
In Vivo Assessment of Laboratory-Grown Kidney Tissue Grafts

 ,
,
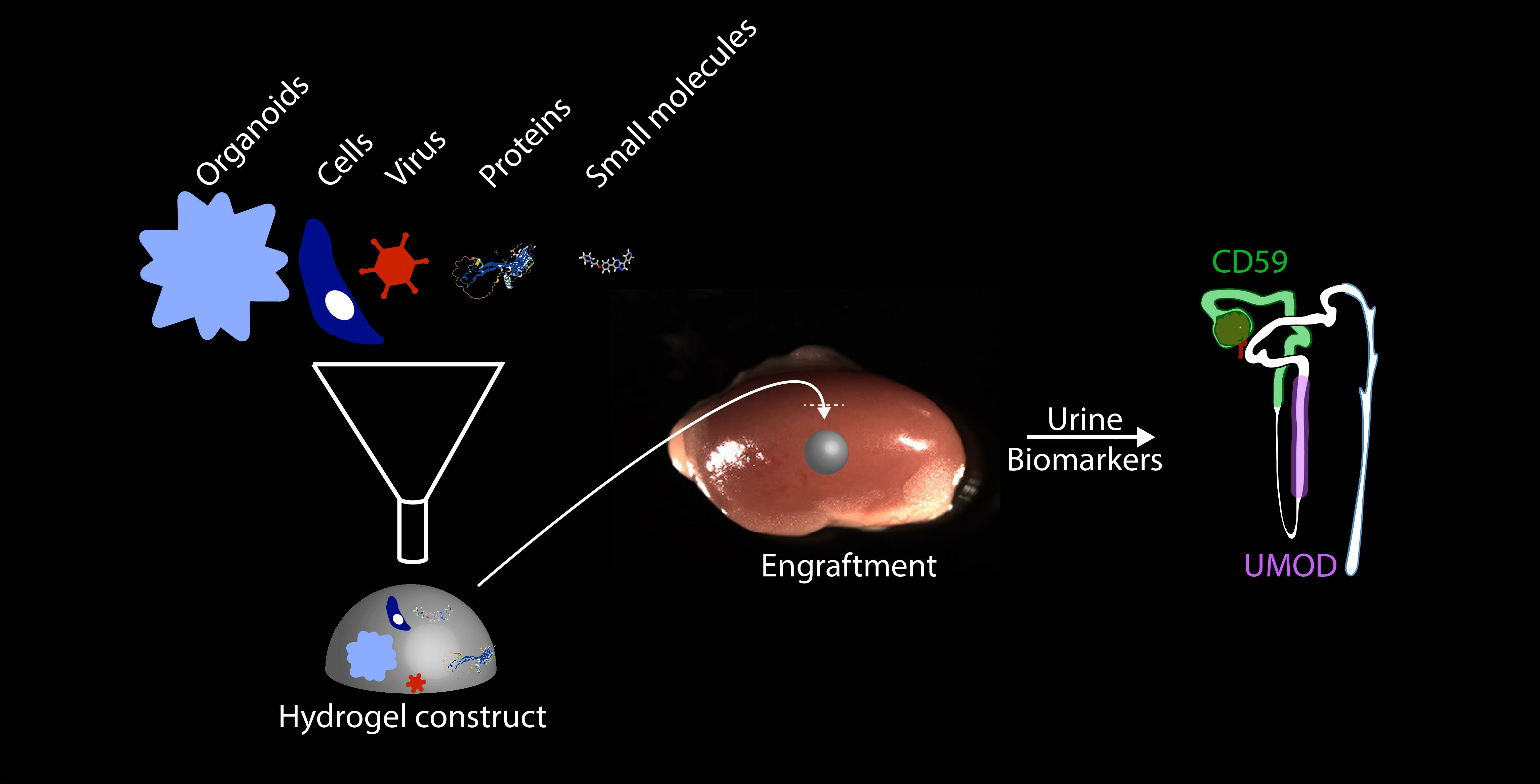
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Preparation and Gelation of Fibrinogen
2.3. Culture of Kidney Organoids
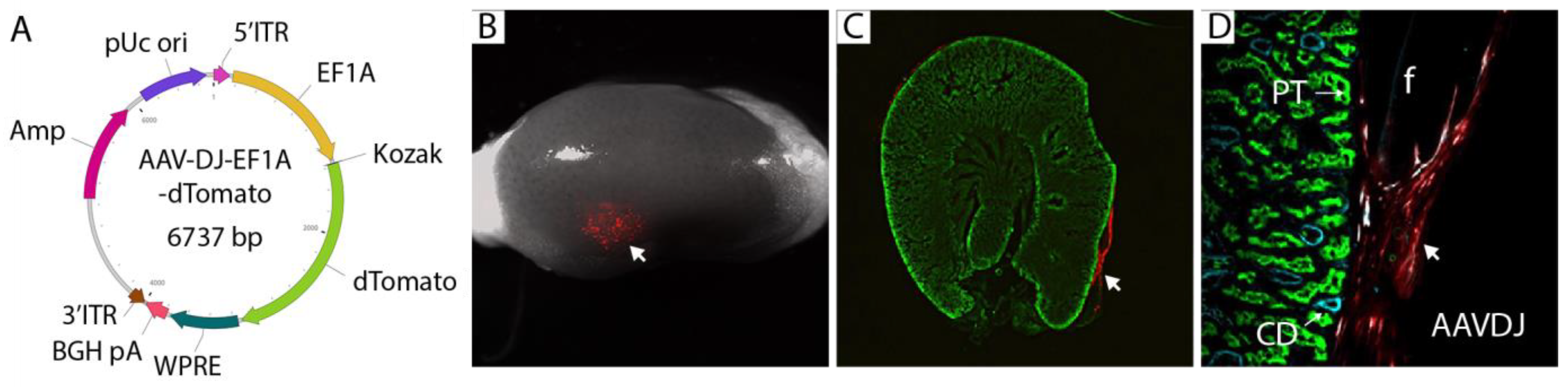
2.4. Adeno Associated Virus (AAV)
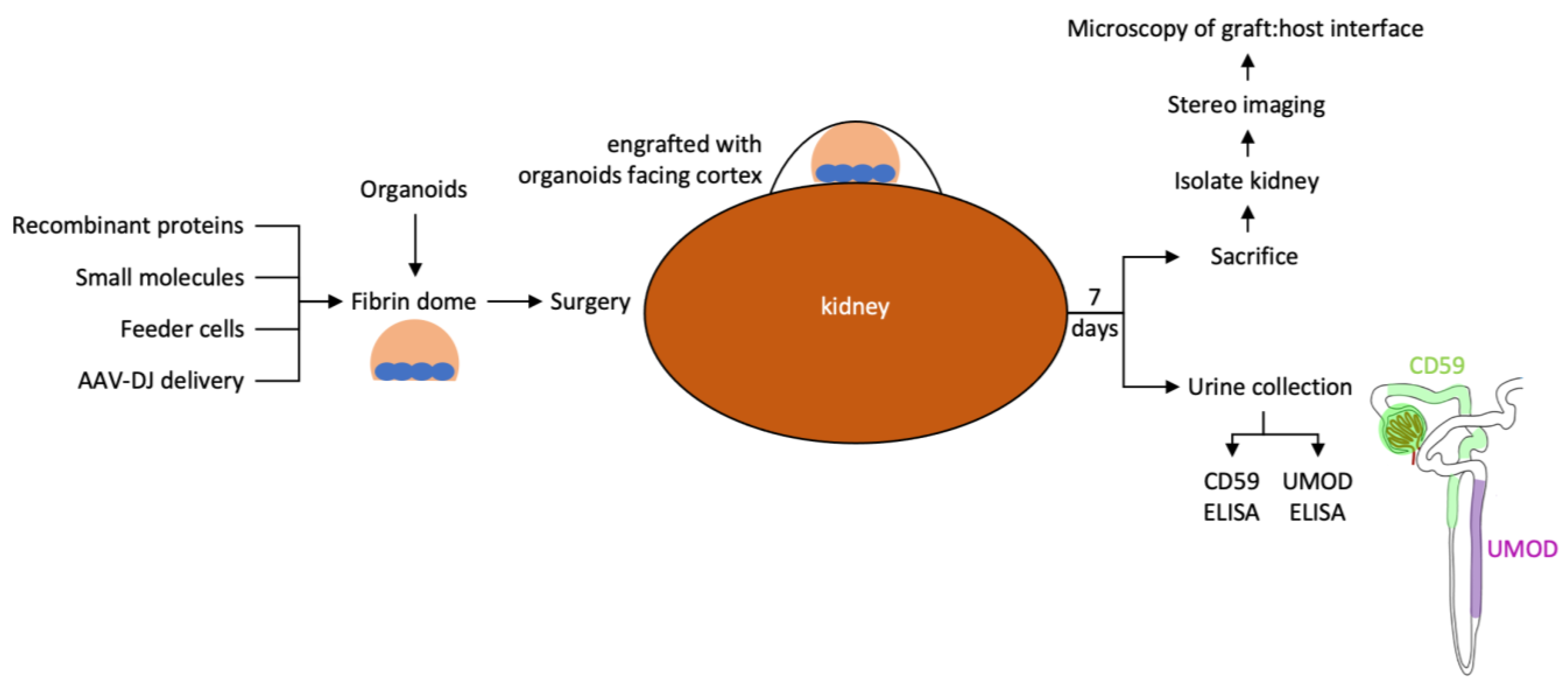
2.5. Surgery
2.6. Tissue Preparation and Staining
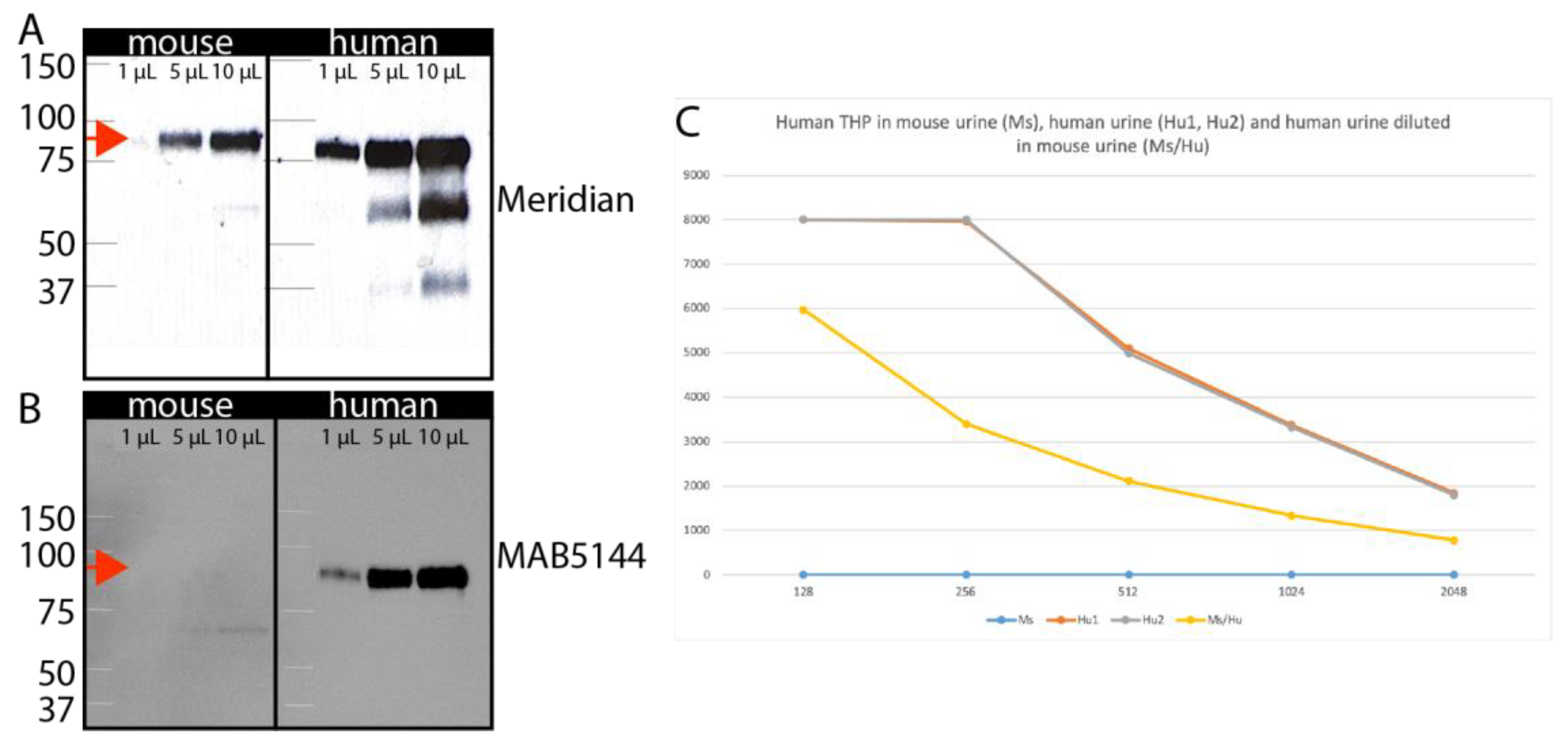
2.7. Urine Samples
2.8. ELISA
3. Results
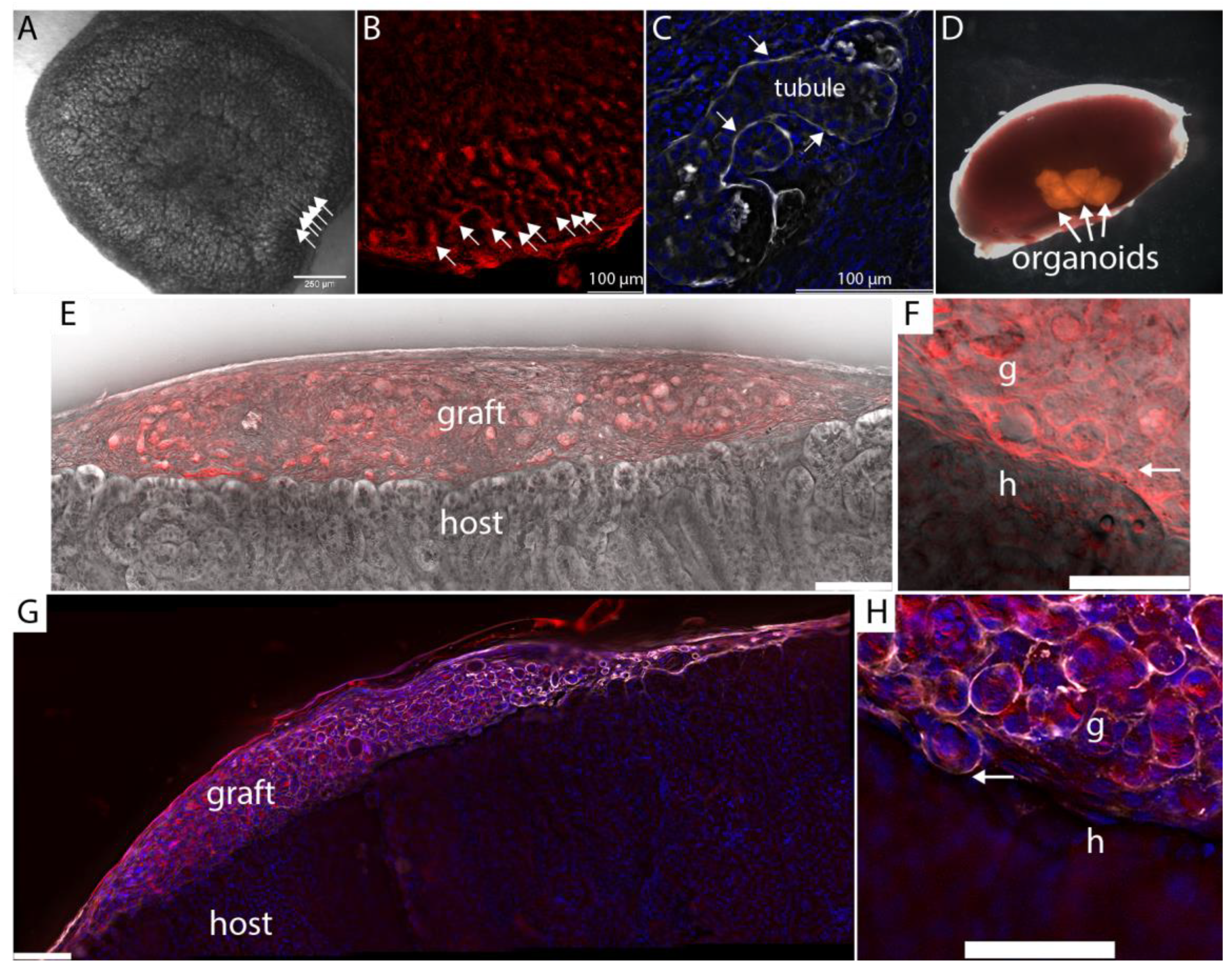
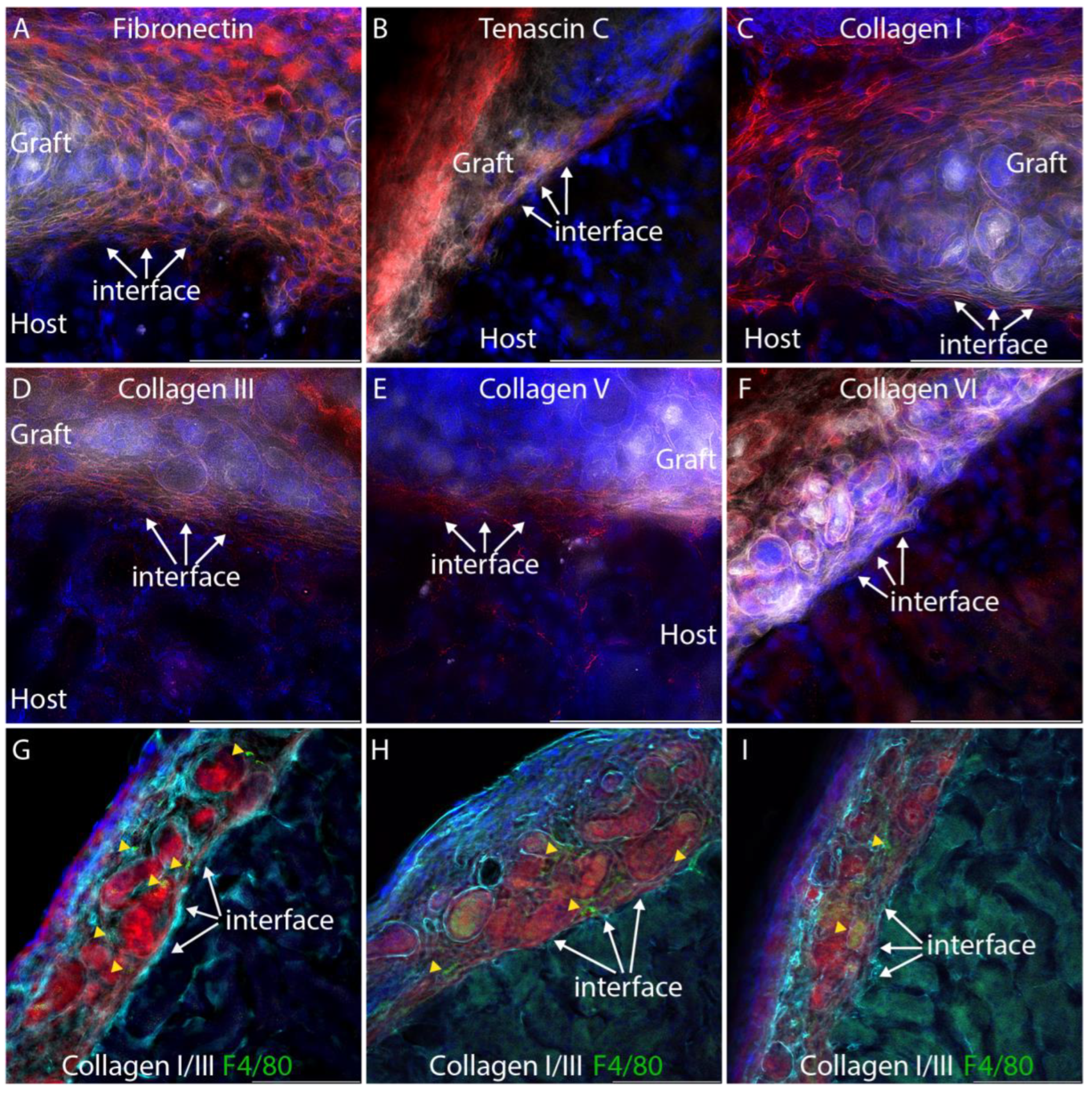
3.1. A Fibrin-Based Vehicle for Subcapsular Engraftment
- Thaw fibrinogen and thrombin stocks on ice.
- On ice, mix desired components (e.g., feeder cell suspension, recombinant proteins, virus), and stock fibrinogen and PBS (no Ca or Mg) to a final concentration of 33 mg/mL fibrinogen. Make 8 µL per clot and reserve on ice.
- Pipette 0.8 µL thrombin into one Eppendorf tube for each clot to be made and place on ice.
- Place organoids in 8 µL droplets of PBS on the surface of a Petri dish, one droplet per clot separated by at least 2 cm—it is important to make as small a footprint of PBS as possible with the droplet because it will determine the final diameter of the fibrin clot.
- From this point on, make the clots one at a time.
- Remove PBS from one droplet containing organoids. Make sure to drain as thoroughly as possible. Otherwise, the concentration of fibrinogen will be inaccurate.
- Withdraw 7.2 µL fibrinogen mix using a Gilson P10 or P20, and while the liquid is still in the tip, adjust the volume of the pipette to 8 µL.
- Depress the plunger enough to expel the air and pipette the fibrinogen solution into one Eppendorf tube containing thrombin.
- Rapidly aspirate and expel three times to mix the components. Make sure you do not introduce air bubbles.
- Immediately pipette the solution on top of the organoids without increasing the size of the footprint created by the PBS droplet.
- After all clots have been pipetted, transfer them to a 37 °C humidified incubator.
- After 30 min at 37 °C, keep on ice for 30 min.
- For surgery, it is most convenient to flood the dish with serum-free medium and keep the plate on ice. During surgery, detach the clots from the plate immediately before implantation by cutting with a scalpel as close to the dish surface as possible and implant under the kidney capsule with the side that was attached to the dish facing the cortex. This ensures that the organoids are in direct contact with the kidney cortex.
- Alternatively, the clots can all be detached after curing and placed in a tube or Petri dish containing a few mL of serum-free medium on ice awaiting surgery—this is most convenient if clots are to be transported, but the disadvantage is that it is difficult to distinguish the two different sides of the clot when doing the surgery.
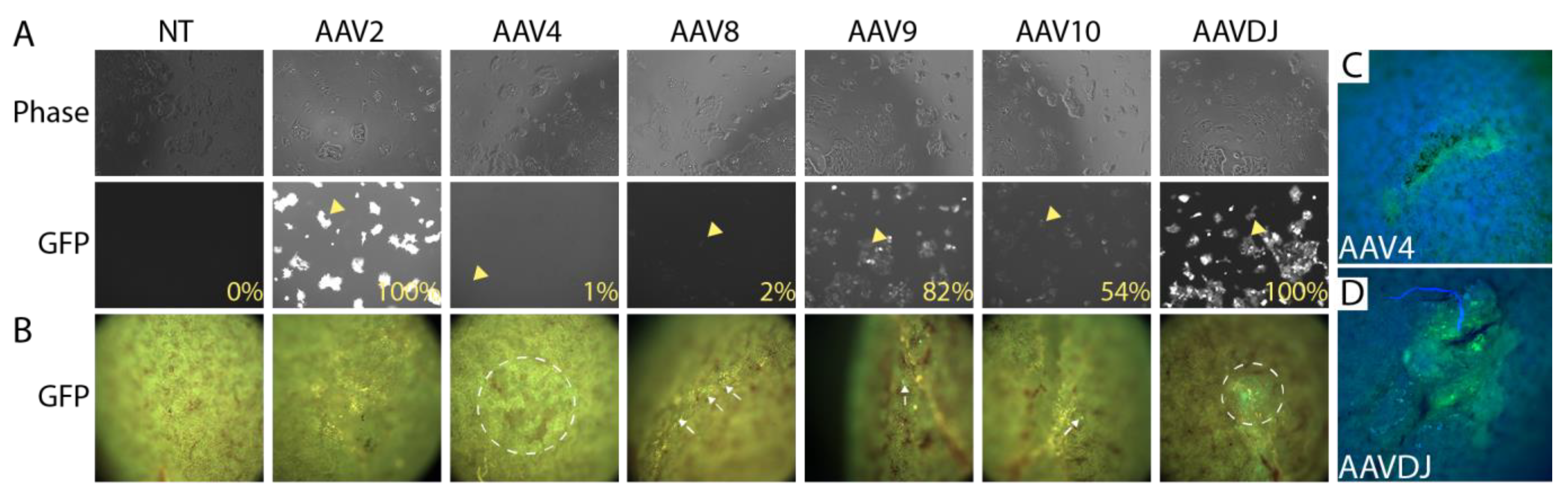
3.2. Selection of a Gene Therapy Vector for Local Delivery in Fibrin Hydrogel
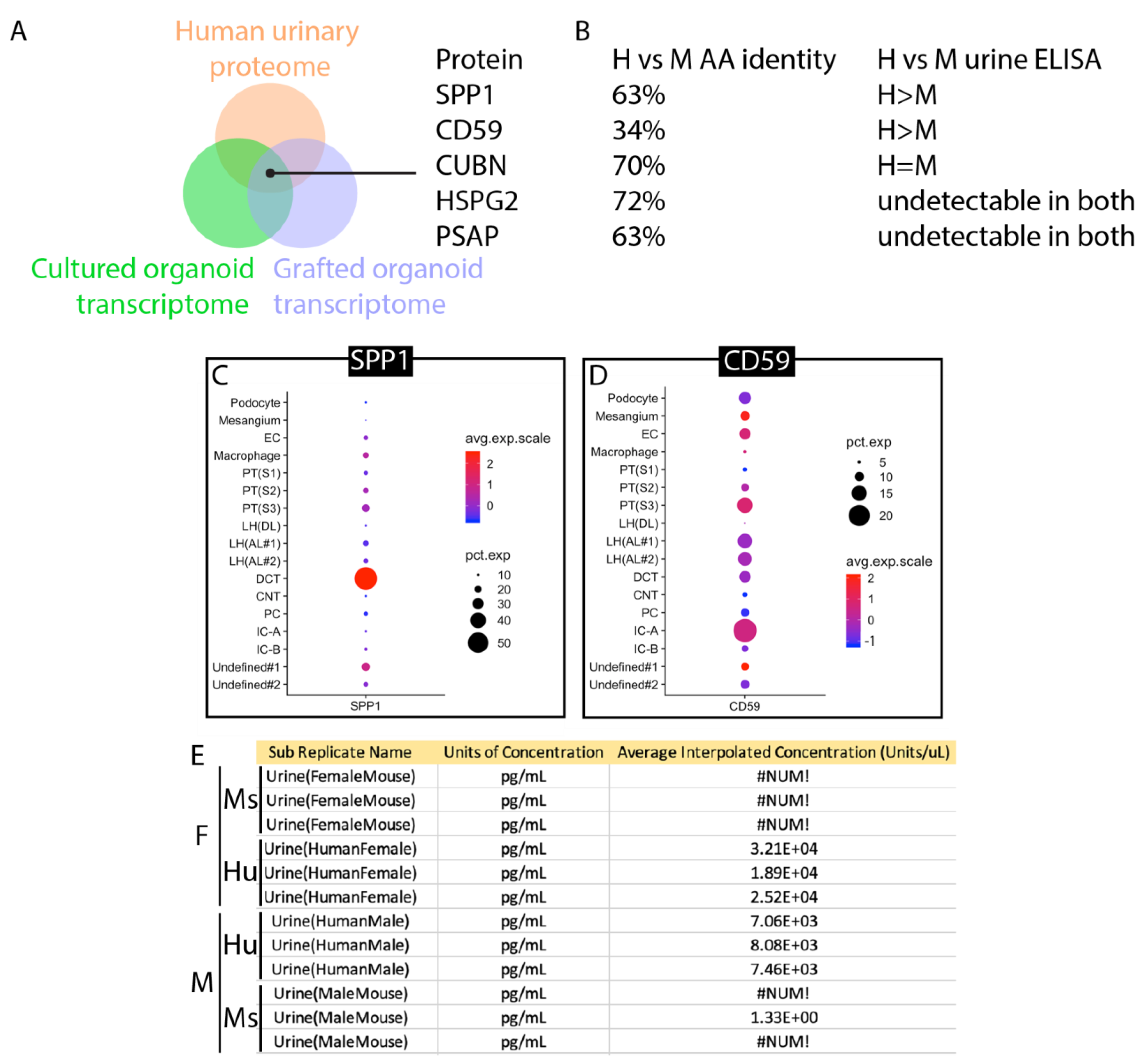
3.3. Identification of Human-Specific Urinary Biomarkers That Can Be Detected in Mouse Urine
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoerger, T.J.; Simpson, S.A.; Yarnoff, B.O.; Pavkov, M.E.; Ríos Burrows, N.; Saydah, S.H.; Williams, D.E.; Zhuo, X. The Future Burden of CKD in the United States: A Simulation Model for the CDC CKD Initiative. Am. J. Kidney Dis. 2015, 65, 403–411. [Google Scholar] [CrossRef]
- United States Renal Data System. 2021 USRDS Annual Data Report: Epidemiology of Kidney Disease in the United States; USRDS: Minneapolis, MN, USA, 2021.
- Kaballo, M.A.; Canney, M.; O’Kelly, P.; Williams, Y.; O’Seaghdha, C.M.; Conlon, P.J. A Comparative Analysis of Survival of Patients on Dialysis and after Kidney Transplantation. Clin. Kidney J. 2018, 11, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Maung, S.; Sara, A.E.; Cohen, D.; Chapman, C.; Saggi, S.; Cukor, D. Sleep Disturbance and Depressive Affect in Patients Treated with Haemodialysis. J. Ren. Care 2017, 43, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Takasato, M.; Er, P.X.; Chiu, H.S.; Maier, B.; Baillie, G.J.; Ferguson, C.; Parton, R.G.; Wolvetang, E.J.; Roost, M.S.; Chuva de Sousa Lopes, S.M.; et al. Kidney Organoids from Human iPS Cells Contain Multiple Lineages and Model Human Nephrogenesis. Nature 2015, 526, 564–568. [Google Scholar] [CrossRef]
- Taguchi, A.; Kaku, Y.; Ohmori, T.; Sharmin, S.; Ogawa, M.; Sasaki, H.; Nishinakamura, R. Redefining the in Vivo Origin of Metanephric Nephron Progenitors Enables Generation of Complex Kidney Structures from Pluripotent Stem Cells. Cell Stem Cell 2014, 14, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Morizane, R.; Lam, A.Q.; Freedman, B.S.; Kishi, S.; Valerius, M.T.; Bonventre, J.V. Nephron Organoids Derived from Human Pluripotent Stem Cells Model Kidney Development and Injury. Nat. Biotechnol. 2015, 33, 1193–1200. [Google Scholar] [CrossRef]
- Oxburgh, L.; Carroll, T.J.; Cleaver, O.; Gossett, D.R.; Hoshizaki, D.K.; Hubbell, J.A.; Humphreys, B.D.; Jain, S.; Jensen, J.; Kaplan, D.L.; et al. (Re)Building a Kidney. J. Am. Soc. Nephrol. 2017, 28, 1370–1378. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Nishinakamura, R. Building Kidney Organoids from Pluripotent Stem Cells. Curr. Opin. Nephrol. Hypertens. 2022, 31, 367–373. [Google Scholar] [CrossRef]
- Kumar Gupta, A.; Sarkar, P.; Wertheim, J.A.; Pan, X.; Carroll, T.J.; Oxburgh, L. Asynchronous Mixing of Kidney Progenitor Cells Potentiates Nephrogenesis in Organoids. Commun. Biol. 2020, 3, 231. [Google Scholar] [CrossRef]
- van den Berg, C.W.; Ritsma, L.; Avramut, M.C.; Wiersma, L.E.; van den Berg, B.M.; Leuning, D.G.; Lievers, E.; Koning, M.; Vanslambrouck, J.M.; Koster, A.J.; et al. Renal Subcapsular Transplantation of PSC-Derived Kidney Organoids Induces Neo-Vasculogenesis and Significant Glomerular and Tubular Maturation In Vivo. Stem Cell Rep. 2018, 10, 751–765. [Google Scholar] [CrossRef]
- Ryan, A.R.; England, A.R.; Chaney, C.P.; Cowdin, M.A.; Hiltabidle, M.; Daniel, E.; Gupta, A.K.; Oxburgh, L.; Carroll, T.J.; Cleaver, O. Vascular Deficiencies in Renal Organoids and Ex Vivo Kidney Organogenesis. Dev. Biol. 2021, 477, 98–116. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, C.W.; Koudijs, A.; Ritsma, L.; Rabelink, T.J. In Vivo Assessment of Size-Selective Glomerular Sieving in Transplanted Human Induced Pluripotent Stem Cell-Derived Kidney Organoids. J. Am. Soc. Nephrol. 2020, 31, 921–929. [Google Scholar] [CrossRef]
- Brown, A.C.; Muthukrishnan, S.D.; Oxburgh, L. A Synthetic Niche for Nephron Progenitor Cells. Dev. Cell 2015, 34, 229–241. [Google Scholar] [CrossRef]
- Shultz, L.D.; Goodwin, N.; Ishikawa, F.; Hosur, V.; Lyons, B.L.; Greiner, D.L. Subcapsular Transplantation of Tissue in the Kidney. Cold Spring Harb. Protoc. 2014, 2014, 737–740. [Google Scholar] [CrossRef] [PubMed][Green Version]
- van Hinsbergh, V.W.; Collen, A.; Koolwijk, P. Role of Fibrin Matrix in Angiogenesis. Ann. N. Y. Acad. Sci. 2001, 936, 426–437. [Google Scholar] [CrossRef]
- Kamei, C.N.; Gallegos, T.F.; Liu, Y.; Hukriede, N.; Drummond, I.A. Wnt Signaling Mediates New Nephron Formation during Zebrafish Kidney Regeneration. Development 2019, 146, dev168294. [Google Scholar] [CrossRef] [PubMed]
- Lipp, S.N.; Jacobson, K.R.; Hains, D.S.; Schwarderer, A.L.; Calve, S. 3D Mapping Reveals a Complex and Transient Interstitial Matrix During Murine Kidney Development. J. Am. Soc. Nephrol. 2021, 32, 1649–1665. [Google Scholar] [CrossRef]
- Bond, K.H.; Chiba, T.; Wynne, K.P.H.; Vary, C.P.H.; Sims-Lucas, S.; Coburn, J.M.; Oxburgh, L. The Extracellular Matrix Environment of Clear Cell Renal Cell Carcinoma Determines Cancer Associated Fibroblast Growth. Cancers 2021, 13, 5873. [Google Scholar] [CrossRef]
- Capuani, S.; Malgir, G.; Chua, C.Y.X.; Grattoni, A. Advanced Strategies to Thwart Foreign Body Response to Implantable Devices. Bioeng. Transl. Med. 2022, 7, e10300. [Google Scholar] [CrossRef]
- Shultz, L.D.; Schweitzer, P.A.; Christianson, S.W.; Gott, B.; Schweitzer, I.B.; Tennent, B.; McKenna, S.; Mobraaten, L.; Rajan, T.V.; Greiner, D.L. Multiple Defects in Innate and Adaptive Immunologic Function in NOD/LtSz-Scid Mice. J. Immunol. Baltim. Md. 1950 1995, 154, 180–191. [Google Scholar] [CrossRef]
- Issa, S.S.; Shaimardanova, A.A.; Solovyeva, V.V.; Rizvanov, A.A. Various AAV Serotypes and Their Applications in Gene Therapy: An Overview. Cells 2023, 12, 785. [Google Scholar] [CrossRef]
- Srivastava, A. In Vivo Tissue-Tropism of Adeno-Associated Viral Vectors. Curr. Opin. Virol. 2016, 21, 75–80. [Google Scholar] [CrossRef]
- Ito, K.; Chen, J.; Khodadadian, J.J.; Vaughan, E.D.; Lipkowitz, M.; Poppas, D.P.; Felsen, D. Adeno-Associated Viral Vector Transduction of Green Fluorescent Protein in Kidney: Effect of Unilateral Ureteric Obstruction. BJU Int. 2008, 101, 376–381. [Google Scholar] [CrossRef]
- Chung, D.C.; Fogelgren, B.; Park, K.M.; Heidenberg, J.; Zuo, X.; Huang, L.; Bennett, J.; Lipschutz, J.H. Adeno-Associated Virus-Mediated Gene Transfer to Renal Tubule Cells via a Retrograde Ureteral Approach. Nephron Extra 2011, 1, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Lee, J.S.; Wang, L.; Desai, T.; Akache, B.; Storm, T.A.; Kay, M.A. In Vitro and in Vivo Gene Therapy Vector Evolution via Multispecies Interbreeding and Retargeting of Adeno-Associated Viruses. J. Virol. 2008, 82, 5887–5911. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.K.; Majumdar, S.S.; Alam, P.; Gulati, N.; Brahmachari, V. Epigenetic Regulation of Cytomegalovirus Major Immediate-Early Promoter Activity in Transgenic Mice. Gene 2009, 428, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.Y.; Zhang, L.; Clift, K.L.; Hulur, I.; Xiang, A.P.; Ren, B.-Z.; Lahn, B.T. Systematic Comparison of Constitutive Promoters and the Doxycycline-Inducible Promoter. PLoS ONE 2010, 5, e10611. [Google Scholar] [CrossRef] [PubMed]
- LaFavers, K.; Garimella, P.S. Uromodulin: More than a Marker for Chronic Kidney Disease Progression. Curr. Opin. Nephrol. Hypertens. 2023, 32, 271–277. [Google Scholar] [CrossRef]
- Stricklett, P.K.; Taylor, D.; Nelson, R.D.; Kohan, D.E. Thick Ascending Limb-Specific Expression of Cre Recombinase. Am. J. Physiol. Renal Physiol. 2003, 285, F33–F39. [Google Scholar] [CrossRef] [PubMed]
- Belge, H.; Gailly, P.; Schwaller, B.; Loffing, J.; Debaix, H.; Riveira-Munoz, E.; Beauwens, R.; Devogelaer, J.-P.; Hoenderop, J.G.; Bindels, R.J.; et al. Renal Expression of Parvalbumin Is Critical for NaCl Handling and Response to Diuretics. Proc. Natl. Acad. Sci. USA 2007, 104, 14849–14854. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.-H.; Leong, W.-S.; Ismail, Z.; Gam, L.-H. Qualification and Application of an ELISA for the Determination of Tamm Horsfall Protein (THP) in Human Urine and Its Use for Screening of Kidney Stone Disease. Int. J. Biol. Sci. 2008, 4, 215–222. [Google Scholar] [CrossRef]
- Zhao, M.; Li, M.; Yang, Y.; Guo, Z.; Sun, Y.; Shao, C.; Li, M.; Sun, W.; Gao, Y. A Comprehensive Analysis and Annotation of Human Normal Urinary Proteome. Sci. Rep. 2017, 7, 3024. [Google Scholar] [CrossRef] [PubMed]
- Phipson, B.; Er, P.X.; Combes, A.N.; Forbes, T.A.; Howden, S.E.; Zappia, L.; Yen, H.-J.; Lawlor, K.T.; Hale, L.J.; Sun, J.; et al. Evaluation of Variability in Human Kidney Organoids. Nat. Methods 2019, 16, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Sidhom, E.-H.; Emani, M.; Vernon, K.; Sahakian, N.; Zhou, Y.; Kost-Alimova, M.; Slyper, M.; Waldman, J.; Dionne, D.; et al. Single Cell Census of Human Kidney Organoids Shows Reproducibility and Diminished Off-Target Cells after Transplantation. Nat. Commun. 2019, 10, 5462. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Malone, A.F.; Donnelly, E.L.; Kirita, Y.; Uchimura, K.; Ramakrishnan, S.M.; Gaut, J.P.; Humphreys, B.D. Single-Cell Transcriptomics of a Human Kidney Allograft Biopsy Specimen Defines a Diverse Inflammatory Response. J. Am. Soc. Nephrol. 2018, 29, 2069–2080. [Google Scholar] [CrossRef]
- Rooney, I.A.; Davies, A.; Griffiths, D.; Williams, J.D.; Davies, M.; Meri, S.; Lachmann, P.J.; Morgan, B.P. The Complement-Inhibiting Protein, Protectin (CD59 Antigen), Is Present and Functionally Active on Glomerular Epithelial Cells. Clin. Exp. Immunol. 1991, 83, 251–256. [Google Scholar] [CrossRef]
- Ichida, S.; Yuzawa, Y.; Okada, H.; Yoshioka, K.; Matsuo, S. Localization of the Complement Regulatory Proteins in the Normal Human Kidney. Kidney Int. 1994, 46, 89–96. [Google Scholar] [CrossRef]
- Matsuo, S.; Nishikage, H.; Yoshida, F.; Nomura, A.; Piddlesden, S.J.; Morgan, B.P. Role of CD59 in Experimental Glomerulonephritis in Rats. Kidney Int. 1994, 46, 191–200. [Google Scholar] [CrossRef][Green Version]
- Ren, S.; Duffield, J.S. Pericytes in Kidney Fibrosis. Curr. Opin. Nephrol. Hypertens. 2013, 22, 471–480. [Google Scholar] [CrossRef]
- Kuraoka, S.; Tanigawa, S.; Taguchi, A.; Hotta, A.; Nakazato, H.; Osafune, K.; Kobayashi, A.; Nishinakamura, R. PKD1-Dependent Renal Cystogenesis in Human Induced Pluripotent Stem Cell-Derived Ureteric Bud/Collecting Duct Organoids. J. Am. Soc. Nephrol. 2020, 31, 2355–2371. [Google Scholar] [CrossRef]
- Shi, M.; McCracken, K.W.; Patel, A.B.; Zhang, W.; Ester, L.; Valerius, M.T.; Bonventre, J.V. Human Ureteric Bud Organoids Recapitulate Branching Morphogenesis and Differentiate into Functional Collecting Duct Cell Types. Nat. Biotechnol. 2023, 41, 252–261. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, T.; Bejar, J.; Yue, Z.; Slavinsky, M.; Marciano, D.; Drummond, I.; Oxburgh, L. In Vivo Assessment of Laboratory-Grown Kidney Tissue Grafts. Bioengineering 2023, 10, 1261. https://doi.org/10.3390/bioengineering10111261
Chuang T, Bejar J, Yue Z, Slavinsky M, Marciano D, Drummond I, Oxburgh L. In Vivo Assessment of Laboratory-Grown Kidney Tissue Grafts. Bioengineering. 2023; 10(11):1261. https://doi.org/10.3390/bioengineering10111261
Chicago/Turabian StyleChuang, Tinghsien, Justin Bejar, Zhiwei Yue, Mary Slavinsky, Denise Marciano, Iain Drummond, and Leif Oxburgh. 2023. "In Vivo Assessment of Laboratory-Grown Kidney Tissue Grafts" Bioengineering 10, no. 11: 1261. https://doi.org/10.3390/bioengineering10111261
APA StyleChuang, T., Bejar, J., Yue, Z., Slavinsky, M., Marciano, D., Drummond, I., & Oxburgh, L. (2023). In Vivo Assessment of Laboratory-Grown Kidney Tissue Grafts. Bioengineering, 10(11), 1261. https://doi.org/10.3390/bioengineering10111261