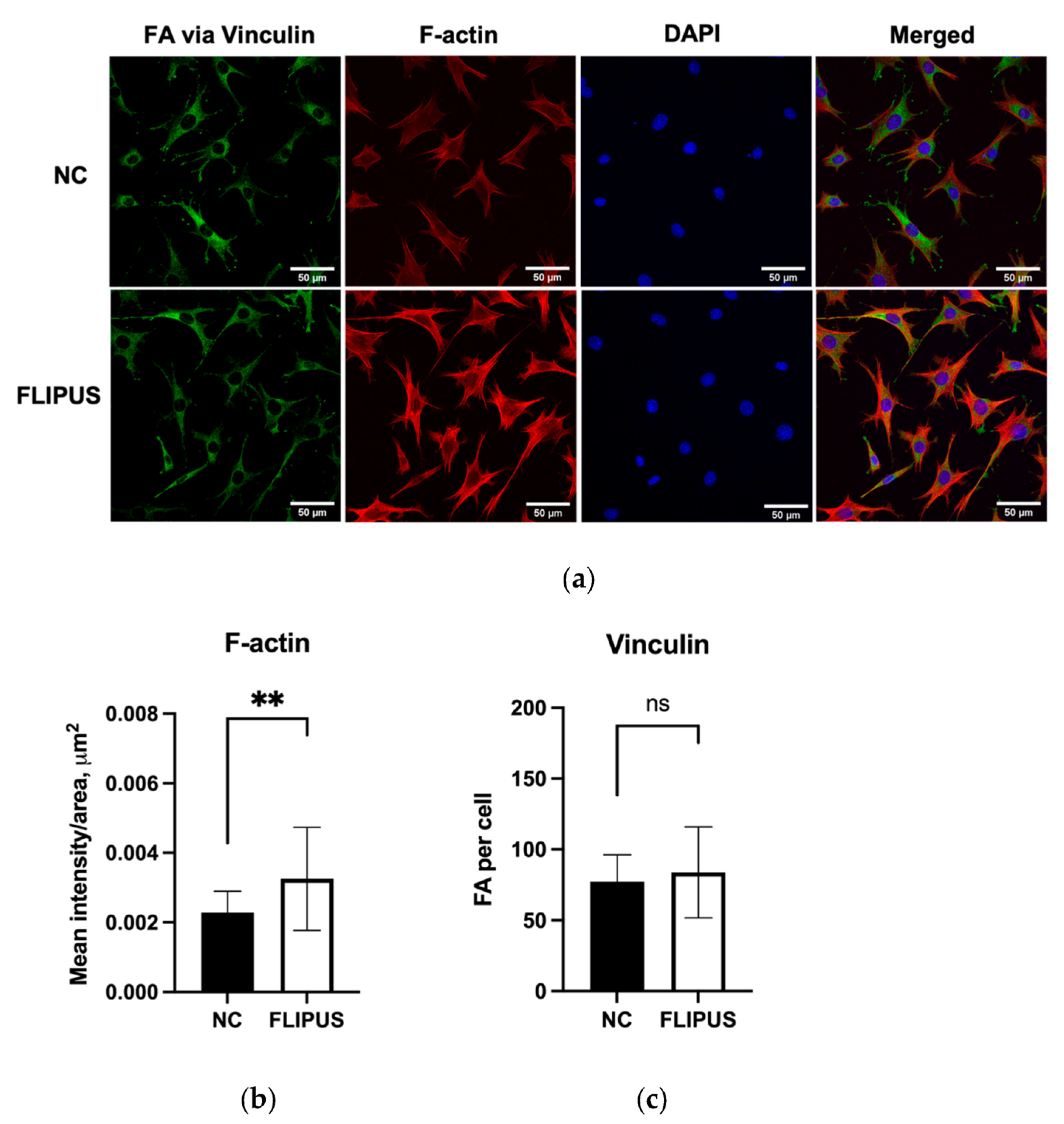
2.5. Fluorescence Staining
2.6. Dye Uptake Assay
2.7. Apoptosis Assay
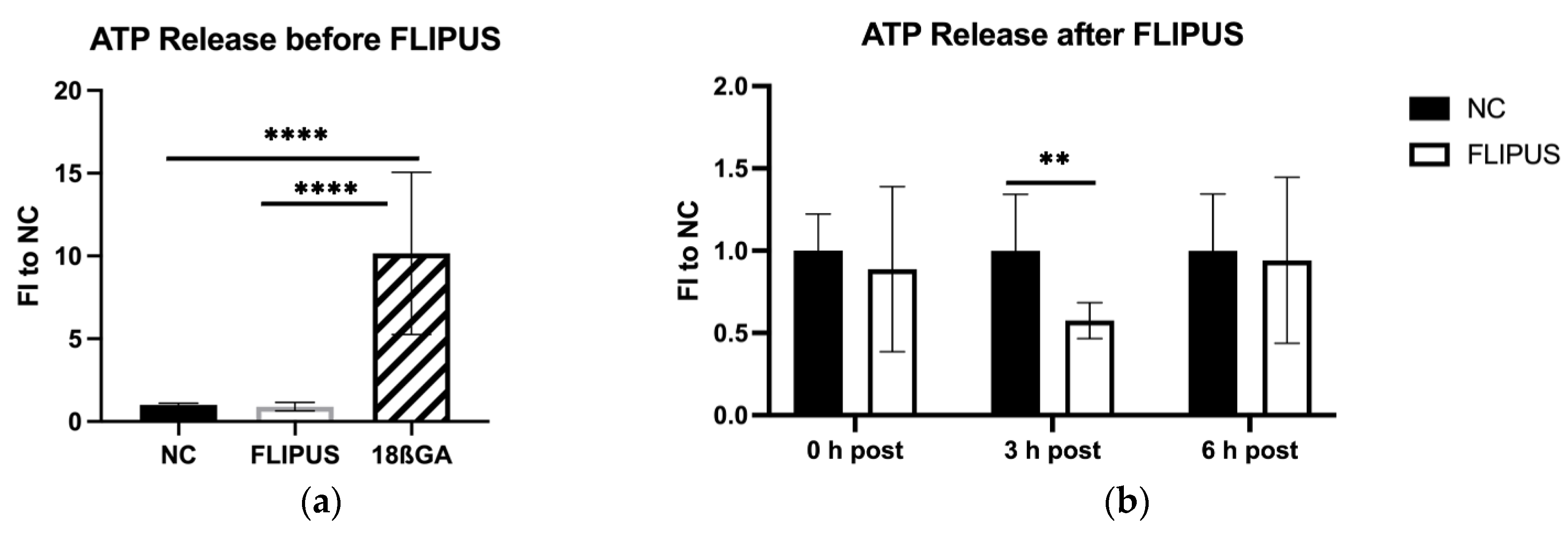
2.8. ATP Release Assay
2.9. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | NC Non-Starv | FLIPUS Non-Starv | NC Starv | FLIPUS Starv | ||||
|---|---|---|---|---|---|---|---|---|
| MNE | ± SD | MNE | ± SD | MNE | ± SD | MNE | ± SD | |
| C-JUN | 0.4099 | 0.4105 | 0.7510 | 0.8272 | 0.1145 | 0.0435 | 0.1466 | 0.0672 |
| C-FOS | 0.0106 | 0.0087 | 0.0182 | 0.0195 | 0.0052 | 0.0017 | 0.0068 | 0.0015 |
| C-MYC | 0.1394 | 0.1273 | 0.2203 | 0.1734 | 0.2900 | 0.0860 | 0.4054 | 0.1368 |
| COX-2 | 0.3130 | 0.0537 | 0.3524 | 0.0956 | 0.2211 | 0.0530 | 0.2930 | 0.0917 |
| CYR61 | 1.1436 | 0.5831 | 1.8863 | 1.1375 | 2.5143 | 0.8496 | 3.4944 | 1.3695 |
| Gene Name | 1 h Post, MNE | 3 h Post, MNE | 24 h Post, MNE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NC | ± SD | FLIPUS | ± SD | NC | ± SD | FLIPUS | ± SD | NC | ± SD | FLIPUS | ± SD | |
| BIRC5 | 3.389 | 1.357 | 3.892 | 1.041 | 5.022 | 1.896 | 6.638 | 3.177 | 3.178 | 1.230 | 3.573 | 1.420 |
| COX-2 | 0.246 | 0.100 | 0.300 | 0.085 | 0.134 | 0.086 | 0.201 | 0.095 | 0.063 | 0.029 | 0.079 | 0.038 |
| CX43 | 1.215 | 0.552 | 1.587 | 0.391 | 3.181 | 1.834 | 4.110 | 2.164 | 1.322 | 1.269 | 1.560 | 1.509 |
| CYR61 | 1.451 | 0.669 | 1.916 | 0.365 | 1.621 | 0.712 | 2.219 | 0.741 | 0.560 | 0.313 | 0.729 | 0.440 |
| E11 | 0.707 | 0.412 | 0.919 | 0.339 | 1.029 | 0.405 | 1.160 | 0.438 | 0.375 | 0.188 | 0.449 | 0.218 |
| OCN | 0.041 | 0.025 | 0.047 | 0.023 | 0.096 | 0.061 | 0.107 | 0.061 | 0.367 | 0.236 | 0.372 | 0.215 |
| RANKL | 0.473 | 0.259 | 0.519 | 0.198 | 0.360 | 0.143 | 0.450 | 0.311 | 0.774 | 0.262 | 0.886 | 0.350 |
| SOST | 0.309 | 0.123 | 0.332 | 0.072 | 0.167 | 0.068 | 0.152 | 0.050 | 0.123 | 0.038 | 0.133 | 0.028 |
References
- Schaffler, M.B.; Cheung, W.Y.; Majeska, R.; Kennedy, O. Osteocytes: Master orchestrators of bone. Calcif. Tissue Int. 2014, 94, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Burra, S.; Nicolella, D.P.; Francis, W.L.; Freitas, C.J.; Mueschke, N.J.; Poole, K.; Jiang, J.X. Dendritic processes of osteocytes are mechanotransducers that induce the opening of hemichannels. Proc. Natl. Acad. Sci. USA 2010, 107, 13648–13653. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.R.; Rubin, C.T.; Rubin, J. Mechanical regulation of signaling pathways in bone. Gene 2012, 503, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. Bone “mass” and the “mechanostat”: A proposal. Anat. Rec. 1987, 219, 1–9. [Google Scholar] [CrossRef]
- Qin, L.; Liu, W.; Cao, H.; Xiao, G. Molecular mechanosensors in osteocytes. Bone Res. 2020, 8, 23. [Google Scholar] [CrossRef]
- Bonewald, L.F. Establishment and characterization of an osteocyte-like cell line, MLO-Y4. J. Bone Miner. Metab. 1999, 17, 61–65. [Google Scholar] [CrossRef]
- Lampe, P.D.; Lau, A.F. Regulation of gap junctions by phosphorylation of connexins. Arch. Biochem. Biophys. 2000, 384, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, L.I.; Manolagas, S.C.; Bellido, T. Transduction of cell survival signals by connexin-43 hemichannels. J. Biol. Chem. 2002, 277, 8648–8657. [Google Scholar] [CrossRef]
- Xu, H.; Gu, S.; Riquelme, M.A.; Burra, S.; Callaway, D.; Cheng, H.; Guda, T.; Schmitz, J.; Fajardo, R.J.; Werner, S.L.; et al. Connexin 43 channels are essential for normal bone structure and osteocyte viability. J. Bone Miner. Res. 2015, 30, 436–448. [Google Scholar] [CrossRef]
- Hua, R.; Zhang, J.; Riquelme, M.A.; Jiang, J.X. Connexin gap junctions and hemichannels link oxidative stress to skeletal physiology and pathology. Curr. Osteoporos. Rep. 2021, 19, 66–74. [Google Scholar] [CrossRef]
- Cheng, B.; Zhao, S.; Luo, J.; Sprague, E.; Bonewald, L.F.; Jiang, J.X. Expression of functional gap junctions and regulation by fluid flow in osteocyte-like MLO-Y4 cells. J. Bone Miner. Res. 2001, 16, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Kato, Y.; Zhao, S.; Luo, J.; Sprague, E.; Bonewald, L.F.; Jiang, J.X. PGE(2) is essential for gap junction-mediated intercellular communication between osteocyte-like MLO-Y4 cells in response to mechanical strain. Endocrinology 2001, 142, 3464–3473. [Google Scholar] [CrossRef] [PubMed]
- Cherian, P.P.; Cheng, B.; Gu, S.; Sprague, E.; Bonewald, L.F.; Jiang, J.X. Effects of mechanical strain on the function of Gap junctions in osteocytes are mediated through the prostaglandin EP2 receptor. J. Biol. Chem. 2003, 278, 43146–43156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Barragan-Adjemian, C.; Ye, L.; Kotha, S.; Dallas, M.; Lu, Y.; Zhao, S.; Harris, M.; Harris, S.E.; Feng, J.Q.; et al. E11/gp38 selective expression in osteocytes: Regulation by mechanical strain and role in dendrite elongation. Mol. Cell. Biol. 2006, 26, 4539–4552. [Google Scholar] [CrossRef]
- Wehmeyer, C.; Naylor, A.; Moeller, K.; Poologasundarampillai, G.; Pap, T.; Buckley, C. P124/O27 Osteocyte-derived podoplanin is an important regulator of bone remodelling in the KBxN serum transfer model of rheumatoid arthritis. Ann. Rheum. Dis. 2019, 78 (Suppl. 1), A55–A56. [Google Scholar]
- Padilla, F.; Puts, R.; Vico, L.; Guignandon, A.; Raum, K. Stimulation of bone repair with ultrasound. Adv. Exp. Med. Biol. 2016, 880, 385–427. [Google Scholar]
- Poolman, R.W.; Agoritsas, T.; Siemieniuk, R.A.; Harris, I.A.; Schipper, I.B.; Mollon, B.; Smith, M.; Albin, A.; Nador, S.; Sasges, W.; et al. Low intensity pulsed ultrasound (LIPUS) for bone healing: A clinical practice guideline. BMJ 2017, 356, j576. [Google Scholar] [CrossRef]
- Trust Investigators Writing Group; Busse, J.W.; Bhandari, M.; Einhorn, T.A.; Schemitsch, E.; Heckman, J.D.; Tornetta, P., 3rd; Leung, K.S.; Heels-Ansdell, D.; Makosso-Kallyth, S.; et al. Re-evaluation of low intensity pulsed ultrasound in treatment of tibial fractures (TRUST): Randomized clinical trial. BMJ 2016, 355, i5351. [Google Scholar]
- Schandelmaier, S.; Kaushal, A.; Lytvyn, L.; Heels-Ansdell, D.; Siemieniuk, R.A.; Agoritsas, T.; Guyatt, G.H.; Vandvik, P.O.; Couban, R.; Mollon, B.; et al. Low intensity pulsed ultrasound for bone healing: Systematic review of randomized controlled trials. BMJ 2017, 356, j656. [Google Scholar] [CrossRef]
- Puts, R.; Vico, R.; Beilfuss, N.; Shaka, M.; Padilla, F.; Raum, K. Pulsed ultrasound for bone regeneration—Outcomes and hurdles in the clinical application: A systematic review. Eur. Cell. Mater. 2021, 42, 281–311. [Google Scholar] [CrossRef]
- Puts, R.; Ruschke, K.; Ambrosi, T.H.; Kadow-Romacker, A.; Knaus, P.; Jenderka, K.V.; Raum, K. A Focused low-intensity pulsed ultrasound (FLIPUS) system for cell stimulation: Physical and biological proof of principle. IEEE Trans. Ultrason. Ferroelectr. Freq. Control. 2016, 63, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Padilla, F.; Puts, R.; Vico, L.; Raum, K. Stimulation of bone repair with ultrasound: A review of the possible mechanic effects. Ultrasonics 2014, 54, 1125–1145. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Fujita, N.; Tsuji-Tamura, K.; Kitagawa, Y.; Fujisawa, T.; Tamura, M.; Sato, M. Osteocytes as main responders to low-intensity pulsed ultrasound treatment during fracture healing. Sci. Rep. 2021, 11, 10298. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, Z.; Zhang, H.; Chen, W.; Chen, M.; Zhu, Z. Low-intensity pulsed ultrasound regulates proliferation and differentiation of osteoblasts through osteocytes. Biochem. Biophys. Res. Commun. 2012, 418, 296–300. [Google Scholar] [CrossRef]
- Fung, C.H.; Cheung, W.H.; Pounder, N.M.; Harrison, A.; Leung, K.S. Osteocytes exposed to far field of therapeutic ultrasound promotes osteogenic cellular activities in pre-osteoblasts through soluble factors. Ultrasonics 2014, 54, 1358–1365. [Google Scholar] [CrossRef]
- Saini, V.; Yadav, S.; McCormick, S. Low-intensity pulsed ultrasound modulates shear stress induced PGHS-2 expression and PGE2 synthesis in MLO-Y4 osteocyte-like cells. Ann. Biomed. Eng. 2011, 39, 378–393. [Google Scholar] [CrossRef]
- Kato, Y.; Windle, J.J.; Koop, B.A.; Mundy, G.R.; Bonewald, L.F. Establishment of an osteocyte-like cell line, MLO-Y4. J. Bone Miner. Res. 1997, 12, 2014–2023. [Google Scholar] [CrossRef]
- Huang, C.; Geng, J.; Wei, X.; Zhang, R.; Jiang, S. MiR-144-3p regulates osteogenic differentiation and proliferation of murine mesenchymal stem cells by specifically targeting Smad4. FEBS Lett. 2016, 590, 795–807. [Google Scholar] [CrossRef]
- Kang, J.; Boonanantanasarn, K.; Baek, K.; Woo, K.M.; Ryoo, H.M.; Baek, J.H.; Kim, G.S. Hyperglycemia increases the expression levels of sclerostin in a reactive oxygen species- and tumor necrosis factor-alpha-dependent manner. J. Periodontal Implant. Sci. 2015, 45, 101–110. [Google Scholar] [CrossRef]
- Cheung, W.Y.; Fritton, J.C.; Morgan, S.A.; Seref-Ferlengez, Z.; Basta-Pljakic, J.; Thi, M.M.; Suadicani, S.O.; Spray, D.C.; Majeska, R.J.; Schaffler, M.B. Pannexin-1 and P2X7-receptor are required for apoptotic osteocytes in fatigued bone to trigger RANKL production in neighboring bystander osteocytes. J. Bone Miner. Res. 2016, 31, 890–899. [Google Scholar] [CrossRef]
- Puts, R.; Rikeit, P.; Ruschke, K.; Kadow-Romacker, A.; Hwang, S.; Jenderka, K.V.; Knaus, P.; Raum, K. Activation of mechanosensitive transcription factors in murine C2C12 mesenchymal precursors by focused low-intensity pulsed ultrasound (FLIPUS). IEEE Trans. Ultrason. Ferroelectr. Freq. Control 2016, 63, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Puts, R.; Rikeit, P.; Ruschke, K.; Knaus, P.; Schreivogel, S.; Raum, K. Functional regulation of YAP mechanosensitive transcriptional coactivator by Focused Low-Intensity Pulsed Ultrasound (FLIPUS) enhances proliferation of murine mesenchymal precursors. PLoS ONE 2018, 13, e0206041. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.U.; Coombs, K.M. Serum-reduced media impacts on cell viability and protein expression in human lung epithelial cells. J. Cell. Physiol. 2019, 234, 7718–7724. [Google Scholar] [CrossRef] [PubMed]
- Puts, R.; Ambrosi, T.; Kadow-Romacker, A.; Raum, K.; Ruschke, K.; Knaus, P. In-vitro stimulation of cells of the musculoskeletal system with focused low-intensity pulsed ultrasound (FLIPUS): Analyses of cellular activities in response to the optimized acoustic dose. In Proceedings of the IEEE International Ultrasonics Symposium, Chicago, IL, USA, 3–6 September 2014; pp. 1630–1633. [Google Scholar]
- Zhou, Z.Q.; Shung, C.Y.; Ota, S.; Akiyama, H.; Keene, D.R.; Hurlin, P.J. Sequential and coordinated actions of c-Myc and N-Myc control appendicular skeletal development. PLoS ONE 2011, 6, e18795. [Google Scholar] [CrossRef]
- Piek, E.; Sleumer, L.S.; van Someren, E.P.; Heuver, L.; de Haan, J.R.; de Grijs, I.; Gilissen, C.; Hendriks, J.M.; van Ravestein-van Os, R.I.; Bauerschmidt, S.; et al. Osteo-transcriptomics of human mesenchymal stem cells: Accelerated gene expression and osteoblast differentiation induced by vitamin D reveals c-MYC as an enhancer of BMP2-induced osteogenesis. Bone 2010, 46, 613–627. [Google Scholar] [CrossRef]
- Melnik, S.; Werth, N.; Boeuf, S.; Hahn, E.M.; Gotterbarm, T.; Anton, M.; Richter, W. Impact of c-MYC expression on proliferation, differentiation, and risk of neoplastic transformation of human mesenchymal stromal cells. Stem Cell. Res. Ther. 2019, 10, 73. [Google Scholar] [CrossRef]
- Wagner, E.F. Functions of AP1 (Fos/Jun) in bone development. Ann. Rheum. Dis. 2002, 61 (Suppl. 2), ii40–ii42. [Google Scholar] [CrossRef]
- Morello, D.; Fitzgerald, M.J.; Babinet, C.; Fausto, N. c-myc, c-fos, and c-jun regulation in the regenerating livers of normal and H-2K/c-myc transgenic mice. Mol. Cell. Biol. 1990, 10, 3185–3193. [Google Scholar]
- Hadjiargyrou, M.; Ahrens, W.; Rubin, C.T. Temporal expression of the chondrogenic and angiogenic growth factor CYR61 during fracture repair. J. Bone Miner. Res. 2000, 15, 1014–1023. [Google Scholar] [CrossRef]
- Zarka, M.; Hay, E.; Cohen-Solal, M. YAP/TAZ in bone and cartilage biology. Front. Cell. Dev. Biol. 2021, 9, 788773. [Google Scholar] [CrossRef]
- Bartkowiak, K.; Heidrich, I.; Kwiatkowski, M.; Gorges, T.M.; Andreas, A.; Geffken, M.; Verpoort, K.; Muller, V.; Schluter, H.; Pantel, K. Cysteine-rich angiogenic Inducer 61: Pro-survival function and role as a biomarker for disseminating breast cancer cells. Cancers 2021, 13, 563. [Google Scholar] [CrossRef] [PubMed]
- Kitase, Y.; Barragan, L.; Qing, H.; Kondoh, S.; Jiang, J.X.; Johnson, M.L.; Bonewald, L.F. Mechanical induction of PGE2 in osteocytes blocks glucocorticoid-induced apoptosis through both the beta-catenin and PKA pathways. J. Bone Miner. Res. 2010, 25, 2657–2668. [Google Scholar] [CrossRef]
- Jiang, J.X.; Cheng, B. Mechanical stimulation of gap junctions in bone osteocytes is mediated by prostaglandin E2. Cell. Commun. Adhes. 2001, 8, 283–288. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, R.; Li, G.; Chen, M.; Shang, P.; Yang, H.; Jiang, J.X.; Xu, H. Connexin 43 channels in osteocytes regulate bone responses to mechanical unloading. Front. Physiol. 2020, 11, 299. [Google Scholar] [CrossRef]
- Civitelli, R.; Ziambaras, K.; Warlow, P.M.; Lecanda, F.; Nelson, T.; Harley, J.; Atal, N.; Beyer, E.C.; Steinberg, T.H. Regulation of connexin43 expression and function by prostaglandin E2 (PGE2) and parathyroid hormone (PTH) in osteoblastic cells. J. Cell. Biochem. 1998, 68, 8–21. [Google Scholar] [CrossRef]
- Yao, H.; Zhang, L.; Yan, S.; He, Y.; Zhu, H.; Li, Y.; Wang, D.; Yang, K. Low-intensity pulsed ultrasound/nanomechanical force generators enhance osteogenesis of BMSCs through microfilaments and TRPM7. J. Nanobiotechnol. 2022, 20, 378. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, L.I.; Mathov, I.; Aguirre, J.I.; Parfitt, A.M.; Manolagas, S.C.; Bellido, T. Mechanical stimulation prevents osteocyte apoptosis: Requirement of integrins, Src kinases, and ERKs. Am. J. Physiol. Cell. Physiol. 2005, 289, C633–C643. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Riquelme, M.A.; Gu, S.; Kar, R.; Gao, X.; Sun, L.; Jiang, J.X. Osteocytic connexin hemichannels suppress breast cancer growth and bone metastasis. Oncogene 2016, 35, 5597–5607. [Google Scholar] [CrossRef]
- Ayna, G.; Krysko, D.V.; Kaczmarek, A.; Petrovski, G.; Vandenabeele, P.; Fesus, L. ATP release from dying autophagic cells and their phagocytosis are crucial for inflammasome activation in macrophages. PLoS ONE 2012, 7, e40069. [Google Scholar] [CrossRef]
- McCutcheon, S.; Majeska, R.J.; Spray, D.C.; Schaffler, M.B.; Vazquez, M. Apoptotic osteocytes induce RANKL production in bystanders via purinergic signaling and activation of pannexin channels. J. Bone Miner. Res. 2020, 35, 966–977. [Google Scholar] [CrossRef]
- Feng, Y.; Mei, L.; Wang, M.; Huang, Q.; Huang, R. Anti-inflammatory and pro-apoptotic effects of 18beta-glycyrrhetinic acid in vitro and in vivo models of rheumatoid arthritis. Front. Pharmacol. 2021, 12, 681525. [Google Scholar] [CrossRef]
- Lee, C.S.; Kim, Y.J.; Lee, M.S.; Han, E.S.; Lee, S.J. 18beta-glycyrrhetinic acid induces apoptotic cell death in SiHa cells and exhibits a synergistic effect against antibiotic anti-cancer drug toxicity. Life Sci. 2008, 83, 481–489. [Google Scholar] [CrossRef]
- Wang, D.; Wong, H.K.; Feng, Y.B.; Zhang, Z.J. 18beta-glycyrrhetinic acid induces apoptosis in pituitary adenoma cells via ROS/MAPKs-mediated pathway. J. Neurooncol. 2014, 116, 221–230. [Google Scholar] [CrossRef]









| Gene | Forward (F) and Reverse (R) Primers |
|---|---|
| Proto-oncogene C-JUN | F: 5′–TGTTTGTTTGTTTGGGTGTCC–3′ R: 5′–GAGGTTGGGGGCTACTTTTC–3′ |
| Proto-Oncogene C-FOS | F: 5′–AATGGTGAAGACCGTGTCAG–3′ R: 5′–CAGCCATCTTATTCCGTTCC–3′ |
| Cellular myelocytomatosis oncogene (C-MYC) | F: 5′–GCTGTTTGAAGGCTGGATTT–3′ R: 5′–CTCTGCTGTTGCTGGTGATAG–3′ |
| Cyclooxygenase 2 (COX-2) | F: 5′–ATCCTGAGTGGGGTGATGAG–3′ R: 5′–GGAACTGCTGGTTGAAAAGG–3′ |
| Cysteine-rich angiogenic inducer 61 (CYR61) | F: 5′–TGCTGTAAGGTCTGCGCTAA–3′ R: 5′–AGGGTCTGCCTTCTGACTGA–3′ |
| Baculoviral inhibitor of apoptosis repeat-containing 5 (BIRC5) | F: 5′–CATCGCCACCTTCAAGAACT–3′ R: 5′–AAAACACTGGGCCAAATCAG–3′ |
| Connexin 43 (CX43) | F: 5′–CGGTTGTGAAAATGTCTGCTATG–3′ R: 5′–GGCACAGACACGAATATGATCTG–3′ |
| Podoplanin (E11) | F: 5′–TTGGAATCATAGTTGGCGTCT–3′ R: 5′–TTGGAATCATAGTTGGCGTCT–3′ |
| Osteocalcin (OCN) | F: 5′–GCAGGAGGGCAATAAGGTAG–3′ R: 5′–CCATAGATGCGTTTGTAGGC–3′ |
| Sclerostine (SOST) | F: 5′–AGCCTTCAGGAATGATGCCAC–3′ R: 5′–CTTTGGCGTCATAGGGATGGT–3′ |
| Receptor Activator of Nuclear Kappa- B Ligand (RANKL) | F: 5′–AGGAGGGAGCACGAAAAACT–3′ R: 5′–AAGGGTTGGACACCTGAATG–3′ |
| Hypoxanthine-guanine-phosphoribosyltransferase (HPRT) | F: 5′–TGTTGTTGGATATGCCCTTG–3′ R: 5′–ACTGGCAACATCAACAGGACT–3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puts, R.; Khaffaf, A.; Shaka, M.; Zhang, H.; Raum, K. Focused Low-Intensity Pulsed Ultrasound (FLIPUS) Mitigates Apoptosis of MLO-Y4 Osteocyte-like Cells. Bioengineering 2023, 10, 387. https://doi.org/10.3390/bioengineering10030387
Puts R, Khaffaf A, Shaka M, Zhang H, Raum K. Focused Low-Intensity Pulsed Ultrasound (FLIPUS) Mitigates Apoptosis of MLO-Y4 Osteocyte-like Cells. Bioengineering. 2023; 10(3):387. https://doi.org/10.3390/bioengineering10030387
Chicago/Turabian StylePuts, Regina, Aseel Khaffaf, Maria Shaka, Hui Zhang, and Kay Raum. 2023. "Focused Low-Intensity Pulsed Ultrasound (FLIPUS) Mitigates Apoptosis of MLO-Y4 Osteocyte-like Cells" Bioengineering 10, no. 3: 387. https://doi.org/10.3390/bioengineering10030387
APA StylePuts, R., Khaffaf, A., Shaka, M., Zhang, H., & Raum, K. (2023). Focused Low-Intensity Pulsed Ultrasound (FLIPUS) Mitigates Apoptosis of MLO-Y4 Osteocyte-like Cells. Bioengineering, 10(3), 387. https://doi.org/10.3390/bioengineering10030387