Submarine Groundwater Discharge Differentially Modifies Photosynthesis, Growth, and Morphology for Two Contrasting Species of Gracilaria (Rhodophyta)


Abstract
1. Introduction
2. Materials and Methods
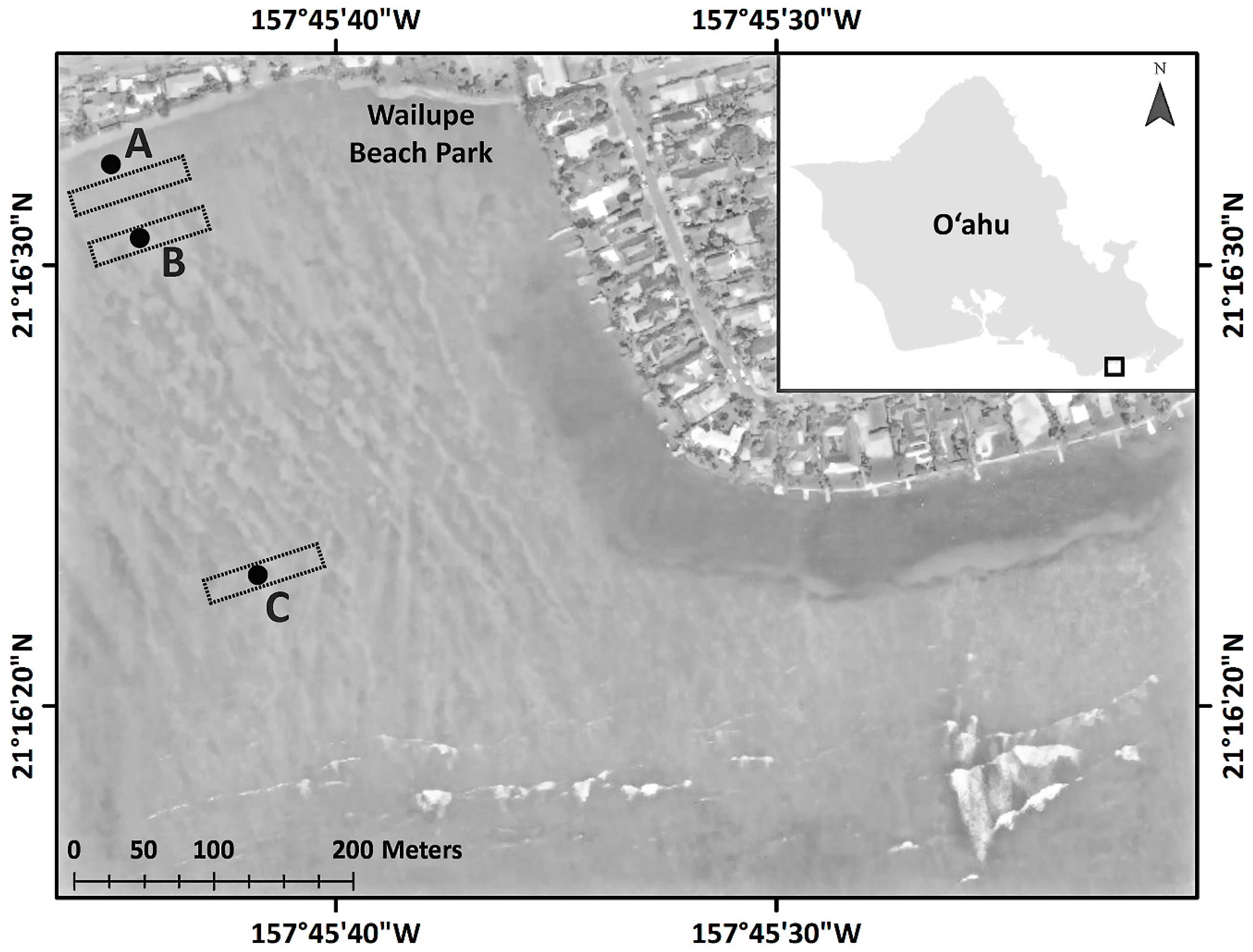
2.1. Field Experiment: Wailupe Reef
2.1.1. Sample Selection, Pretreatment, and Initial Measurements
2.1.2. Sample and Data Logger Deployment
2.1.3. Benthic Community Assessment
2.2. Simulated SGD Study
2.2.1. Sample Selection, Pretreatment, Mesocosm Design, and Initial Measurements
2.2.2. Data Collection
3. Results
3.1. Wailupe Reef Study
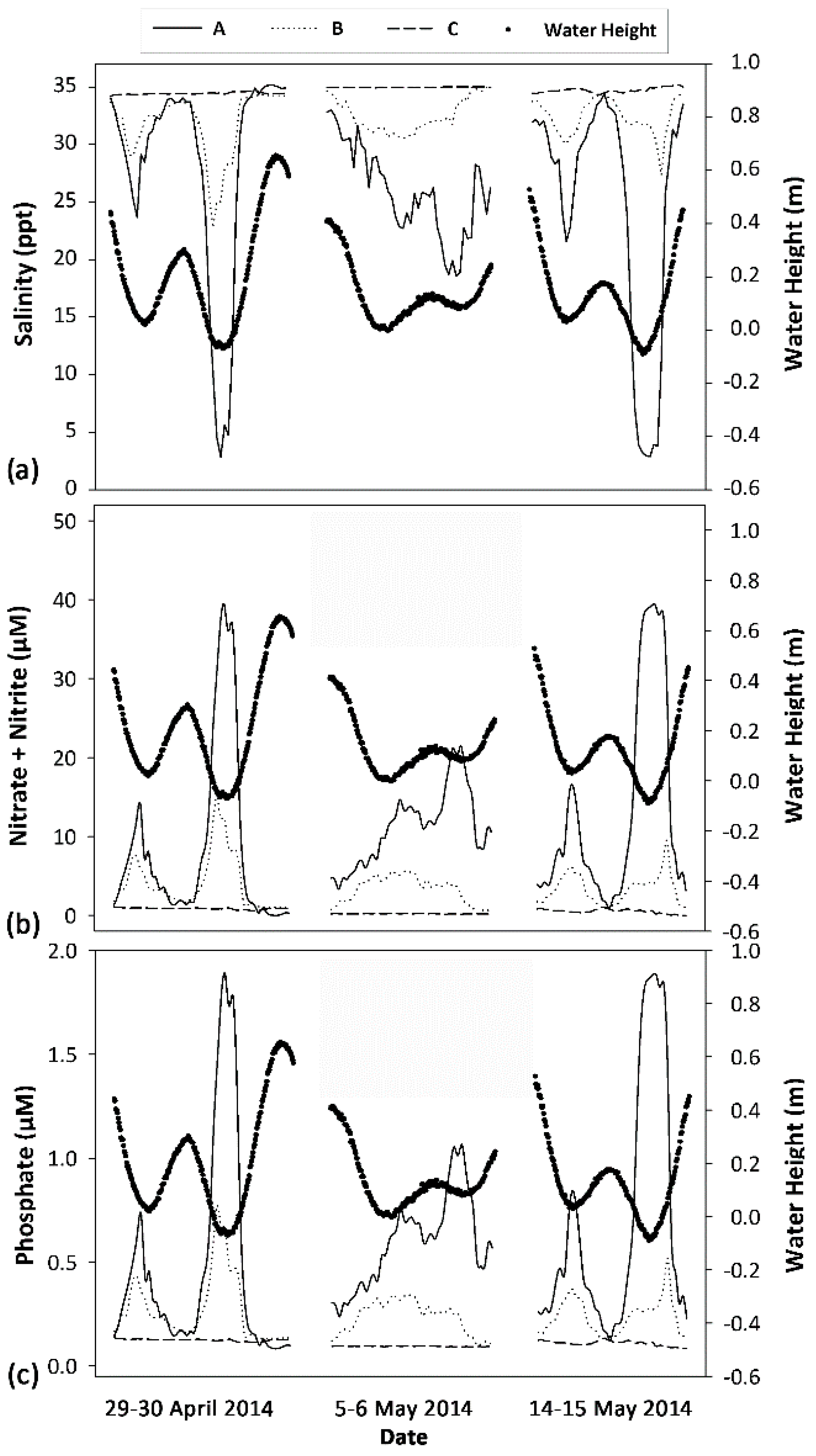
3.1.1. Physical Conditions at Wailupe Reef
3.1.2. Physiological Response of Deployed Algae
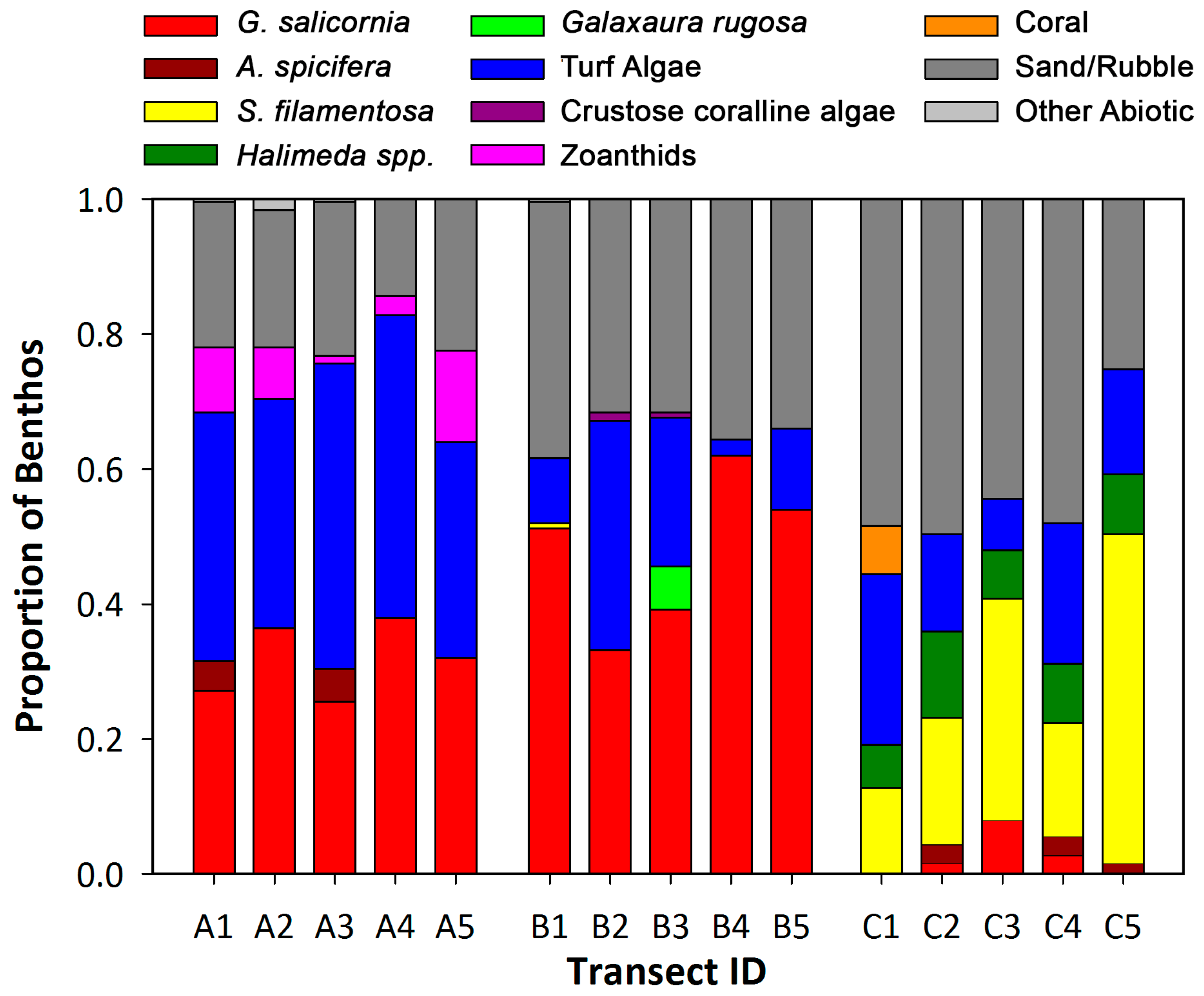
3.1.3. Benthic Community Assessment of Wailupe Reef
3.2. Simulated SGD Study
3.2.1. Growth Rate and Branch Development: Pooled Trials
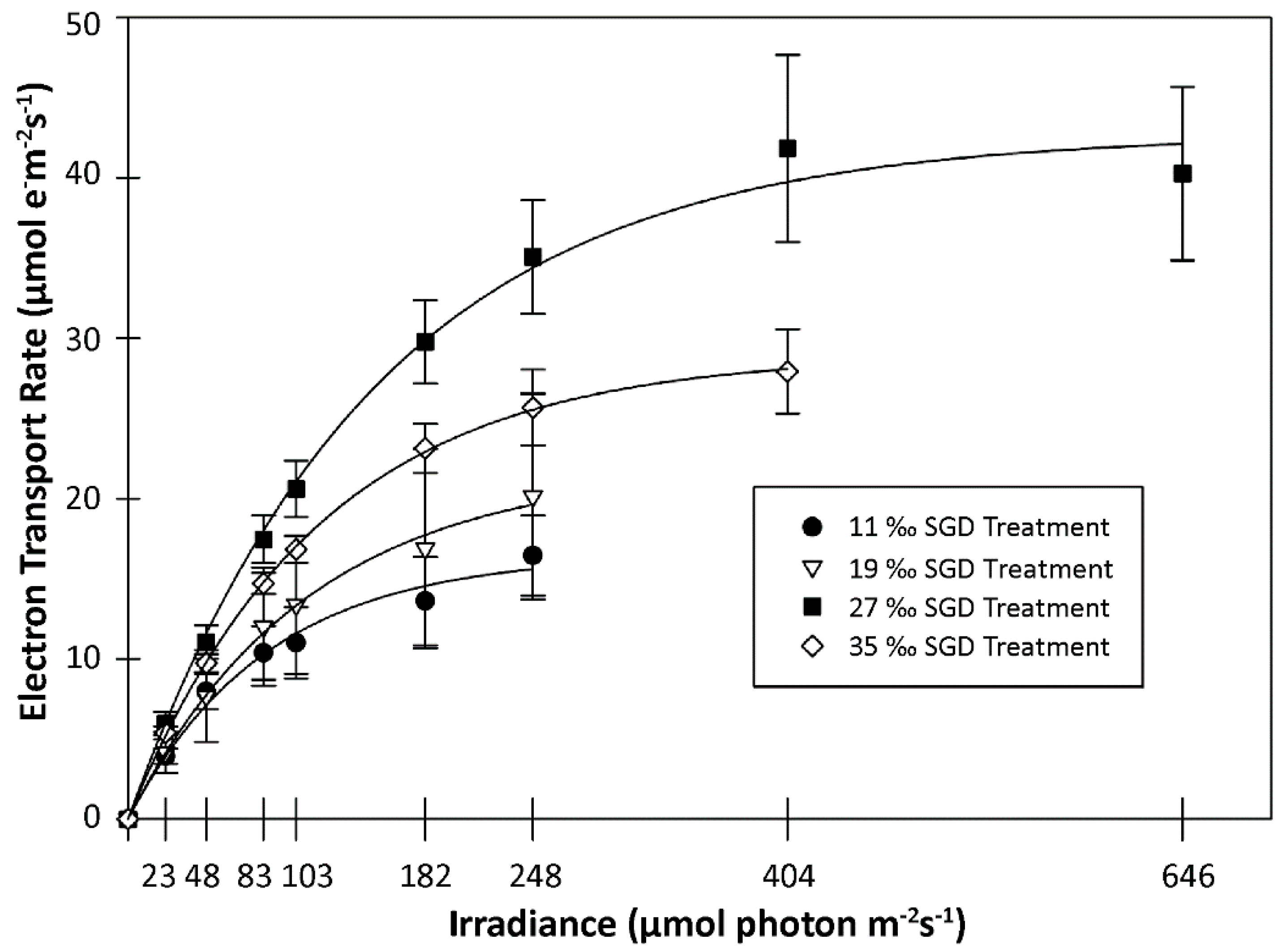
3.2.2. Photosynthetic Response: Replicate Trial 2
4. Discussion
4.1. Physiological Responses of Gracilaria to SGD
4.2. Benthic Community Structure along an SGD Gradient
4.3. Coastal Management Implications
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | G. coronopifolia | G. salicornia | |
|---|---|---|---|
| δ15N (‰) | 3 | 5.7 ± 0.3 | 5.3 ± 0.3 |
| N% | 3 | 1.0 ± 0.1 | 0.6 ± 0.1 |
| C% | 3 | 25.9 ± 1.3 | 10.0 ± 5.9 |
| C:N | 3 | 25.7 ± 1.4 | 16.8 ± 8.4 |
| ETRmax | 6 | 21.6 ± 3.6 | 23.3 ± 5.2 |
| alpha | 6 | 0.25 ± 0.0 | 0.23 ± 0.1 |
| Ek | 6 | 85.2 ± 6.0 | 112.9 ± 42.3 |
| Sample Name | Date | Latitude | Longitude | Salinity | PO43− | SiO42− | NO3− + NO2− | NH4+ |
|---|---|---|---|---|---|---|---|---|
| Wailupe A Low | 6 May 2014 | 21.27565 | −157.76250 | 21.3 | 1.04 | 312.20 | 23.50 | 1.04 |
| Wailupe B Low | 6 May 2014 | 21.27517 | −157.76230 | 34.9 | 0.08 | 16.50 | 0.58 | 0.29 |
| Wailupe C Low | 6 May 2014 | 21.27298 | −157.76151 | 35.0 | 0.09 | 1.60 | 0.09 | 0.46 |
| Wailupe A High | 14 May 2014 | 21.27565 | −157.76250 | 34.3 | 0.14 | 25.40 | 1.86 | 0.75 |
| Wailupe B High | 14 May 2014 | 21.27517 | −157.76230 | 34.7 | 0.05 | 6.70 | 0.18 | 0.38 |
| Wailupe C High | 14 May 2014 | 21.27298 | −157.76151 | 34.5 | 0.02 | 9.50 | 0.08 | 3.25 |
| Wailupe A LowLow | 15 May 2014 | 21.27565 | −157.76250 | 3.0 | 1.95 | 668.80 | 42.70 | 0.64 |
| Wailupe B LowLow1 | 15 May 2014 | 21.27517 | −157.76230 | 11.0 | 1.26 | 492.60 | 21.66 | 0.78 |
| Wailupe B LowLow2 | 15 May 2014 | 21.27517 | −157.76230 | 24.0 | 0.59 | 217.20 | 8.08 | 1.66 |
| Wailupe C LowLow | 15 May 2014 | 21.27298 | −157.76151 | 34.4 | 0.02 | 2.30 | 0.08 | 0.45 |
| Wailupe PP1 Low | 30 May 2010 | 21.27543 | −157.76248 | 25.82 | 0.56 | 210.40 | 15.85 | 0.98 |
| Wailupe PP2 Low | 30 May 2010 | 21.28592 | −157.79432 | 34.98 | 0.23 | 21.60 | 0.58 | 0.52 |
| Wailupe PP3 Low | 30 May 2010 | 21.27545 | −157.76247 | 25.98 | 0.74 | 202.50 | 11.94 | 5.41 |
| Wailupe PP4 Low | 30 May 2010 | 21.27530 | −157.76245 | 33.51 | 0.09 | 42.50 | 0.57 | 0.74 |
| Wailupe PP5 Low | 30 May 2010 | 21.27520 | −157.76240 | 33.98 | 0.13 | 25.30 | 0.38 | 0.39 |
| Wailupe PP1 High | 30 May 2010 | 21.27543 | −157.76248 | 35.13 | 0.12 | 7.70 | 0.18 | 0.28 |
| Wailupe PP2 High | 30 May 2010 | 21.28592 | −157.79432 | 35.12 | 0.10 | 7.30 | 0.17 | 0.45 |
| Wailupe PP3 High | 30 May 2010 | 21.27545 | −157.76247 | 35.14 | 0.13 | 7.90 | 0.16 | 0.50 |
| Wailupe PP4 High | 30 May 2010 | 21.27530 | −157.76245 | 35.14 | 0.12 | 8.30 | 0.16 | 0.26 |
| Wailupe PP5 High | 30 May 2010 | 21.27520 | −157.76240 | 35.15 | 0.10 | 6.40 | 0.13 | 0.17 |
References
- Burnett, W.C.; Bokuniewicz, H.; Huettel, M.; Moore, W.S.; Taniguchi, M. Groundwater and pore water inputs to the coastal zone. Biogeochemistry 2003, 66, 3–33. [Google Scholar] [CrossRef]
- Beusen, A.H.W.; Slomp, C.P.; Bouwman, A.F. Global land–ocean linkage: Direct inputs of nitrogen to coastal waters via submarine groundwater discharge. Environ. Res. Lett. 2013, 8, 034035. [Google Scholar] [CrossRef]
- Moore, W.S. The effect of submarine groundwater discharge on the ocean. Ann. Rev. Mar. Sci. 2010, 2, 59–88. [Google Scholar] [CrossRef] [PubMed]
- Dulai, H.; Kleven, A.; Ruttenberg, K.; Briggs, R.; Thomas, F. Evaluation of submarine groundwater discharge as a coastal nutrient source and its role in coastal groundwater quality and quantity. In Emerging Issues in Groundwater Resources; Fares, A., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 187–221. [Google Scholar] [CrossRef]
- Kwon, E.Y.; Kim, G.; Primeau, F.; Moore, W.S.; Cho, H.-M.; DeVries, T.; Sarmiento, J.L.; Charette, M.A.; Cho, Y.-K. Global estimate of submarine groundwater discharge based on an observationally constrained radium isotope model. Geophys. Res. Lett. 2014, 41, 8438–8444. [Google Scholar] [CrossRef]
- Lecher, A.; Mackey, K. Synthesizing the effects of submarine groundwater discharge on marine biota. Hydrology 2018, 5, 60. [Google Scholar] [CrossRef]
- Kim, G.; Kim, J.S.; Hwang, D.W. Submarine groundwater discharge from oceanic islands standing in oligotrophic oceans: Implications for global biological production and organic carbon fluxes. Limnol. Oceanogr. 2011, 56, 673–682. [Google Scholar] [CrossRef]
- Larned, S.T. Nitrogen- versus phosphorus-limited growth and sources of nutrients for coral reef macroalgae. Mar. Biol. 1998, 132, 409–421. [Google Scholar] [CrossRef]
- Downing, J.; McClain, M.; Twilley, R.; Melack, J.; Elser, J.; Rabalais, N.; Lewis, W., Jr.; Turner, R.; Corredor, J.; Soto, D. The impact of accelerating land-use change on the N-cycle of tropical aquatic ecosystems: Current conditions and projected changes. Biogeochemistry 1999, 46, 109–148. [Google Scholar] [CrossRef]
- Johnson, A.G.; Glenn, C.R.; Burnett, W.C.; Peterson, R.N.; Lucey, P.G. Aerial infrared imaging reveals large nutrient-rich groundwater inputs to the ocean. Geophys. Res. Lett. 2008, 35, L15606. [Google Scholar] [CrossRef]
- McCoy, C.A.; Corbett, D.R. Review of submarine groundwater discharge (SGD) in coastal zones of the Southeast and Gulf Coast regions of the United States with management implications. J. Environ. Manag. 2009, 90, 644–651. [Google Scholar] [CrossRef]
- Bishop, J.M.; Glenn, C.R.; Amato, D.W.; Dulai, H. Effect of land use and groundwater flow path on submarine groundwater discharge nutrient flux. J. Hydrol. Reg. Stud. 2017, 11, 194–218. [Google Scholar] [CrossRef]
- Knee, K.; Street, J.H.; Grossman, E.G.; Paytan, A. Nutrient inputs to the coastal ocean from submarine groundwater discharge in a groundwater-dominated system: Relation to land use (Kona coast, Hawaii, U.S.A.). Limnol. Oceanogr. 2010, 55, 1105–1122. [Google Scholar] [CrossRef]
- Moosdorf, N.; Stieglitz, T.; Waska, H.; Dürr, H.H.; Hartmann, J. Submarine groundwater discharge from tropical islands: A review. Grundwasser 2015, 20, 53–67. [Google Scholar] [CrossRef]
- Dailer, M.L.; Smith, J.E.; Smith, C.M. Responses of bloom forming and non-bloom forming macroalgae to nutrient enrichment in Hawai‘i, USA. Harmful Algae 2012, 17, 111–125. [Google Scholar] [CrossRef]
- Pedersen, M.F.; Borum, J. Nutrient control of algal growth in estuarine waters. Nutrient limitation and the importance of nitrogen requirements and nitrogen storage among phytoplankton and species of macroalgae. Mar. Ecol. Prog. Ser. 1996, 142, 261–272. [Google Scholar] [CrossRef]
- Amato, D.W.; Bishop, J.M.; Glenn, C.R.; Dulai, H.; Smith, C.M. Impact of submarine groundwater discharge on marine water quality and reef biota of Maui. PLoS ONE 2016, 11, e0165825. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 54, 995–1010. [Google Scholar] [CrossRef]
- Valiela, I.; Foreman, K.; Lamontagne, M.; Hersh, D.; Costa, J.; Peckol, P.; Demeoandreson, B.; Davanzo, C.; Babione, M.; Sham, C.H.; et al. Couplings of watersheds and coastal waters: Sources and consequences of nutrient enrichment in Waquoit Bay, Massachusetts. Estuaries 1992, 15, 443–457. [Google Scholar] [CrossRef]
- Paerl, H.W. Coastal eutrophication and harmful algal blooms: Importance of atmospheric deposition and groundwater as ‘‘new’’ nitrogen and other nutrient sources. Limnol. Oceanogr. 1997, 42, 1154–1165. [Google Scholar] [CrossRef]
- Herrera-Silveira, J.A.; Morales-Ojeda, S.M. Evaluation of the health status of a coastal ecosystem in southeast Mexico: Assessment of water quality, phytoplankton and submerged aquatic vegetation. Mar. Pollut. Bull. 2009, 59, 72–86. [Google Scholar] [CrossRef]
- Smith, J.E.; Runcie, J.W.; Smith, C.M. Characterization of a large-scale ephemeral bloom of the green alga Cladophora sericea on the coral reefs of west Maui, Hawaii. Mar. Ecol. Prog. Ser. 2005, 302, 77–91. [Google Scholar] [CrossRef]
- Kirst, G.O. Salinity tolerance of eukaryotic marine algae. Annu. Rev. Plant Biol. 1989, 40, 21–53. [Google Scholar] [CrossRef]
- Dawes, C.J.; Orduña-rojas, J.; Robledo, D. Response of the tropical red seaweed Gracilaria cornea to temperature, salinity and irradiance. J. Appl. Phycol. 1999, 10, 419–425. [Google Scholar] [CrossRef]
- Choi, T.S.; Kang, E.J.; Kim, J.-h.; Kim, K.Y. Effect of salinity on growth and nutrient uptake of Ulva pertusa (Chlorophyta) from an eelgrass bed. Algae 2010, 25, 17–26. [Google Scholar] [CrossRef]
- Yu, C.-H.; Lim, P.-E.; Phang, S.-M. Effects of irradiance and salinity on the growth of carpospore-derived tetrasporophytes of Gracilaria edulis and Gracilaria tenuistipitata var liui (Rhodophyta). J. Appl. Phycol. 2013, 25, 787–794. [Google Scholar] [CrossRef]
- Smith, J.E.; Hunter, C.L.; Conklin, E.J.; Most, R.; Sauvage, T.; Squair, C.; Smith, C.M. Ecology of the invasive red alga Gracilaria salicornia (Rhodophyta) on Oahu, Hawaii. Pac. Sci. 2004, 58, 325–343. [Google Scholar] [CrossRef]
- Martinez, J.A.; Smith, C.M.; Richmond, R.H. Invasive algal mats degrade coral reef physical habitat quality. Estuar. Coast. Shelf Sci. 2012, 99, 42–49. [Google Scholar] [CrossRef]
- Smith, J.E.; Hunter, C.L.; Smith, C.M. Distribution and reproductive characteristics of nonindigenous and invasive marine algae in the Hawaiian Islands. Pac. Sci. 2002, 56, 299–315. [Google Scholar] [CrossRef]
- Abbott, I.A. Marine Red Algae of the Hawaiian Islands; Bishop Museum Press: Honolulu, HI, USA, 1999. [Google Scholar]
- Abbott, I.A.; Willamson, E.H. Limu: An Ethnobotanical Study of Some Edible Hawaiian Seaweeds; Pacific Tropical Botanical Garden: Lawai, HI, USA, 1974. [Google Scholar]
- Dailer, M.L.; Ramey, H.L.; Saephan, S.; Smith, C.M. Algal δ15N values detect a wastewater effluent plume in nearshore and offshore surface waters and three-dimensionally model the plume across a coral reef on Maui, Hawaii, USA. Mar. Pollut. Bull. 2012, 64, 207–213. [Google Scholar] [CrossRef]
- Swarzenski, P.W.; Dulaiova, H.; Dailer, M.L.; Glenn, C.R.; Smith, C.G.; Storlazzi, C.D. A geochemical and geophysical assessment of coastal groundwater discharge at select sites in Maui and Oahu, Hawaii. In Groundwater in the Coastal Zones of Asia-Pacific; Wetzelhuetter, C., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 27–46. [Google Scholar] [CrossRef]
- McGowan, M.P. Submarine groundwater discharge: Freshwater and nutrient input into Hawaiiʻs coastal zone. Master’s Thesis, University of Hawaii at Mānoa, Honolulu, HI, USA, 2004. [Google Scholar]
- Richardson, C.M.; Dulai, H.; Popp, B.N.; Ruttenberg, K.; Fackrell, J.K. Submarine groundwater discharge drives biogeochemistry in two Hawaiian reefs. Limnol. Oceanogr. 2017, 62, S348–S363. [Google Scholar] [CrossRef]
- Holleman, K.D. Impact of flux, residence time and nutrient load of submarine groundwater discharge on coastal phytoplankton growth in coastal waters of Hawaii. Master’s Thesis, University of Hawaii at Mānoa, Honolulu, HI, USA, 2011. [Google Scholar]
- Dimova, N.T.; Swarzenski, P.W.; Dulaiova, H.; Glenn, C.R. Utilizing multichannel electrical resistivity methods to examine the dynamics of the fresh water–seawater interface in two Hawaiian groundwater systems. J. Geophys. Res. 2012, 117, C02012. [Google Scholar] [CrossRef]
- Sweeney, R.; Liu, K.; Kaplan, I. Oceanic nitrogen isotopes and their uses in determining the source of sedimentary nitrogen. In Stable Isotopes in the Earth Sciences; Robinson, B.W., Ed.; New Zealand DSIR Bulletins: Lower Hutt, New Zealand, 1978; Volume 9, pp. 9–26. [Google Scholar]
- Ralph, P.J.; Gademann, R. Rapid light curves: A powerful tool to assess photosynthetic activity. Aquat. Bot. 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Jokiel, P.L. CRAMP Rapid Assessment: Benthic Protocols. Available online: http://cramp.wcc.hawaii.edu/Rapid_Assessment_Files/RA_benthic_protocol.htm (accessed on 3 January 2014).
- Bird, C.E. PhotoGrid: ecological analysis of digital photographs. University of Hawaii: Honolulu, HI, USA, 2001. Available online: http://www.photogrid.netfirms.com/ (accessed on 12 November 2018).
- Shannon, C.E. A mathematical theory of communication. Bell. Syst. Tech. J. 1948, 27, 623–656. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Karl, D.M.; Tien, G. Temporal variability in dissolved phosphorus concentrations in the subtropical North Pacific Ocean. Mar. Chem. 1997, 56, 77–96. [Google Scholar] [CrossRef]
- Cho, H.-M.; Kim, G.; Kwon, E.Y.; Moosdorf, N.; Garcia-Orellana, J.; Santos, I.R. Radium tracing nutrient inputs through submarine groundwater discharge in the global ocean. Sci. Rep. 2018, 8, 2439. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.L.; Dulai, H.; Glenn, C.R.; Lucey, P.G. Integration of aerial infrared thermography and in situ radon-222 to investigate submarine groundwater discharge to Pearl Harbor, Hawaii, USA. Limnol. Oceanogr. 2018, 999, 20. [Google Scholar] [CrossRef]
- Choi, H.G.; Kim, Y.S.; Kim, J.H.; Lee, S.J.; Park, E.J.; Ryu, J.; Nam, K.W. Effects of temperature and salinity on the growth of Gracilaria verrucosa and G. chorda, with the potential for mariculture in Korea. J. Appl. Phycol. 2006, 18, 269–277. [Google Scholar] [CrossRef]
- Israel, A.; Martinez-Goss, M.; Friedlander, M. Effect of salinity and pH on growth and agar yield of Gracilaria tenuistipitata var. liui in laboratory and outdoor cultivation. J. Appl. Phycol. 1999, 11, 543–549. [Google Scholar] [CrossRef]
- Norton, T.; Mathieson, A.; Neushul, M. Morphology and environment. In The Biology of Seaweeds; Lobban, C.S., Wynne, M., Eds.; University of California Press: Berkeley, CA, USA, 1981; Volume 17, pp. 421–451. [Google Scholar]
- Zablackis, E. The effect of salinity on growth rate and branch morphology in tank cultivated Grateloupia filicina (Rhodophyta) in Hawaii. Aquat. Bot. 1987, 27, 187–193. [Google Scholar] [CrossRef]
- Jordan, A.J.; Vadas, R.L. Influence of environmental parameters on intraspecific variation in Fucus vesiculosus. Mar. Biol. 1972, 14, 248–252. [Google Scholar]
- Mathieson, A.C.; Burns, R.L. Ecological studies of economic red algae. V. growth and reproduction of natural and harvested populations of Chondrus crispus Stackhouse in New Hampshire. J. Exp. Mar. Biol. Ecol. 1975, 17, 137–156. [Google Scholar] [CrossRef]
- Reed, R.; Russell, G. Salinity fluctuations and their influence on “bottle brush” morphogenesis in Enteromorpha intestinalis (L.) Link. Br. Phycol. J. 1978, 13, 149–153. [Google Scholar] [CrossRef]
- Richardson, C.M.; Dulai, H.; Whittier, R.B. Sources and spatial variability of groundwater-delivered nutrients in Maunalua Bay, Oahu, Hawaii. J. Hydrol. Reg. Stud. 2017, 11, 178–193. [Google Scholar] [CrossRef]




| Species | Location | n | δ15N‰ | N% | C% | C:N |
|---|---|---|---|---|---|---|
| G. coronopifolia | A | 3 | 5.9 ± 0.1 A | 1.5 ± 0.2 A | 21.6 ± 4.0 A | 14.9 ± 0.8 A |
| G. coronopifolia | B | 5 | 6.3 ± 0.4 A | 1.0 ± 0.1 B | 20.4 ± 2.4 A | 20.6 ± 1.7 AB |
| G. coronopifolia | C | 6 | 3.6 ± 0.5 B | 0.8 ± 0.0 C | 20.9 ± 3.7 A | 27.5 ± 7.3 B |
| G. salicornia | A | 6 | 7.0 ± 1.3 A | 1.2 ± 0.2 A | 14.0 ± 1.9 A | 12.4 ± 1.3 A |
| G. salicornia | B | 6 | 6.5 ± 0.4 A | 0.7 ± 0.1 B | 9.7 ± 3.3 B | 13.7 ± 3.6 A |
| G. salicornia | C | 6 | 4.3 ± 0.3 B | 0.6 ± 0.1 B | 14.3 ± 2.8 A | 24.9 ± 5.9 B |
| Species | Treatment | n | Growth Rate | Tip Score | Tip Index | ETRmax | alpha | Ek | |
|---|---|---|---|---|---|---|---|---|---|
| Simulated SGD Study | Gc | 11‰ | 12 | 1.4 ± 1.5 AB | 31.0 ± 21.4 AB | −50.4 ± 32.4 A | 13.8 ± 4.1 A | 0.23 ± 0.03 A | 59.5 ± 18.2 A |
| Gc | 19‰ | 12 | 2.2 ± 0.5 B | 110.3 ± 32.7 AB | 83.8 ± 53.8 BCD | 17.1 ± 9.4 A | 0.21 ± 0.07 A | 77.1 ± 23.3 A | |
| Gc | 27‰ | 12 | 3.0 ± 0.6 C | 117.6 ± 35.7 B | 132.7 ± 105.8 C | 44.7 ± 8.1 B | 0.27 ± 0.01 A | 163 ± 27.9 B | |
| Gc | 35‰ | 12 | 1.1 ± 0.4 A | 79.1 ± 15.3 AB | 30.2 ± 41.0 AD | 29.3 ± 3.4 C | 0.24 ± 0.02 A | 124.1 ± 20.2 C | |
| Wailupe Field Study | Gc | A | 3 | 2.2 ± 1.8 A | 99.7 ± 30.2 A | 94.8 ± 154.1 A | 34.8 ± 7.3 A | 0.16 ± 0.02 A | 218.6 ± 49.8 A |
| Gc | B | 5 | 2.7 ± 1.7 A | 128.3 ± 27.0 A | 110.2 ± 64.4 A | 30.2 ± 2.0 A | 0.24 ± 0.05 A | 127.6 ± 23.8 B | |
| Gc | C | 6 | 3.8 ± 4.7 A | 112.2 ± 34.1 A | 110.4 ± 121.0 A | 28.6 ± 3.1 A | 0.19 ± 0.05 A | 159.8 ± 45.9 AB | |
| Gs | A | 6 | 0.7 ± 2.0 A | 28.0 ± 6.0 A | 144.7 ± 105.7 A | 35.8 ± 3.9 A | 0.25 ± 0.06 A | 147.8 ± 34.5 A | |
| Gs | B | 6 | 1.6 ± 1.0 A | 17.1 ± 2.3 B | 59.7 ± 46.8 AB | 40.4 ± 8.8 A | 0.25 ± 0.03 A | 161.1 ± 26.0 A | |
| Gs | C | 6 | −0.8 ± 3.6 A | 16.6 ± 8.4 B | −14.8 ± 57.7 B | 25.0 ± 3.2 B | 0.15 ± 0.02 B | 168.9 ± 33.3 A |
| % | % | % | % | % | % | % | % | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Location | Macro-algae | Invasive | Native | Turf | CCA | Abiotic | Coral | Zoanthid | H’ | λ’ |
| A | 33.7 ± 3.3 A | 33.7 ± 3.3 A | 0.0 ± 0.0 A | 38.6 ± 6.1 A | 0.0 ± 0.0 A | 20.3 ± 3.4 A | 0.0 ± 0.0 A | 7.0 ± 5.1 A | 1.0 ± 0.1 A | 40.7 ± 4.4 A |
| B | 49.4 ± 10.8 A | 47.9 ± 11.6 B | 1.4 ± 2.8 A | 16.0 ± 2.3 BC | 0.4 ± 0.6 A | 34.1 ± 2.7 AC | 0.0 ± 0.0 A | 0.0 ± 0.0 B | 0.5 ± 0.3 B | 65.4 ± 19.8 B |
| C | 38.7 ± 15.4 A | 3.9 ± 3.2 C | 34.8 ± 14.9 B | 16.7 ± 6.7 C | 0.0 ± 0.0 A | 43.1 ± 10.2 BC | 1.4 ± 3.2 A | 0.0 ± 0.0 B | 1.1 ± 0.2 A | 37.9 ± 8.6 A |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amato, D.W.; Smith, C.M.; Duarte, T.K. Submarine Groundwater Discharge Differentially Modifies Photosynthesis, Growth, and Morphology for Two Contrasting Species of Gracilaria (Rhodophyta). Hydrology 2018, 5, 65. https://doi.org/10.3390/hydrology5040065
Amato DW, Smith CM, Duarte TK. Submarine Groundwater Discharge Differentially Modifies Photosynthesis, Growth, and Morphology for Two Contrasting Species of Gracilaria (Rhodophyta). Hydrology. 2018; 5(4):65. https://doi.org/10.3390/hydrology5040065
Chicago/Turabian StyleAmato, Daniel W., Celia M. Smith, and Thomas K. Duarte. 2018. "Submarine Groundwater Discharge Differentially Modifies Photosynthesis, Growth, and Morphology for Two Contrasting Species of Gracilaria (Rhodophyta)" Hydrology 5, no. 4: 65. https://doi.org/10.3390/hydrology5040065
APA StyleAmato, D. W., Smith, C. M., & Duarte, T. K. (2018). Submarine Groundwater Discharge Differentially Modifies Photosynthesis, Growth, and Morphology for Two Contrasting Species of Gracilaria (Rhodophyta). Hydrology, 5(4), 65. https://doi.org/10.3390/hydrology5040065