Eye in the Sky: Using UAV Imagery of Seasonal Riverine Canopy Growth to Model Water Temperature


Abstract
:1. Introduction
2. Materials and Methods
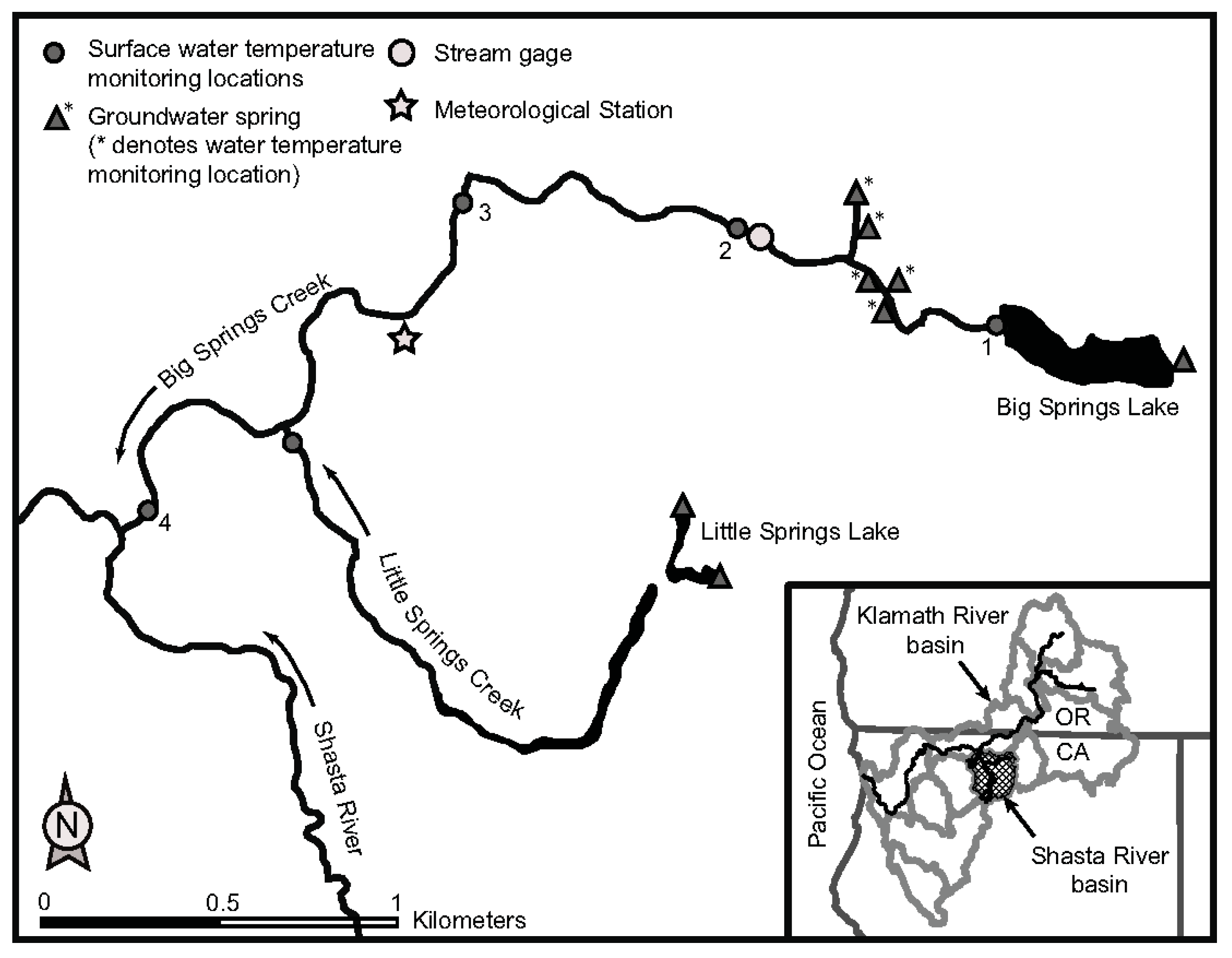
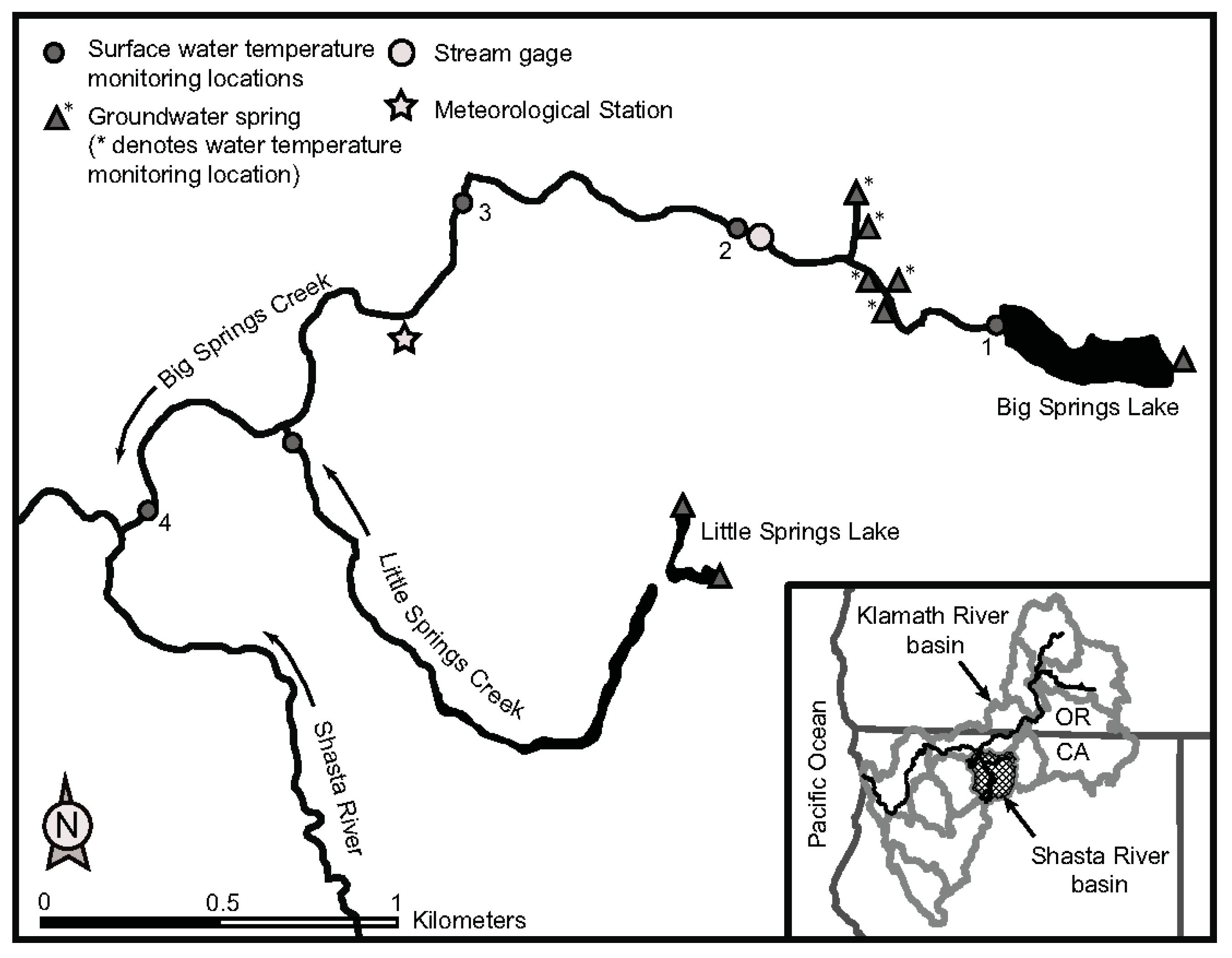
2.1. Study Site and Period
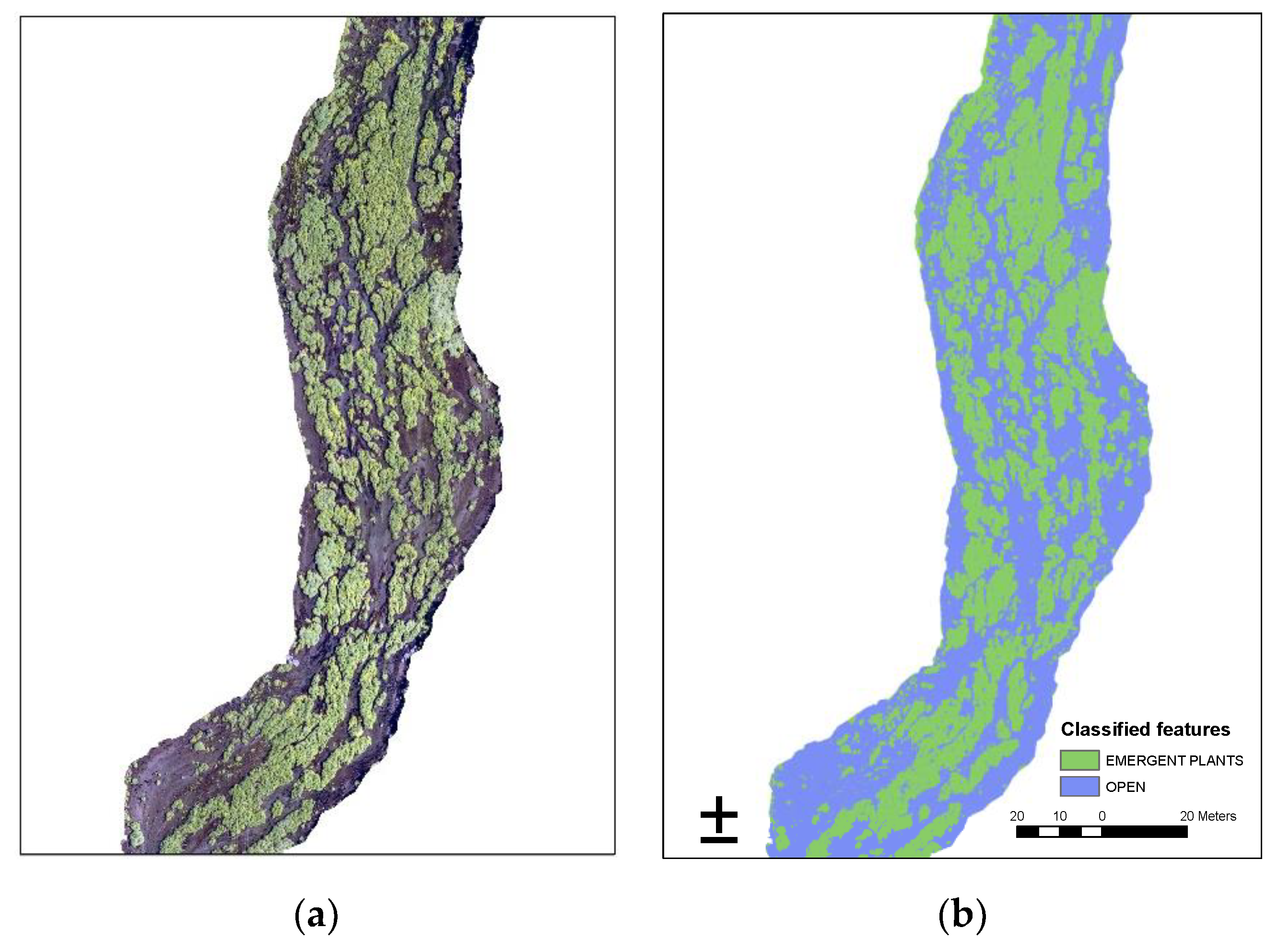
2.2. Riverine Canopy Surveys
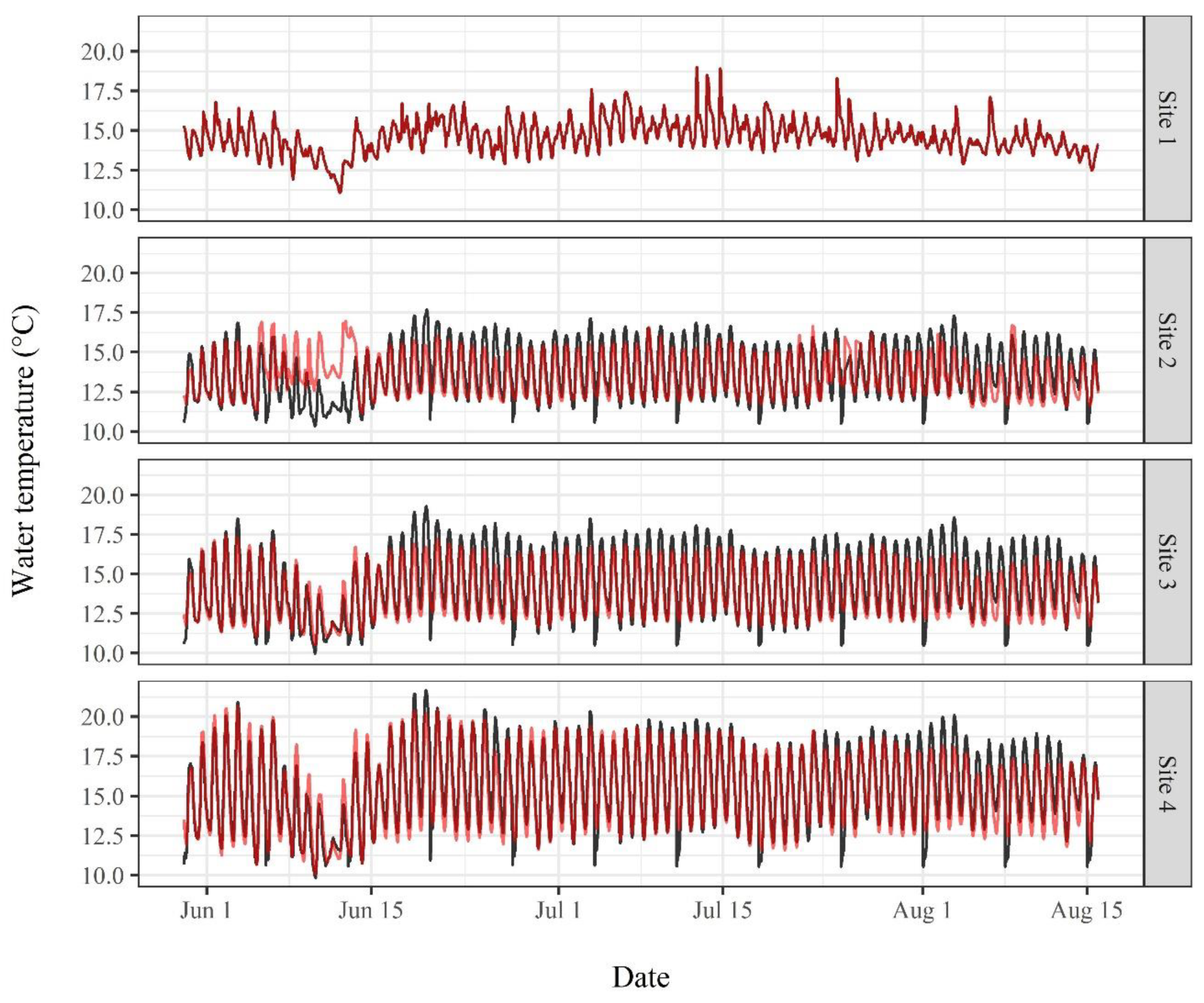
2.3. Water Temperature Modeling
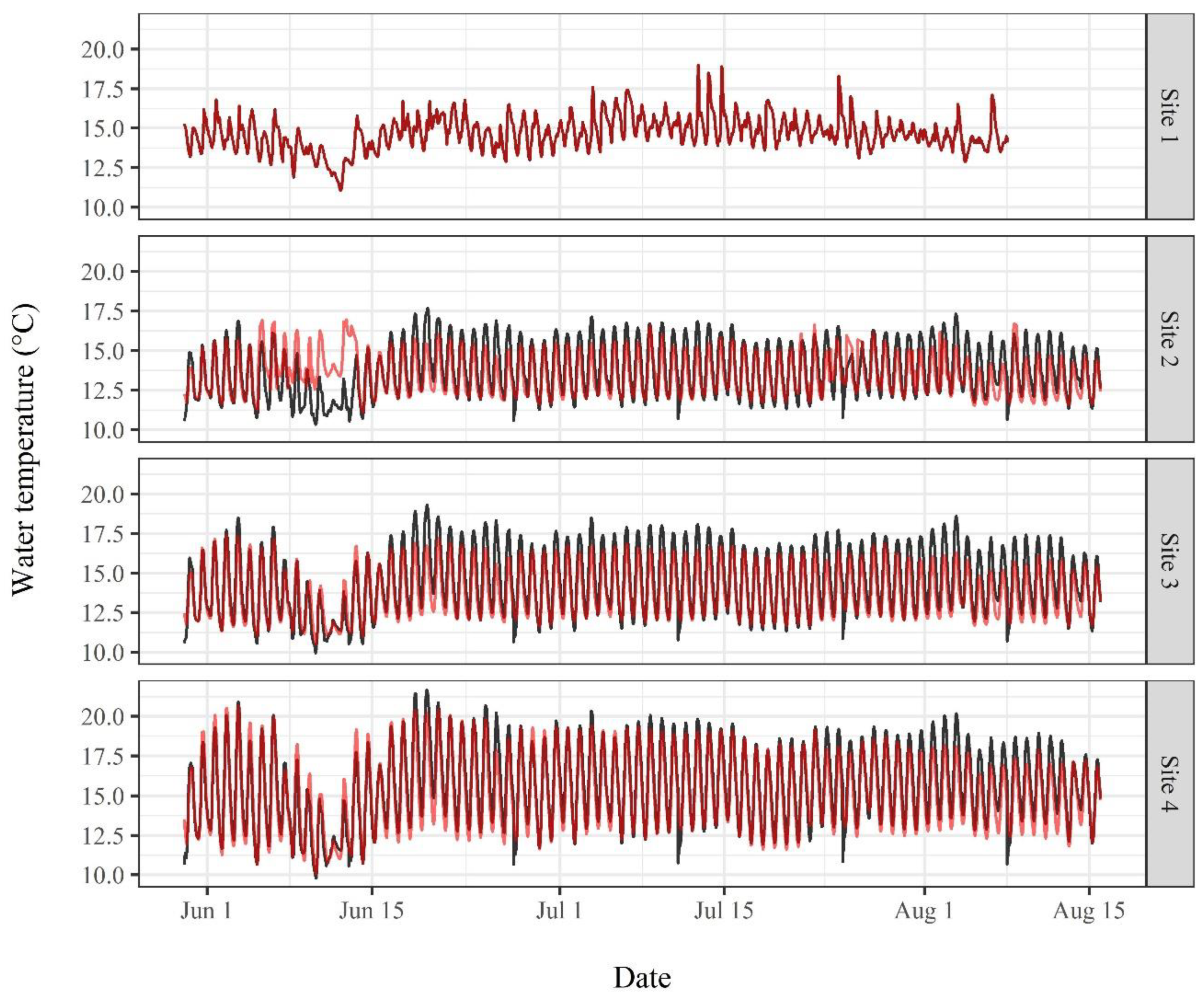
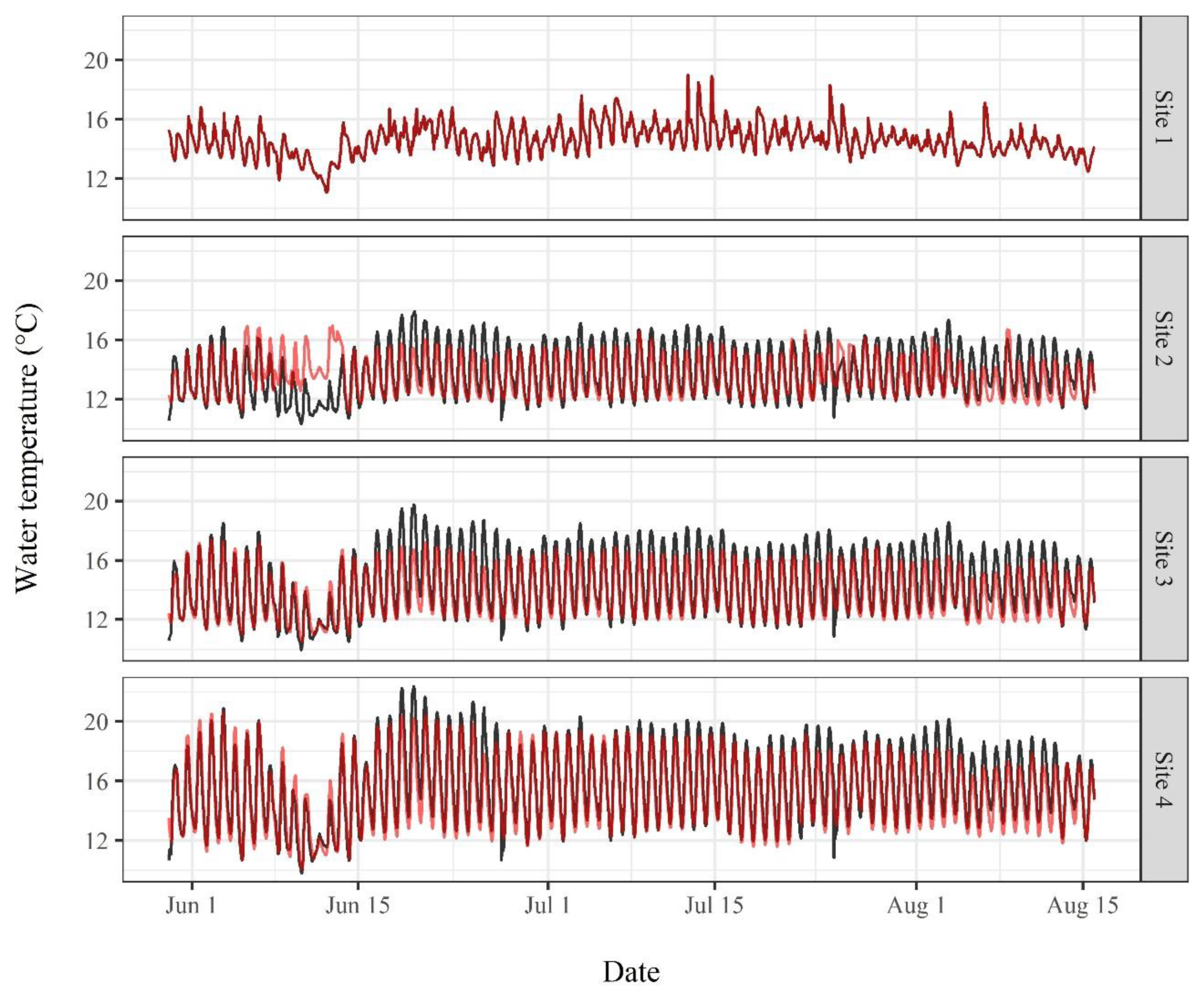
3. Results
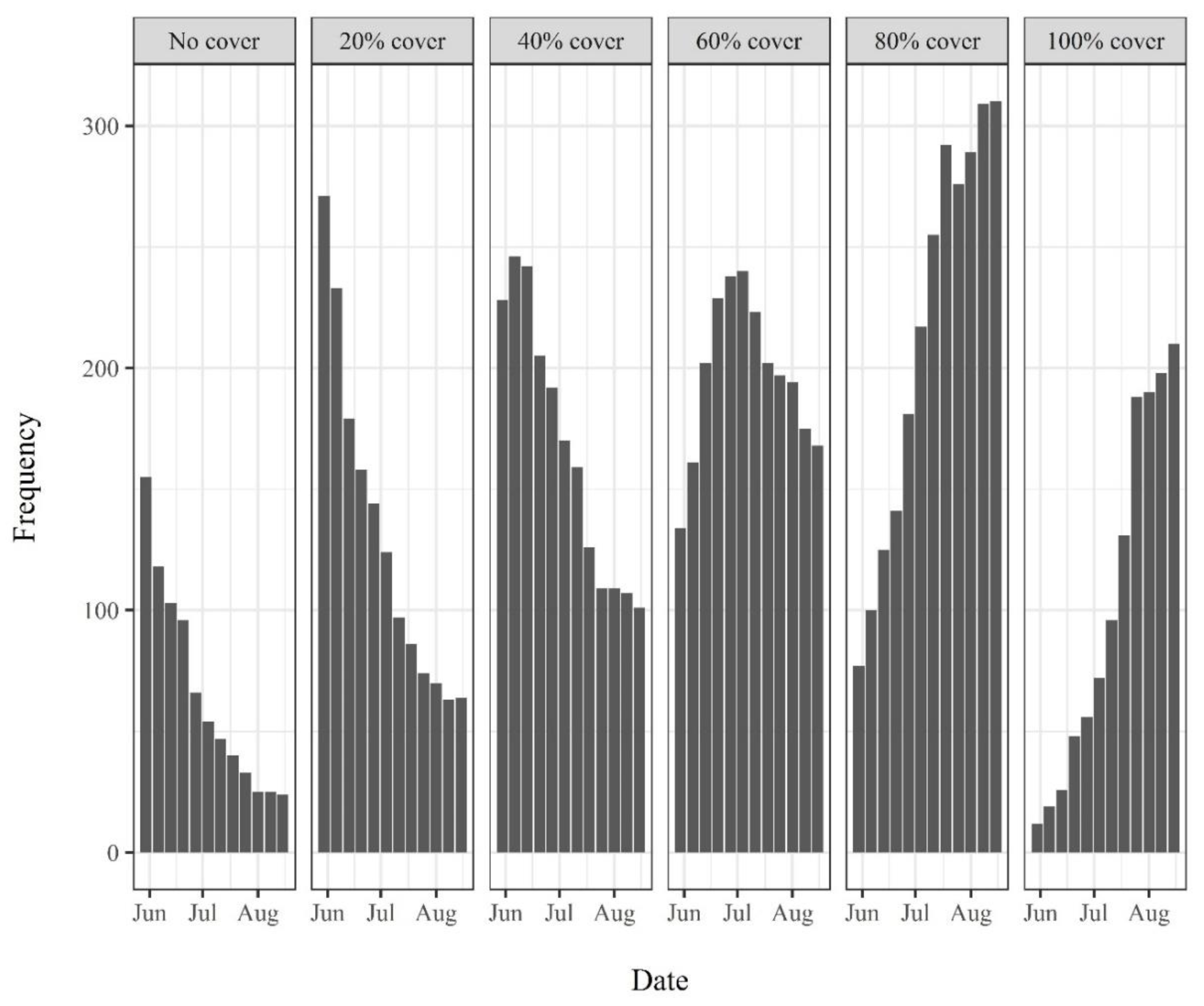
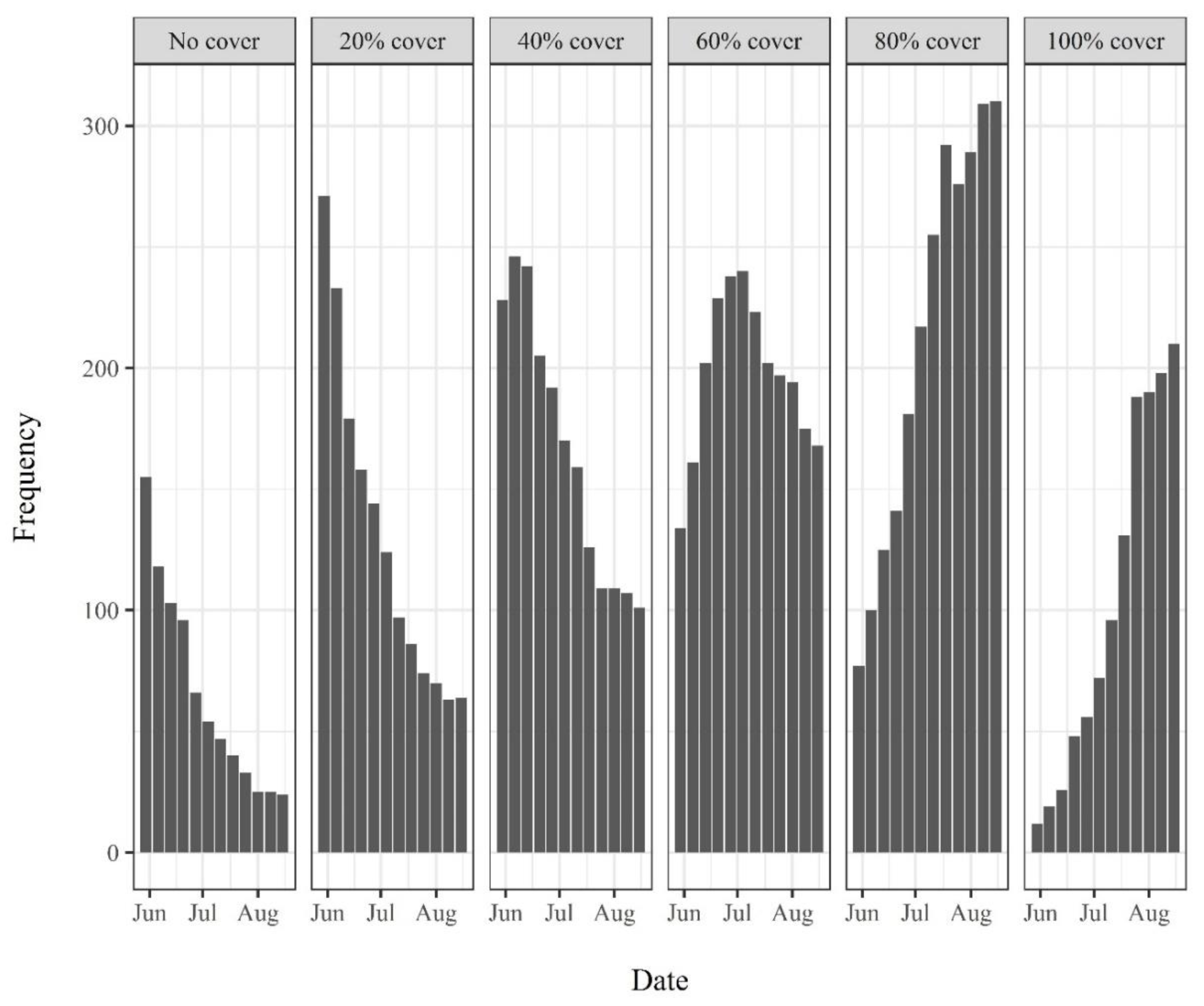
3.1. Riverine Canopy Surveys
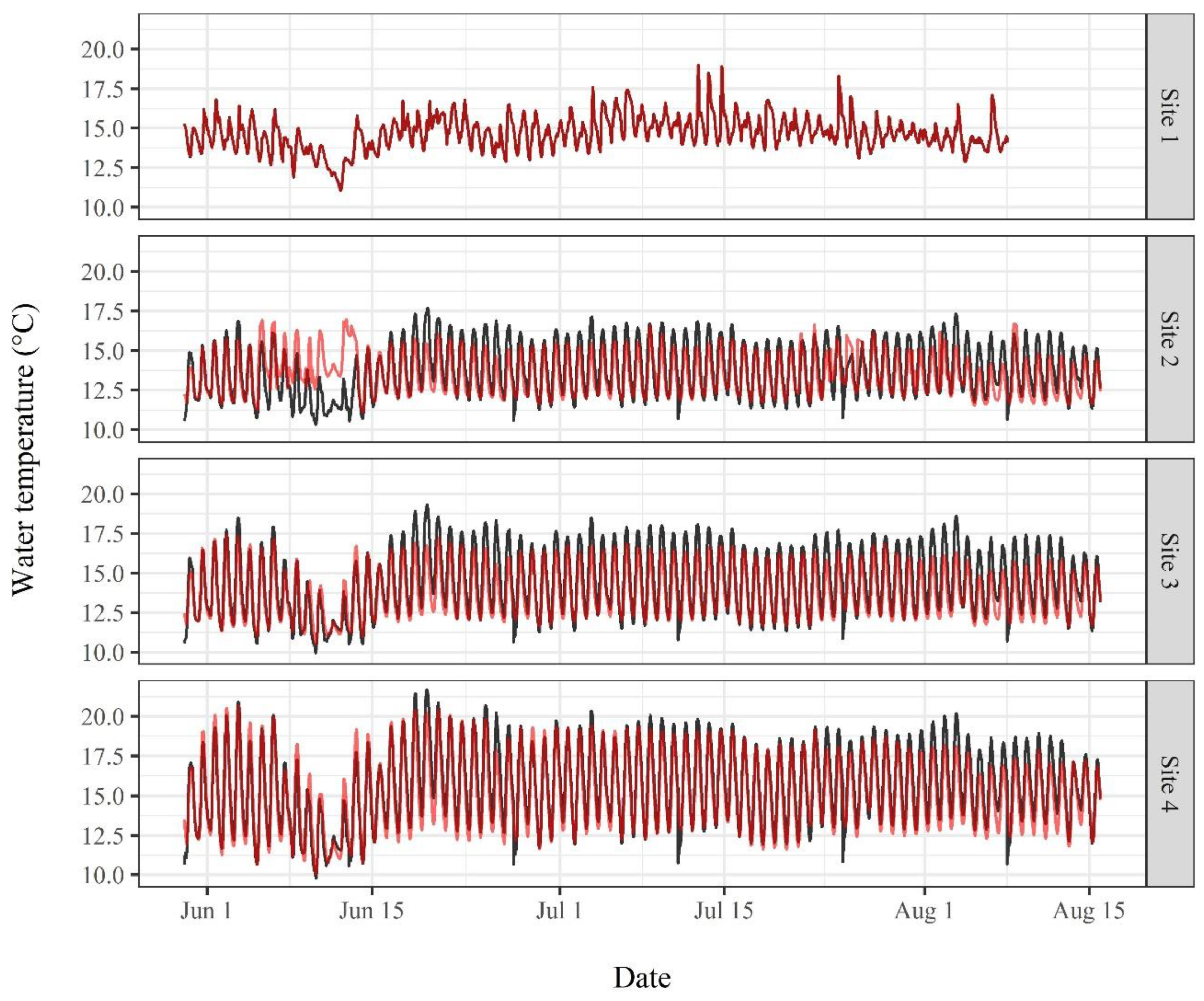
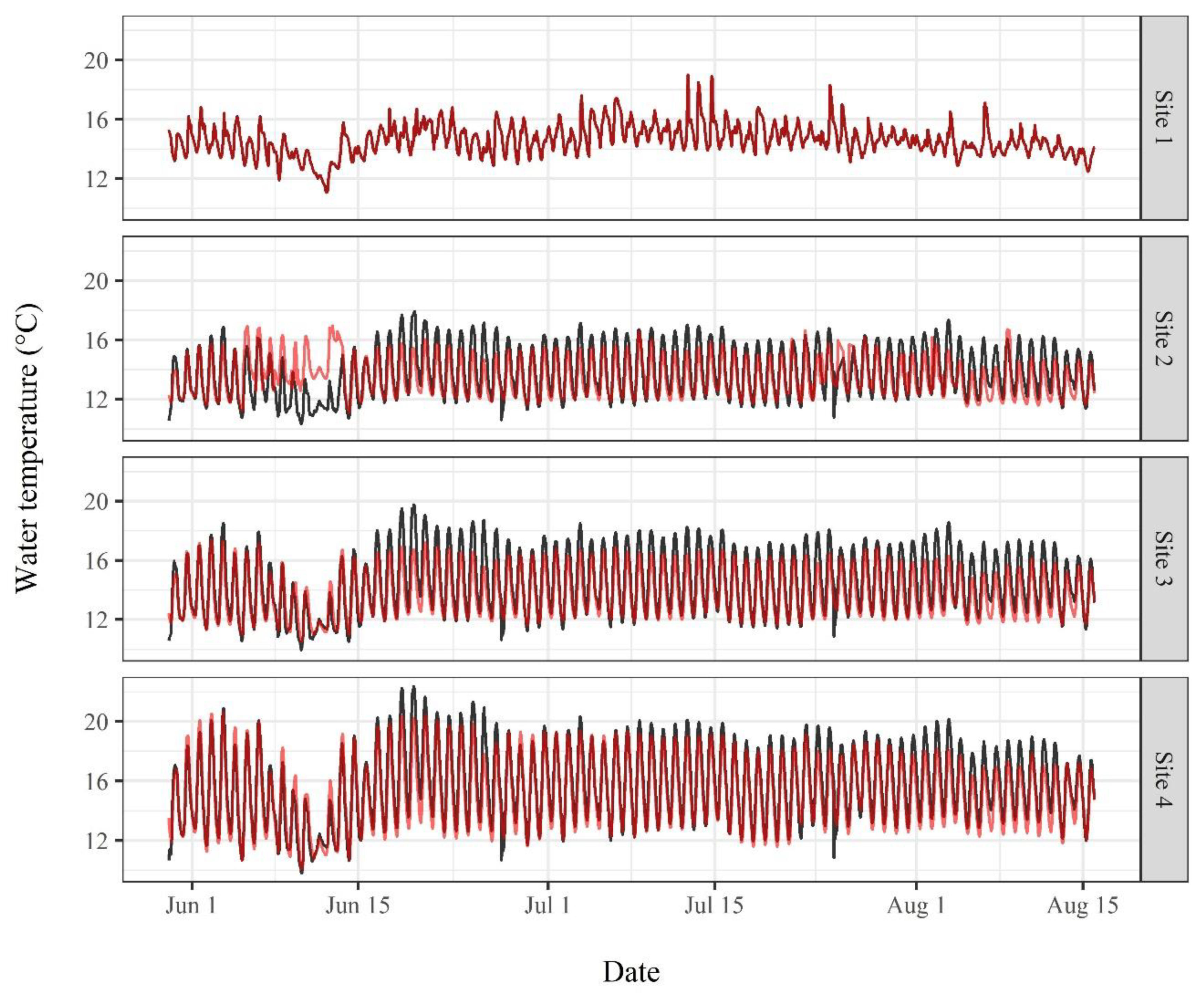
3.2. Water Temperature Modeling
4. Discussion
4.1. UAV Survey Methods
4.2. Riverine Canopy Growth
4.3. Water Temperature Modeling
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Caissie, D. The thermal regime of rivers: A review. Freshw. Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Steel, E.A.; Beechie, T.J.; Torgersen, C.E.; Fullerton, A.H. Envisioning, quantifying, and managing thermal regimes on river networks. BioScience 2017, 67, 506–522. [Google Scholar] [CrossRef]
- Webb, B.W.; Hannah, D.M.; Moore, R.D.; Brown, L.E.; Nobilis, F. Recent advances in stream and river temperature research. Hydrol. Process. 2008, 22, 902–918. [Google Scholar] [CrossRef]
- Davis, J.; Baxter, C.; Rosi-Marshall, E.; Pierce, J.; Crosby, B. Anticipating stream ecosystem responses to climate change: Toward predictions that incorporate effects via land–water linkages. Ecosystems 2013, 16, 909–922. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2093–2106. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.P.; Peckarsky, B.L. Emergence cues of a mayfly in a high-altitude stream ecosystem: Potential response to climate change. Ecol. Appl. 2006, 16, 612–621. [Google Scholar] [CrossRef]
- Isaak, D.J.; Wenger, S.J.; Young, M.K. Big biology meets microclimatology: Defining thermal niches of ectotherms at landscape scales for conservation planning. Ecol. Appl. 2017, 27, 977–990. [Google Scholar] [CrossRef]
- Isaak, D.J.; Wenger, S.J.; Peterson, E.E.; Ver Hoef, J.M.; Nagel, D.E.; Luce, C.H.; Hostetler, S.W.; Dunham, J.B.; Roper, B.B.; Wollrab, S.P. The NorWEST summer stream temperature model and scenarios for the western U.S.: A crowd-sourced database and new geospatial tools foster a user community and predict broad climate warming of rivers and streams. Water Resour. Res. 2017, 53, 9181–9205. [Google Scholar] [CrossRef]
- Dugdale, S.J.; Hannah, D.M.; Malcolm, I.A. River temperature modelling: A review of process-based approaches and future directions. Earth Sci. Rev. 2017, 175, 87–113. [Google Scholar] [CrossRef]
- Hannah, D.M.; Garner, G. River water temperature in the United Kingdom: Changes over the 20th century and possible changes over the 21st century. Prog. Phys. Geogr. 2015, 39, 68–92. [Google Scholar] [CrossRef] [Green Version]
- Webb, B. Trends in stream and river temperature. Hydrol. Process. 1996, 10, 205–226. [Google Scholar] [CrossRef]
- Moore, R.D. Stream temperature patterns in British Columbia, Canada, based on routine spot measurements. Can. Water Resour. J. 2006, 31, 41–56. [Google Scholar] [CrossRef]
- Johnson, S.L. Factors influencing stream temperatures in small streams: Substrate effects and a shading experiment. Can. J. Fish. Aquat. Sci. 2004, 61, 913–923. [Google Scholar] [CrossRef]
- Garner, G.; Malcolm, I.A.; Sadler, J.P.; Hannah, D.M. The role of riparian vegetation density, channel orientation and water velocity in determining river temperature dynamics. J. Hydrol. 2017, 553, 471–485. [Google Scholar] [CrossRef]
- Moore, R.; Spittlehouse, D.; Story, A. Riparian microclimate and stream temperature response to forest harvesting: A review 1. JAWRA J. Am. Water Resour. Assoc. 2005, 41, 813–834. [Google Scholar] [CrossRef]
- Willis, A.D.; Nichols, A.L.; Holmes, E.J.; Jeffres, C.A.; Fowler, A.C.; Babcock, C.A.; Deas, M.L. Seasonal aquatic macrophytes reduce water temperatures via a riverine canopy in a spring-fed stream. Freshw. Sci. 2017, 36, 508–522. [Google Scholar] [CrossRef]
- Kalny, G.; Laaha, G.; Melcher, A.; Trimmel, H.; Weihs, P.; Rauch, H.P. The influence of riparian vegetation shading on water temperature during low flow conditions in a medium sized river. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 5. [Google Scholar] [CrossRef]
- Rutherford, C.J.; Meleason, M.A.; Davies-Colley, R.J. Modelling stream shade: 2. Predicting the effects of canopy shape and changes over time. Ecol. Eng. 2018, 120, 487–496. [Google Scholar] [CrossRef]
- Sinokrot, B.A.; Stefan, H.G. Stream temperature dynamics: Measurements and modeling. Water Resour. Res. 1993, 29, 2299–2312. [Google Scholar] [CrossRef]
- Benyahya, L.; Caissie, D.; St-Hilaire, A.; Ouarda, T.B.; Bobée, B. A review of statistical water temperature models. Can. Water Resour. J. 2007, 32, 179–192. [Google Scholar] [CrossRef]
- Schabenberger, O.; Gotway, C.A. Statistical Methods for Spatial Data Analysis; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Trimmel, H.; Weihs, P.; Leidinger, D.; Formayer, H.; Kalny, G.; Melcher, A. Can riparian vegetation shade mitigate the expected rise in stream temperatures due to climate change during heat waves in a human-impacted pre-alpine river? Hydrol. Earth Syst. Sci. 2018, 22, 437–461. [Google Scholar] [CrossRef] [Green Version]
- Loicq, P.; Moatar, F.; Jullian, Y.; Dugdale, S.J.; Hannah, D.M. Improving representation of riparian vegetation shading in a regional stream temperature model using lidar data. Sci. Total Environ. 2018, 624, 480–490. [Google Scholar] [CrossRef] [PubMed]
- McGrath, E.; Neumann, N.; Nichol, C. A statistical model for managing water temperature in streams with anthropogenic influences. River Res. Appl. 2017, 33, 123–134. [Google Scholar] [CrossRef]
- Wondzell, S.M.; Diabat, M.; Haggerty, R. What matters most: Are future stream temperatures more sensitive to changing air temperatures, discharge, or riparian vegetation? JAWRA J. Am. Water Resour. Assoc. 2018. [Google Scholar] [CrossRef]
- Van Vliet, M.; Ludwig, F.; Zwolsman, J.; Weedon, G.; Kabat, P. Global river temperatures and sensitivity to atmospheric warming and changes in river flow. Water Resour. Res. 2011, 47. [Google Scholar] [CrossRef]
- Arora, R.; Tockner, K.; Venohr, M. Changing river temperatures in northern Germany: Trends and drivers of change. Hydrol. Process. 2016, 30, 3084–3096. [Google Scholar] [CrossRef]
- Ptak, M.; Choiński, A.; Kirviel, J. Long-term water temperature fluctuations in coastal rivers (southern Baltic) in Poland. Bull. Geogr. Phys. Geogr. Ser. 2016, 11, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Null, S.E.; Viers, J.H.; Deas, M.L.; Tanaka, S.K.; Mount, J.F. Stream temperature sensitivity to climate warming in California’s Sierra Nevada: Impacts to coldwater habitat. Clim. Chang. 2013, 116, 149–170. [Google Scholar] [CrossRef]
- Moyle, P.B.; Lusardi, R.A.; Samuel, P.; Katz, J. State of the Salmonids: Status of Ccalifornia’s Emblematic Fishes 2017; University of California: Davis, CA, USA, 2017; p. 579. [Google Scholar]
- Vasseur, D.A.; DeLong, J.P.; Gilbert, B.; Greig, H.S.; Harley, C.D.; McCann, K.S.; Savage, V.; Tunney, T.D.; O’Connor, M.I. Increased temperature variation poses a greater risk to species than climate warming. Proc. R. Soc. Lond. B Biol. Sci. 2014, 281, 20132612. [Google Scholar] [CrossRef]
- Null, S.E.; Deas, M.L.; Lund, J.R. Flow and water temperature simulation for habitat restoration in the Shasta River, California. River Res. Appl. 2010, 26, 663–681. [Google Scholar] [CrossRef]
- Willis, A.D.; Campbell, A.M.; Fowler, A.C.; Babcock, C.A.; Howard, J.K.; Deas, M.L.; Nichols, A.L. Instream flows: New tools to quantify water quality conditions for returning adult Chinook salmon. J. Water Resour. Plan. Manag. 2015, 04015056. [Google Scholar] [CrossRef]
- Nichols, A.L.; Willis, A.D.; Jeffres, C.A.; Deas, M.L. Water temperature patterns below large groundwater springs: Management implications for coho salmon in the Shasta River, California. River Res. Appl. 2014, 30, 442–455. [Google Scholar] [CrossRef]
- Lowney, C.L. Stream temperature variation in regulated rivers: Evidence for a spatial pattern in daily minimum and maximum magnitudes. Water Resour. Res. 2000, 36, 2947–2955. [Google Scholar] [CrossRef] [Green Version]
- Verschoren, V.; Schoelynck, J.; Buis, K.; Visser, F.; Meire, P.; Temmerman, S. Mapping the spatio-temporal distribution of key vegetation cover properties in lowland river reaches, using digital photography. Environ. Monit. Assess. 2017, 189, 294. [Google Scholar] [CrossRef] [PubMed]
- Clark, P.E.; Johnson, D.E.; Hardegree, S.P. A direct approach for quantifying stream shading. Rangel. Ecol. Manag. 2008, 61, 339–345. [Google Scholar] [CrossRef]
- Kelley, C.E.; Krueger, W.C. Canopy cover and shade determinations in riparian zones 1. JAWRA J. Am. Water Resour. Assoc. 2005, 41, 37–046. [Google Scholar] [CrossRef]
- Richter, A.; Kolmes, S.A. Maximum temperature limits for chinook, coho, and chum salmon, and steelhead trout in the Pacific Northwest. Rev. Fish. Sci. 2005, 13, 23–49. [Google Scholar] [CrossRef]
- Rutherford, J.C.; Macaskill, J.B.; Williams, B.L. Natural water temperature variations in the lower Waikato River, New Zealand. N. Z. J. Mar. Freshw. Res. 1993, 27, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.; Webb, B.; Ladle, M. Microthermal gradients and ecological implications in Dorset rivers. Hydrol. Process. 1999, 13, 423–438. [Google Scholar] [CrossRef]
- Leung, L.R.; Qian, Y.; Bian, X.; Washington, W.M.; Han, J.; Roads, J.O. Mid-century ensemble regional climate change scenarios for the western United States. Clim. Chang. 2004, 62, 75–113. [Google Scholar] [CrossRef]
- Fabris, L.; Malcolm, I.A.; Buddendorf, W.B.; Soulsby, C. Integrating process-based flow and temperature models to assess riparian forests and temperature amelioration in salmon streams. Hydrol. Process. 2018, 32, 776–791. [Google Scholar] [CrossRef]
- Bartholow, J. Estimating cumulative effects of clearcutting on stream temperatures. Rivers 2000, 7, 284–297. [Google Scholar]
- Bal, K.; Struyf, E.; Vereecken, H.; Viaene, P.; De Doncker, L.; de Deckere, E.; Mostaert, F.; Meire, P. How do macrophyte distribution patterns affect hydraulic resistances? Ecol. Eng. 2011, 37, 529–533. [Google Scholar] [CrossRef]
- O’Hare, J.; O’Hare, M.; Gurnell, A.; Dunbar, M.; Scarlett, P.; Laize, C. Physical constraints on the distribution of macrophytes linked with flow and sediment dynamics in british rivers. River Res. Appl. 2011, 27, 671–683. [Google Scholar] [CrossRef]
- Green, J.C. Velocity and turbulence distribution around lotic macrophytes. Aquat. Ecol. 2005, 39, 1–10. [Google Scholar] [CrossRef]
- Lacey, R.J.; Millar, R.G. Reach scale hydraulic assessment of instream salmonid habitat restoration 1. JAWRA J. Am. Water Resour. Assoc. 2004, 40, 1631–1644. [Google Scholar] [CrossRef]
- McMahon, T.E.; Hartman, G.F. Influence of cover complexity and current velocity on winter habitat use by juvenile coho salmon (oncorhynchus kisutch). Can. J. Fish. Aquat. Sci. 1989, 46, 1551–1557. [Google Scholar] [CrossRef]
- Kurylyk, B.L.; Moore, R.D.; MacQuarrie, K.T. Scientific briefing: Quantifying streambed heat advection associated with groundwater–surface water interactions. Hydrol. Process. 2016, 30, 987–992. [Google Scholar] [CrossRef]
- Caissie, D.; Luce, C.H. Quantifying streambed advection and conduction heat fluxes. Water Resour. Res. 2017, 53, 1595–1624. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survey | Flight Dates | Canopy Cover (%) | Canopy Cover (m2) |
|---|---|---|---|
| Survey 1 | 22–23 May 2017 | 38 | 61,668 |
| Survey 2 | 19–20 June 2017 | 53 | 87,776 |
| Survey 3 | 23–24 July 2017 | 71 | 117,348 |
| Survey 4 | 15–16 August 2017 | 74 | 121,583 |
| Class Change | Area (m2) | Area (%) |
|---|---|---|
| Canopy to open channel | 25,989 | 16 |
| Open channel to canopy | 3197 | 2 |
| No change | 108,352 | 66 |
| Area not analyzed | 26,765 | 16 |
| Site | River Kilometer | Weekly Growth | Biweekly Growth | Monthly Growth | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (rkm) | Mean Bias | MAE b | RMSE c | Mean Bias | MAE | RMSE | Mean Bias | MAE | RMSE | |
| 1 a | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 2 | 2.6 | 0.3 | 0.9 | 1.2 | 0.3 | 0.9 | 1.1 | 0.4 | 0.9 | 1.2 |
| 3 | 1.7 | 0.6 | 0.8 | 1.0 | 0.7 | 0.8 | 1.0 | 0.7 | 0.9 | 1.1 |
| 4 | 0.0 | 0.3 | 0.7 | 0.9 | 0.4 | 0.7 | 0.8 | 0.5 | 0.7 | 0.9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willis, A.; Holmes, E. Eye in the Sky: Using UAV Imagery of Seasonal Riverine Canopy Growth to Model Water Temperature. Hydrology 2019, 6, 6. https://doi.org/10.3390/hydrology6010006
Willis A, Holmes E. Eye in the Sky: Using UAV Imagery of Seasonal Riverine Canopy Growth to Model Water Temperature. Hydrology. 2019; 6(1):6. https://doi.org/10.3390/hydrology6010006
Chicago/Turabian StyleWillis, Ann, and Eric Holmes. 2019. "Eye in the Sky: Using UAV Imagery of Seasonal Riverine Canopy Growth to Model Water Temperature" Hydrology 6, no. 1: 6. https://doi.org/10.3390/hydrology6010006
APA StyleWillis, A., & Holmes, E. (2019). Eye in the Sky: Using UAV Imagery of Seasonal Riverine Canopy Growth to Model Water Temperature. Hydrology, 6(1), 6. https://doi.org/10.3390/hydrology6010006