
Acute Cd Toxicity, Metal Accumulation, and Ion Loss in Southern Catfish (Silurus meridionalis Chen)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Water and Reagents
2.2. Experimental Fish and Acclimation
2.3. Acute Cd Exposure
2.4. Tissue Cd and Ion (Na+ and Ca2+) Analysis
2.5. Calculations and Statistical Analysis
3. Results
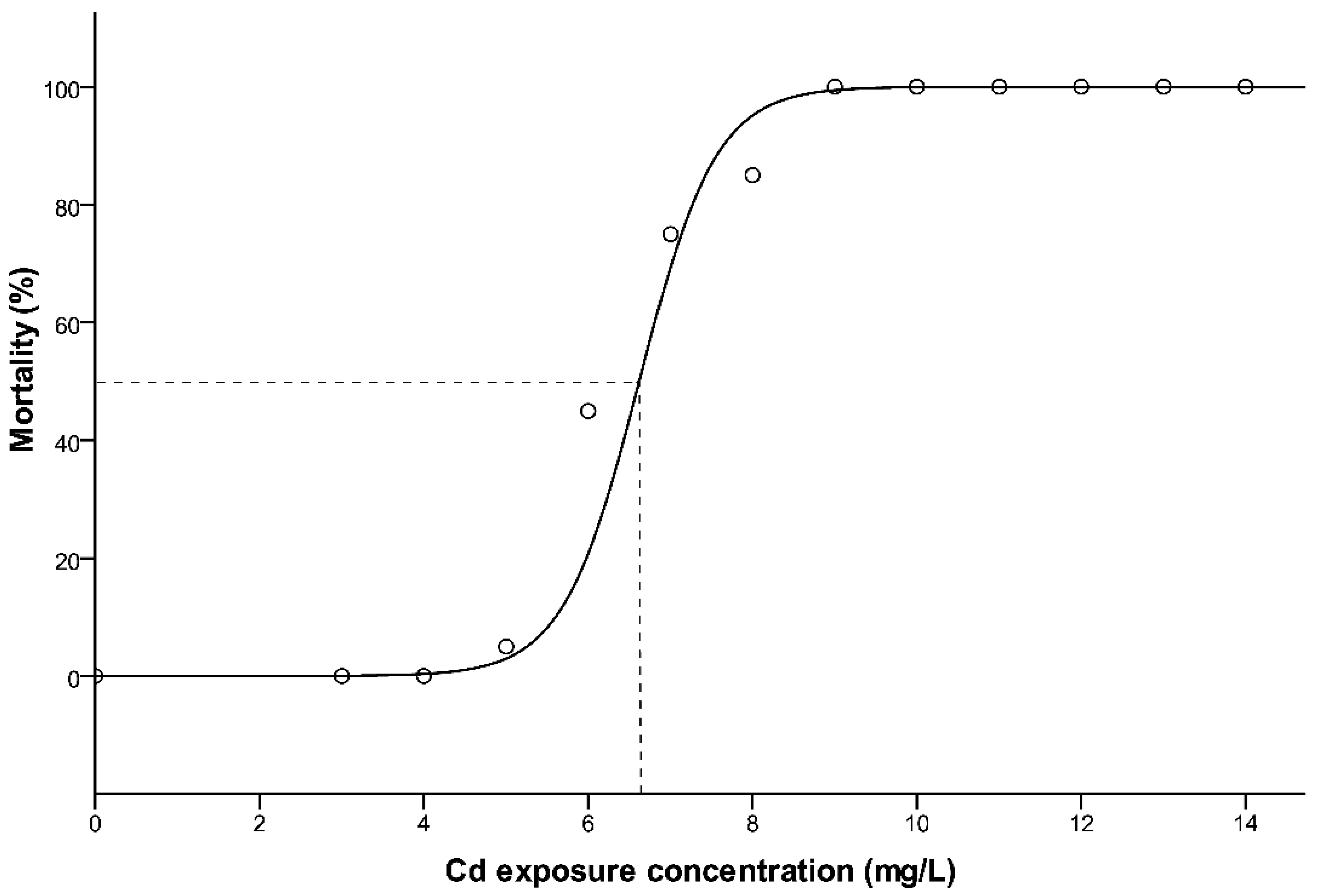
3.1. Mortality and 96 h LC50
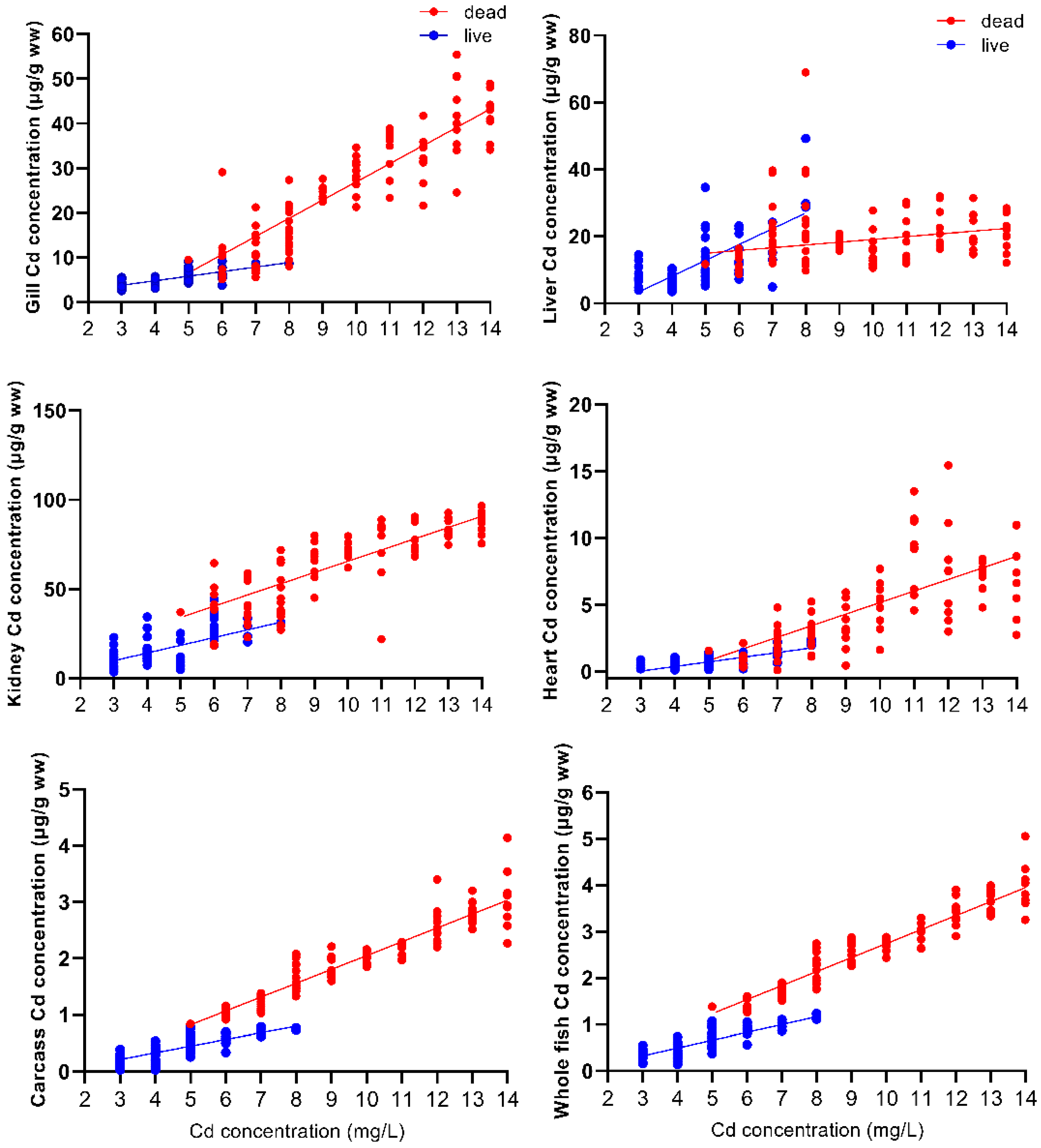
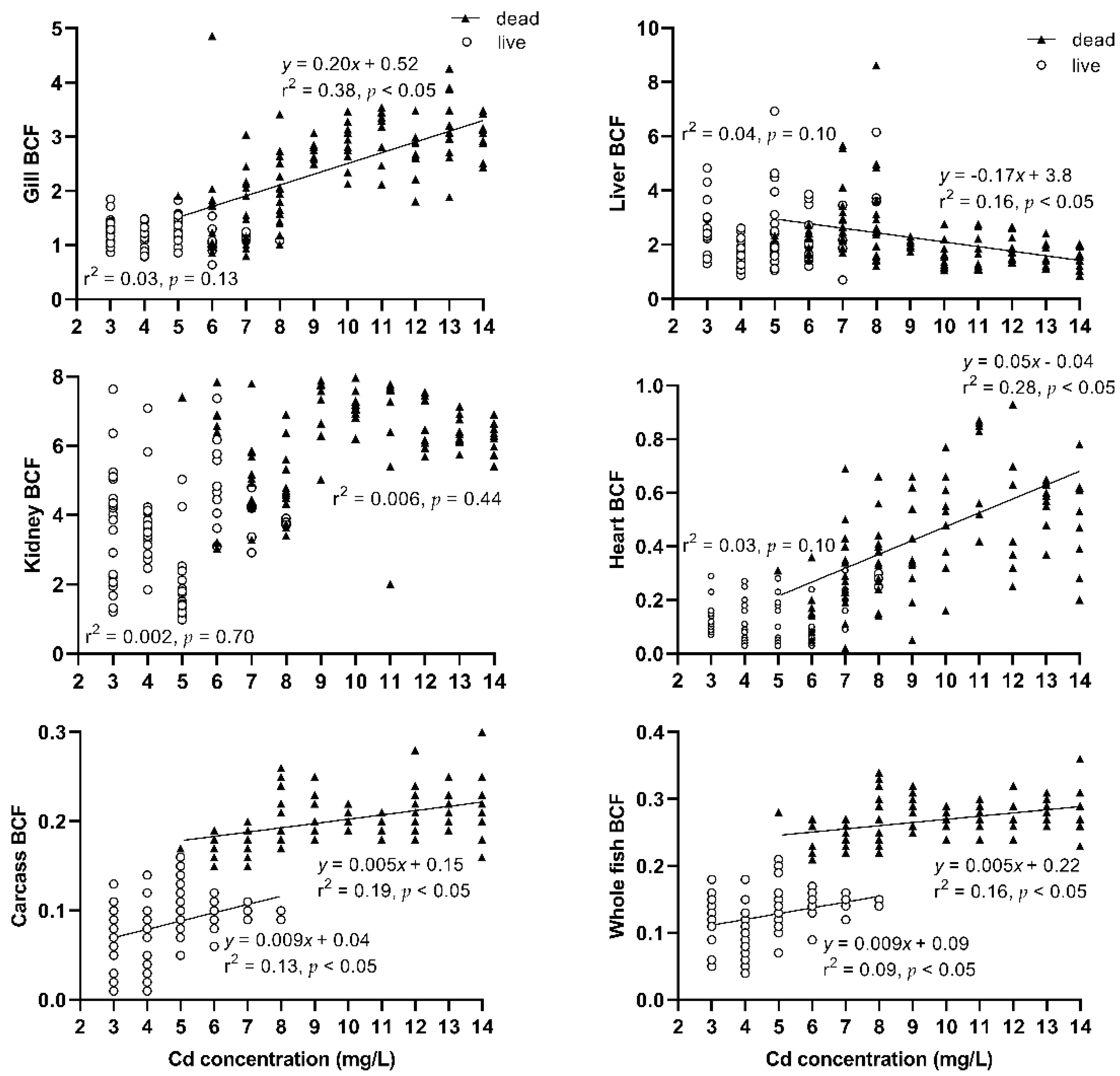
3.2. Cd Accumulation and Accumulation Rate Analysis of Tissues
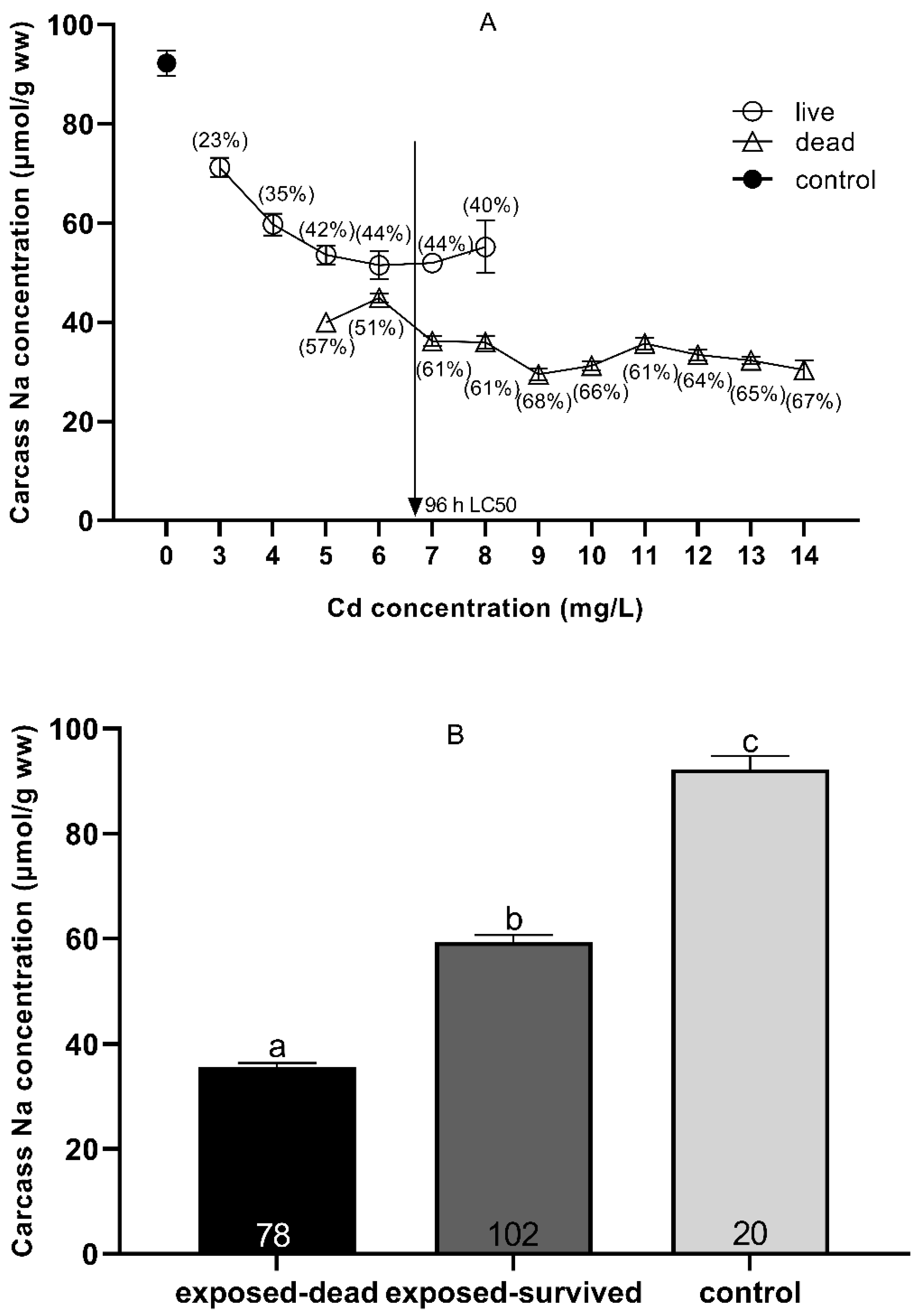
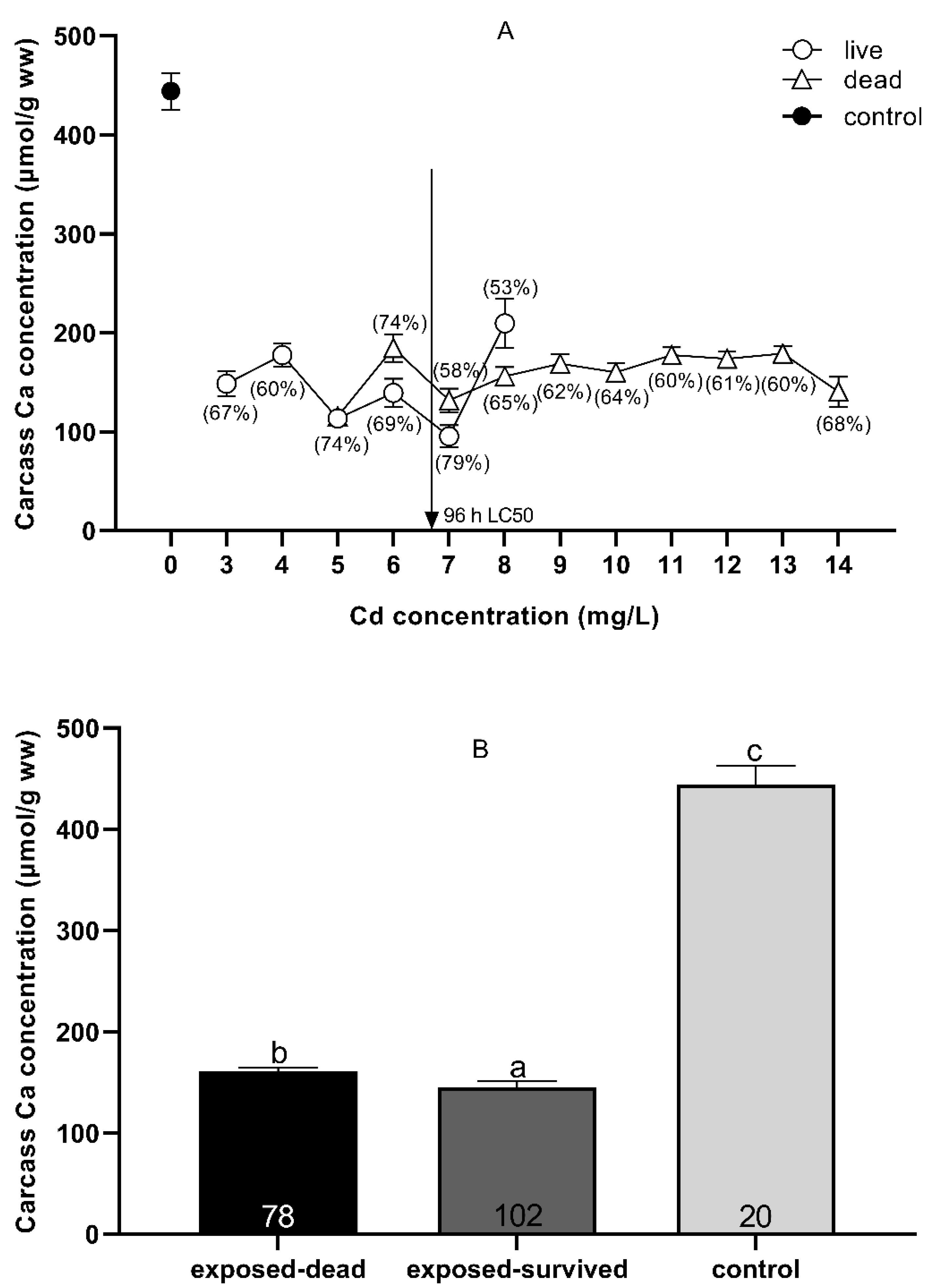
3.3. Carcass Na+ and Ca2+ Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Sokolova, I.M.; Lannig, G. Interactive effects of metal pollution and temperature on metabolism in aquatic ectotherms: Implications of global climate change. Clim. Res. 2008, 37, 181–201. [Google Scholar] [CrossRef] [Green Version]
- Fazio, F.; Piccione, G.; Tribulato, K.; Ferrantelli, V.; Giangrosso, G.; Arfuso, F.; Faggio, C. Bioaccumulation of heavy metals in blood and tissue of striped mullet in two Italian lakes. J. Aquat. Anim. Health 2014, 26, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Renieri, E.; Alegakis, A.; Kiriakakis, M.; Vinceti, M.; Ozcagli, E.; Wilks, M.; Tsatsakis, A. Cd, Pb and Hg Biomonitoring in Fish of the Mediterranean Region and Risk Estimations on Fish Consumption. Toxics 2014, 2, 417–442. [Google Scholar] [CrossRef] [Green Version]
- McGeer, J.C.; Szebedinszky, C.; Gordon McDonald, D.; Wood, C.M. Effects of chronic sublethal exposure to waterborne Cu, Cd or Zn in rainbow trout 2: Tissue specific metal accumulation. Aquat. Toxicol. 2000, 50, 245–256. [Google Scholar] [CrossRef]
- De Silva, N.A.L.; Marsden, I.D.; Gaw, S.; Glover, C.N. Acute waterborne cadmium toxicity in the estuarine pulmonate mud snail, Amphibola crenata. Ecotoxicol. Environ. Saf. 2018, 158, 274–283. [Google Scholar] [CrossRef]
- Shekh, K.; Tang, S.; Kodzhahinchev, V.; Niyogi, S.; Hecker, M. Species and life-stage specific differences in cadmium accumulation and cadmium induced oxidative stress, metallothionein and heat shock protein responses in white sturgeon and rainbow trout. Sci. Total Environ. 2019, 673, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Deidda, I.; Russo, R.; Bonaventura, R.; Costa, C.; Zito, F.; Lampiasi, N. Neurotoxicity in Marine Invertebrates: An Update. Biology 2021, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Sonone, S.S.; Jadhav, S.; Sankhla, M.S.; Kumar, R. Water Contamination by Heavy Metals and their Toxic Effect on Aquaculture and Human Health through Food Chain. Lett. Appl. NanoBioScience 2020, 10, 2148–2166. [Google Scholar] [CrossRef]
- Guo, S.; Zheng, J.; Yuan, S.; Zhu, Q. Effects of heat and cadmium exposure on stress-related responses in the liver of female zebrafish: Heat increases cadmium toxicity. Sci. Total Environ. 2018, 618, 1363–1370. [Google Scholar] [CrossRef]
- Yang, J.; Liu, D.; He, Y.; Wang, L. Mitochondrial energy metabolism in the hepatopancreas of freshwater crabs (Sinopotamon henanense) after cadmium exposure. Environ. Sci. Process Impacts 2014, 17, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.B.; Wu, Q.; Gao, G.L. Remediation of Lead-Zinc Contaminated Soil in China. Appl. Mech. Mater. 2012, 209–211, 1116–1119. [Google Scholar] [CrossRef]
- Bervoets, L.; Van Campenhout, K.; Reynders, H.; Knapen, D.; Covaci, A.; Blust, R. Bioaccumulation of micropollutants and biomarker responses in caged carp (Cyprinus carpio). Ecotoxicol. Environ. Saf. 2009, 72, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Tawwab, M.; Wafeek, M. Fluctuations in water temperature affected waterborne cadmium toxicity: Hematology, anaerobic glucose pathway, and oxidative stress status of Nile tilapia, Oreochromis niloticus (L.). Aquaculture 2017, 477, 106–111. [Google Scholar] [CrossRef]
- Alsop, D.; Wood, C.M. Metal uptake and acute toxicity in zebrafish: Common mechanisms across multiple metals. Aquat. Toxicol. 2011, 105, 385–393. [Google Scholar] [CrossRef] [PubMed]
- McGeer, J.C.; Niyogi, S.; Smith, D.S. Cadmium. In Fish Physiology; Wood, C.M., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 31B, pp. 125–184. [Google Scholar]
- Arini, A.; Gourves, P.Y.; Gonzalez, P.; Baudrimont, M. Metal detoxification and gene expression regulation after a Cd and Zn contamination: An experimental study on Danio rerio. Chemosphere 2015, 128, 125–133. [Google Scholar] [CrossRef]
- Kakade, A.; Salama, E.; Pengya, F.; Liu, P.; Li, X. Long-term exposure of high concentration heavy metals induced toxicity, fatality, and gut microbial dysbiosis in common carp, Cyprinus carpio. Environ. Pollut. 2020, 266, 115293. [Google Scholar] [CrossRef]
- Acosta, I.B.; Junior, A.S.V.; Silva, E.F.E.; Cardoso, T.F.; Caldas, J.S.; Jardim, R.D.; Corcini, C.D. Effects of exposure to cadmium in sperm cells of zebrafish, Danio rerio. Toxicol. Rep. 2016, 3, 696–700. [Google Scholar] [CrossRef] [Green Version]
- Renieri, E.A.; Sfakianakis, D.G.; Alegakis, A.A.; Safenkova, I.V.; Buha, A.; Matović, V.; Tzardi, M.; Dzantiev, B.B.; Divanach, P.; Kentouri, M.; et al. Nonlinear responses to waterborne cadmium exposure in zebrafish. An in vivo study. Environ. Res. 2017, 157, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Irfan, M.; Liu, X.; Hussain, K.; Mushtaq, S.; Cabrera, J.; Zhang, P. The global research trend on cadmium in freshwater: A bibliometric review. Environ. Sci. Pollut. Res. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Sprague, J.B. Measurement of pollutant toxicity to fish I. bioassay methods for acute toxicity. Water Res. 1969, 3, 793–821. [Google Scholar] [CrossRef]
- Redeker, E.S.; Blust, R. Accumulation and Toxicity of Cadmium in the Aquatic Oligochaete Tubifex tubifex: A Kinetic Modeling Approach. Environ. Sci. Technol. 2004, 38, 537–543. [Google Scholar] [CrossRef]
- Tsui, M.T.K.; Wang, W. Acute Toxicity of Mercury to Daphnia magna under Different Conditions. Environ. Sci. Technol. 2006, 40, 4025–4030. [Google Scholar] [CrossRef] [PubMed]
- US EPA. Ecotox Database. Available online: https://cfpub.epa.gov/ecotox/index.cfm (accessed on 12 October 2020).
- Vergauwen, L.; Knapen, D.; Hagenaars, A.; Blust, R. Hypothermal and hyperthermal acclimation differentially modulate cadmium accumulation and toxicity in the zebrafish. Chemosphere 2013, 91, 521–529. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Zhang, T.; Zhang, R.; Liu, R.; Chen, Y. Molecular mechanism on cadmium-induced activity changes of catalase and superoxide dismutase. Int. J. Biol. Macromol. 2015, 77, 59–67. [Google Scholar] [CrossRef]
- Meador, J.P.; McCarty, L.S.; Escher, B.I.; Adams, W.J. 10th Anniversary Critical Review: The tissue-residue approach for toxicity assessment: Concepts, issues, application, and recommendations. J. Environ. Monit. 2008, 10, 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.J.; Blust, R.; Borgmann, U.; Brix, K.V.; DeForest, D.K.; Green, A.S.; Meyer, J.S.; McGeer, J.C.; Paquin, P.R.; Rainbow, P.S.; et al. Utility of tissue residues for predicting effects of metals on aquatic organisms. Integr. Environ. Assess. Manag. 2011, 7, 75–98. [Google Scholar] [CrossRef]
- Meador, J.P.; Adams, W.J.; Escher, B.I.; McCarty, L.S.; McElroy, A.E.; Sappington, K.G. The tissue residue approach for toxicity assessment: Findings and critical reviews from a Society of Environmental Toxicology and Chemistry Pellston Workshop. Integr. Environ. Assess. Manag. 2011, 7, 310. [Google Scholar] [CrossRef]
- Leonard, E.M.; Marentette, J.R.; Balshine, S.; Wood, C.M. Critical body residues, Michaelis–Menten analysis of bioaccumulation, lethality and behaviour as endpoints of waterborne Ni toxicity in two teleosts. Ecotoxicology 2014, 23, 147–162. [Google Scholar] [CrossRef]
- Wang, W.; Tan, Q. Applications of dynamic models in predicting the bioaccumulation, transport and toxicity of trace metals in aquatic organisms. Environ. Pollut. 2019, 252, 1561–1573. [Google Scholar] [CrossRef] [PubMed]
- Penttinen, S.; Malk, V.; Väisänen, A.; Penttinen, O.P. Using the critical body residue approach to determine the acute toxicity of cadmium at varying levels of water hardness and dissolved organic carbon concentrations. Ecotoxicol. Environ. Saf. 2011, 74, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Fernández, L.; Martínez-Madrid, M.; Rodriguez, P. Toxicity and critical body residues of Cd, Cu and Cr in the aquatic oligochaete Tubifex tubifex (Müller) based on lethal and sublethal effects. Ecotoxicology 2013, 22, 1445–1460. [Google Scholar] [CrossRef]
- Meador, J.P. Tissue concentrations as the dose metric to assess potential toxic effects of metals in field-collected fish: Copper and cadmium. Environ. Toxicol. Chem. 2015, 34, 1309–1319. [Google Scholar] [CrossRef] [Green Version]
- Delahaut, V.; Rašković, B.; Salvado, M.S.; Bervoets, L.; Blust, R.; De Boeck, G. Toxicity and bioaccumulation of Cadmium, Copper and Zinc in a direct comparison at equitoxic concentrations in common carp (Cyprinus carpio) juveniles. PLoS ONE 2020, 15, e220485. [Google Scholar] [CrossRef] [Green Version]
- Pilehvar, A.; Town, R.M.; Blust, R. The effect of thermal pre-incubation and exposure on sensitivity of zebrafish (Danio rerio) to copper and cadmium single and binary exposures. Aquat. Toxicol. 2019, 213, 105226. [Google Scholar] [CrossRef] [PubMed]
- Roch, M.; Maly, E.J. Relationship of Cadmium-Induced Hypocalcemia with Mortality in Rainbow Trout (Salmo gairdneri) and the Influence of Temperature on Toxicity. J. Fish. Res. Board Can. 1979, 36, 1297–1303. [Google Scholar] [CrossRef]
- Suresh, A.; Sivaramakrishna, B.; Radhakrishnaiah, K. Effect of lethal and sublethal concentrations of cadmium on energetics in the gills of fry and fingerlings of Cyprinus-carpio. Bull. Environ. Contam. Toxicol. 1993, 51, 920–926. [Google Scholar] [CrossRef] [PubMed]
- McGeer, J.C.; Szebedinszky, C.; McDonald, D.G.; Wood, C.M. Effects of chronic sublethal exposure to waterborne Cu, Cd or Zn in rainbow trout. 1: Iono-regulatory disturbance and metabolic costs. Aquat. Toxicol. 2000, 50, 231–243. [Google Scholar] [CrossRef]
- Rogers, J.T.; Richards, J.G.; Wood, C.M. Ionoregulatory disruption as the acute toxic mechanism for lead in the rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2003, 64, 215–234. [Google Scholar] [CrossRef]
- Alsop, D.; Wood, C.M. Metal and pharmaceutical mixtures: Is ion loss the mechanism underlying acute toxicity and widespread additive toxicity in zebrafish? Aquat. Toxicol. 2013, 140–141, 257–267. [Google Scholar] [CrossRef]
- Veltman, K.; Hendriks, A.J.; Huijbregts, M.A.J.; Wannaz, C.; Jolliet, O. Toxicokinetic Toxicodynamic (TKTD) Modeling of Ag Toxicity in Freshwater Organisms: Whole-Body Sodium Loss Predicts Acute Mortality Across Aquatic Species. Environ. Sci. Technol. 2014, 48, 14481–14489. [Google Scholar] [CrossRef]
- Castaldo, G.; Pillet, M.; Slootmaekers, B.; Bervoets, L.; Town, R.M.; Blust, R.; De Boeck, G. Investigating the effects of a sub-lethal metal mixture of Cu, Zn and Cd on bioaccumulation and ionoregulation in common carp, Cyprinus carpio. Aquat. Toxicol. 2020, 218, 105363. [Google Scholar] [CrossRef]
- Silva, A.O.F.D.; Martinez, C.B.R. Acute effects of cadmium on osmoregulation of the freshwater teleost Prochilodus lineatus: Enzymes activity and plasma ions. Aquat. Toxicol. 2014, 156, 161–168. [Google Scholar] [CrossRef]
- Shekh, K.; Tang, S.; Hecker, M.; Niyogi, S. Investigating the Role of Ionoregulatory Processes in the Species- and Life-Stage-Specific Differences in Sensitivity of Rainbow Trout and White Sturgeon to Cadmium. Environ. Sci. Technol. 2018, 52, 12868–12876. [Google Scholar] [CrossRef]
- McRae, N.K.; Gaw, S.; Glover, C.N. Effects of waterborne cadmium on metabolic rate, oxidative stress, and ion regulation in the freshwater fish, inanga (Galaxias maculatus). Aquat. Toxicol. 2018, 194, 1–9. [Google Scholar] [CrossRef]
- Xie, X.; Sun, R. The bioenergetics of the southern catfish (Silurus meridionah Chen): Growth rate as a function of ration level, body weight, and temperature. J. Fish. Biol. 1992, 40, 719–730. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, W.; Tang, R.; Li, L.; Refaey, M.M.; Li, D. Thermally processed diet greatly affects profiles of amino acids rather than fatty acids in the muscle of carnivorous Silurus meridionalis. Food Chem. 2018, 256, 244–251. [Google Scholar] [CrossRef]
- Wang, L.; Wang, H. Status and Strategy for Protection of Fishes in Changjiang River. J. Anhui Agric. Sci. 2011, 39, 12876–12877. [Google Scholar]
- Javed, M.; Abbas, S.; Latif, F. Bioaccumulation of Metals in Fish, Channa marulius, Mystus seenghala and Wallago attu during Acute Toxicity Exposures. Int. J. Agric. Biol. 2017, 19, 1566–1570. [Google Scholar] [CrossRef]
- Li, J.; Xie, X. Inconsistent responses of liver mitochondria metabolism and standard metabolism in Silurus meridionalis when exposed to waterborne cadmium. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 214, 17–22. [Google Scholar] [CrossRef]
- Li, J.; Yan, Y.; Xie, X. Tissue-Specific Antioxidative Responses and Cadmium Accumulation in Silurus meridionalis Under Chronic Waterborne Cadmium Exposure. Bull. Environ. Contam. Toxicol. 2018, 100, 485–491. [Google Scholar] [CrossRef]
- Li, J.; Xie, X. Effects of waterborne cadmium exposure on growth performance, specific dynamic action, and energy budget of southern catfish Silurus meridionalis. Aquaculture 2019, 511, 634194. [Google Scholar] [CrossRef]
- Chowdhury, M.J.; Baldisserotto, B.; Wood, C.M. Tissue-Specific Cadmium and Metallothionein Levels in Rainbow Trout Chronically Acclimated to Waterborne or Dietary Cadmium. Arch. Environ. Con. Toxicol. 2005, 48, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, M.; Gül, A.; Karaköse, E. Investigation of acute toxicity and the effect of cadmium chloride (CdCl2·H2O) metal salt on behavior of the guppy (Poecilia reticulata). Chemosphere 2004, 56, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, N.S.; Oliveira, L.H.B.; Agrelli, V.; de Oliveira, A.F.; Nogueira, A.R.A.; Oliveira, A.; Gonzalez, M.H. Bioaccumulation and acute toxicity of As(III) and As(V) in Nile tilapia (Oreochromis niloticus). Chemosphere 2019, 217, 349–354. [Google Scholar] [CrossRef]
- Rainbow, P.S. Trace metal bioaccumulation: Models, metabolic availability and toxicity. Environ. Int. 2007, 33, 576–582. [Google Scholar] [CrossRef] [PubMed]
- de Conto Cinier, C.; Faure, R.; Garin, D. Cadmium bioaccumulation in carp (Cyprinus carpio) tissues during long-term high exposure: Analysis by inductively coupled plasma-mass spectrometry. Ecotoxicol. Environ. Saf. 1997, 38, 137–143. [Google Scholar] [CrossRef]
- Olsson, P.E.; Larsson, A.; Maage, S.; Haux, C.; Bonham, K.; Zafarullah, M.; Gedamu, L. Induction of metallothionein synthesis in rainbow trout, Salmo gairdneri, during long-term exposure to water borne cadmium. Fish Physiol. Biochem. 1989, 6, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Gill, T.S.; Bianchi, C.P.; Epple, A. Trace metal (Cu and Zn) adaptation of organ systems of the American eel, Anguilla rostrata, to external concentrations of cadmium. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1992, 102, 361–371. [Google Scholar] [CrossRef]
- Mount, D.I.; Stephan, C.E. A method for detecting cadmium poisoning in fish. J. Wildl. Manag. 1967, 31, 168. [Google Scholar] [CrossRef]
- Xie, L.; Klerks, P.L. Changes in cadmium accumulation as a mechanism for cadmium resistance in the least killifish Heterandria formosa. Aquat. Toxicol. 2004, 66, 73–81. [Google Scholar] [CrossRef]
- Niimi, A.J.; Kissoon, G.P. Evaluation of the critical body burden concept based on inorganic and organic mercury toxicity to Rainbow Trout (Oncorhynchus mykiss). Arch. Environ. Contam. Toxicol. 1994, 26, 169–178. [Google Scholar] [CrossRef]
- Hollis, L.; Hogstrand, C.; Wood, C.M. Tissue-Specific Cadmium Accumulation, Metallothionein Induction, and Tissue Zinc and Copper Levels During Chronic Sublethal Cadmium Exposure in Juvenile Rainbow Trout. Arch. Environ. Contam. Toxicol. 2001, 41, 468–474. [Google Scholar] [CrossRef]
- Vijver, M.G.; van Gestel, C.A.M.; Lanno, R.P.; van Straalen, N.M.; Peijnenburg, W.J.G.M. Internal Metal Sequestration and Its Ecotoxicological Relevance: A Review. Environ. Sci. Technol. 2004, 38, 4705–4712. [Google Scholar] [CrossRef] [PubMed]
- McGeer, J.C.; Brix, K.V.; Skeaff, J.M.; DeForest, D.K.; Brigham, S.I.; Adams, W.J.; Green, A. Inverse relationship between bioconcentration factor and exposure concentration for metals: Implications for hazard assessment of metals in the aquatic environment. Environ. Toxicol. Chem. 2003, 22, 1017–1037. [Google Scholar] [CrossRef] [PubMed]
- DeForest, D.K.; Brix, K.V.; Adams, W.J. Assessing metal bioaccumulation in aquatic environments: The inverse relationship between bioaccumulation factors, trophic transfer factors and exposure concentration. Aquat. Toxicol. 2007, 84, 236–246. [Google Scholar] [CrossRef]
- Gupta, A.K.; Rajbanshi, V.K. Acute Toxicity of Cadmium to Channa punctutus (Bloch). Acta Hydrochim. Hydrobiol. 1988, 5, 525–535. [Google Scholar] [CrossRef]
- Paul, J.S.; Small, B.C. Chronic exposure to environmental cadmium affects growth and survival, cellular stress, and glucose metabolism in juvenile channel catfish (Ictalurus punctatus). Aquat. Toxicol. 2021, 230, 105705. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, A.; Wenstel, R.; Sappington, K.; Wood, W. Framework for metals risk assessment. Ecotoxicol. Environ. Saf. 2007, 68, 145–227. [Google Scholar] [CrossRef]
- Milligan, C.L.; Wood, C.M. Disturbances in hematology, fluid volume distribution, and circulatory function associated with low environmental pH in the rainbow trout, Salmo gairdneri. J. Exp. Biol. 1982, 99, 397–415. [Google Scholar] [CrossRef]
- Wilson, R.; Taylor, E. The physiological responses of freshwater rainbow trout, Oncorhynchus mykiss, during acutely lethal copper exposure. J. Comp. Physiol. B 1993, 163, 38–47. [Google Scholar] [CrossRef]
- Pilehvar, A.; Cordery, K.I.; Town, R.M.; Blust, R. The synergistic toxicity of Cd(II) and Cu(II) to zebrafish (Danio rerio): Effect of water hardness. Chemosphere 2020, 247, 125942. [Google Scholar] [CrossRef]
- Playle, R.C.; Dixon, D.G. Copper and Cadmium Binding to Fish Gill: Estimates of metal-gill stability constants and modelling of metal accumulation. Can. J. Fish. Aqucrt. Sci. 1993, 50, 2678–2687. [Google Scholar] [CrossRef]
- Verbost, P.M.; Flik, G.; Lock, R.A.C.; Wendelaar Bonga, S.E. Cadmium inhibits plasma membrane calcium transport. J. Membr. Biol. 1988, 102, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Suresh, A.; Sivaramakrishna, B.; Radhakrishnaiah, K. Cadmium induced changes in ion levels and ATPase activities in the muscle of the fry and fingerlings of the freshwater fish, Cyprinus carpio. Chemosphere 1995, 30, 367–375. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Qiu, H.; Yan, Y.; Xie, X. Acute Cd Toxicity, Metal Accumulation, and Ion Loss in Southern Catfish (Silurus meridionalis Chen). Toxics 2021, 9, 202. https://doi.org/10.3390/toxics9090202
Liu W, Qiu H, Yan Y, Xie X. Acute Cd Toxicity, Metal Accumulation, and Ion Loss in Southern Catfish (Silurus meridionalis Chen). Toxics. 2021; 9(9):202. https://doi.org/10.3390/toxics9090202
Chicago/Turabian StyleLiu, Wenming, Hanxun Qiu, Yulian Yan, and Xiaojun Xie. 2021. "Acute Cd Toxicity, Metal Accumulation, and Ion Loss in Southern Catfish (Silurus meridionalis Chen)" Toxics 9, no. 9: 202. https://doi.org/10.3390/toxics9090202
APA StyleLiu, W., Qiu, H., Yan, Y., & Xie, X. (2021). Acute Cd Toxicity, Metal Accumulation, and Ion Loss in Southern Catfish (Silurus meridionalis Chen). Toxics, 9(9), 202. https://doi.org/10.3390/toxics9090202