Effect of 10 UV Filters on the Brine Shrimp Artemia salina and the Marine Microalga Tetraselmis sp.

 and
and 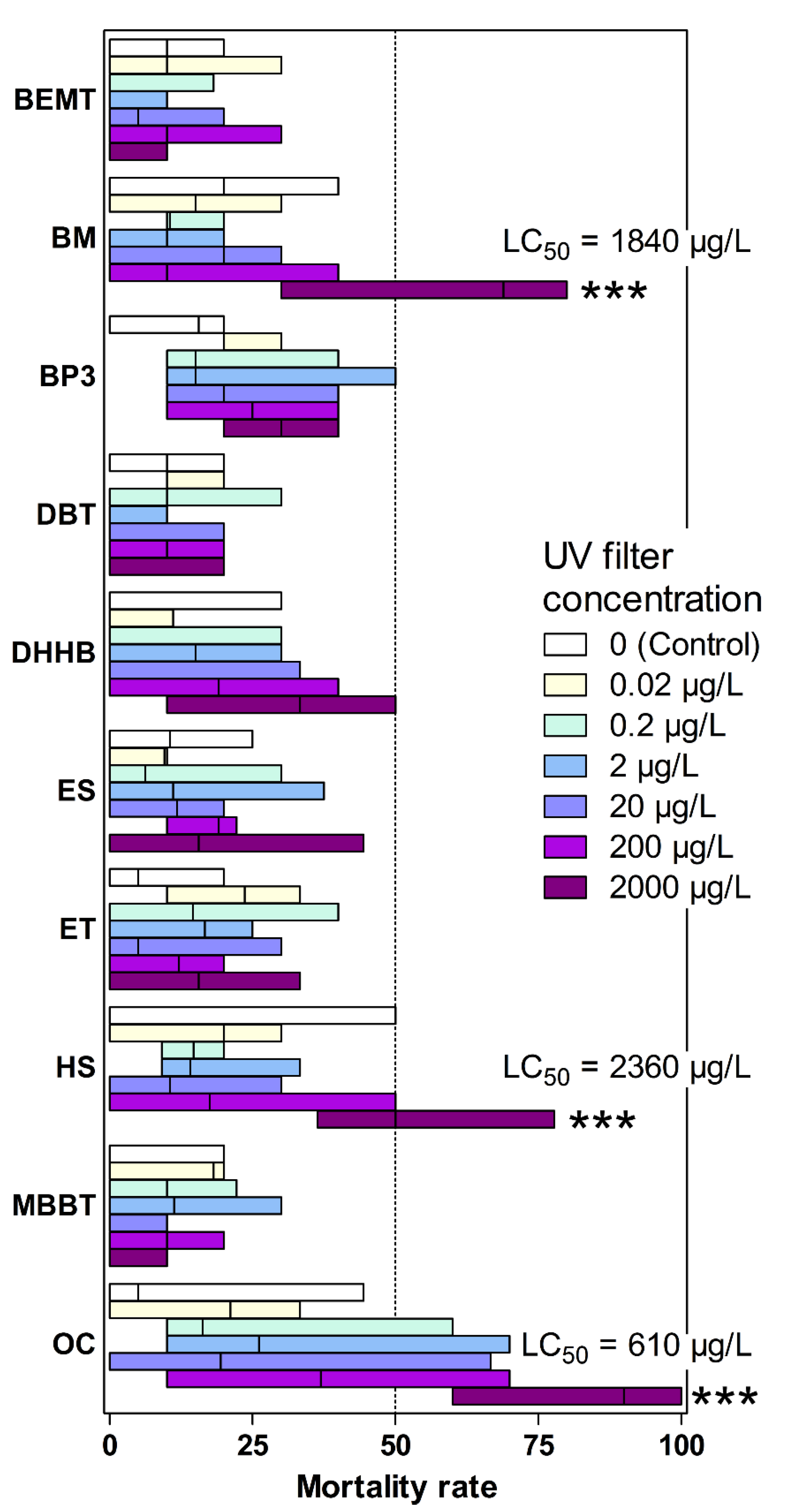{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and methods
2.1. Test Substances and Experimental Solutions
2.2. Artemia salina Mortality Test
2.3. Tetraselmis sp. Toxicity Test
2.3.1. Experimental Procedure
2.3.2. Flow Cytometry (FCM) Analyses
2.4. Statistical Analysis
3. Results and Discussion
3.1. Effects on Artemia salina Mortality
3.2. Effects on Tetraselmis sp.
3.2.1. Growth Rate and EC50 Values
3.2.2. Impact on Cell Morphology
3.2.3. Impact on Autofluorescence
3.2.4. Impact on Cell Metabolic Activity
4. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azoury, S.C.; Lange, J.R. Epidemiology, risk factors, prevention, and early detection of melanoma. Surg. Clin. 2014, 94, 945–962. [Google Scholar] [CrossRef] [PubMed]
- Waldman, R.A.; Grant-Kels, J.M. The role of sunscreen in the prevention of cutaneous melanoma and nonmelanoma skin cancer. J. Am. Acad. Dermatol. 2019, 80, 574–576.e1. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Quiles, D.; Tovar-Sánchez, A. Are sunscreens a new environmental risk associated with coastal tourism? Environ. Int. 2015, 83, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Pirotta, G. The encyclopedia of allowed sunfilters in the world. Skin Care—Househ. Pers. Care Today 2016, 11, 19–21. [Google Scholar]
- Tsui, M.M.P.; Leung, H.W.; Wai, T.-C.; Yamashita, N.; Taniyasu, S.; Liu, W.; Lam, P.K.S.; Murphy, M.B. Occurrence, distribution and ecological risk assessment of multiple classes of UV filters in surface waters from different countries. Water Res. 2014, 67, 55–65. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, A.; Rodrigo Sanz, M. Betancourt Rodriguez, J.R. Occurrence of eight UV filters in beaches of Gran Canaria (Canary Islands). An approach to environmental risk assessment. Chemosphere 2015, 131, 85–90. [Google Scholar] [CrossRef]
- Lozano, C.; Givens, J.; Stien, D.; Matallana-Surget, S.; Lebaron, P. Bioaccumulation and Toxicological Effects of UV-Filters on Marine Species. In The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]
- Downs, C.A.; Kramarsky-Winter, E.; Segal, R.; Fauth, J.; Knutson, S.; Bronstein, O.; Ciner, F.R.; Jeger, R.; Lichtenfeld, Y.; Woodley, C.M.; et al. Toxicopathological effects of the sunscreen UV filter, oxybenzone (benzophenone-3), on coral planulae and cultured primary cells and its environmental contamination in Hawaii and the U.S. Virgin Islands. Arch. Environ. Contam. Toxicol. 2016, 70, 265–288. [Google Scholar] [CrossRef]
- Giokas, D.L.; Salvador, A.; Chisvert, A. UV filters: From sunscreen to human body and the environment. TrAC Trends Anal. Chem. 2007, 26, 360–374. [Google Scholar] [CrossRef]
- Gago-Ferrero, P.; Díaz-Cruz, M.S.; Barceló, D. UV filters bioaccumulation in fish from Iberian river basins. Sci. Total Environ. 2015, 518, 518–525. [Google Scholar] [CrossRef]
- Vidal-Liñán, L.; Villaverde-de-Sáa, E.; Rodil, R.; Quintana, J.B.; Beiras, R. Bioaccumulation of UV filters in Mytilus galloprovincialis mussel. Chemosphere 2018, 190, 267–271. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Hong, H.; Shi, D. Multigenerational effects of 4-methylbenzylidene camphor (4-MBC) on the survival, development and reproduction of the marine copepod Tigriopus japonicus. Aquat. Toxicol. 2018, 194, 94–102. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Tsui, M.M.P.; Tan, C.J.; Ma, C.Y.; Yiu, S.K.F.; Wang, L.H.; Chen, T.H.; Fan, T.Y.; Lam, P.K.S.; Murphy, M.B.; et al. Toxicological effects of two organic ultraviolet filters and a related commercial sunscreen product in adult corals. Environ. Pollut. 2019, 245, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Paredes, E.; Perez, S.; Rodil, R.; Quintana, J.B.; Beiras, R. Ecotoxicological evaluation of four UV filters using marine organisms from different trophic levels Isochrysis galbana, Mytilus galloprovincialis, Paracentrotus lividus, and Siriella armata. Chemosphere 2014, 104, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.J.; Rocha, R.J.M.; Soares, A.M.V.M.; Benedé, J.L.; Chisvert, A.; Monteiro, M.S. Effects of UV filter 4-methylbenzylidene camphor during early development of Solea senegalensis Kaup, 1858. Sci. Total Environ. 2018, 628–629, 1395–1404. [Google Scholar] [CrossRef]
- Barone, A.N.; Hayes, C.E.; Kerr, J.J.; Lee, R.C.; Flaherty, D.B. Acute toxicity testing of TiO2-based vs. oxybenzone-based sunscreens on clownfish (Amphiprion ocellaris). Environ. Sci. Pollut. Res. 2019, 26, 14513–14520. [Google Scholar] [CrossRef]
- Mao, F.; He, Y.; Kushmaro, A.; Gin, K.Y.-H. Effects of benzophenone-3 on the green alga Chlamydomonas reinhardtii and the cyanobacterium Microcystis aeruginosa. Aquat. Toxicol. 2017, 193, 1–8. [Google Scholar] [CrossRef]
- Sieratowicz, A.; Kaiser, D.; Behr, M.; Oetken, M.; Oehlmann, J. Acute and chronic toxicity of four frequently used UV filter substances for Desmodesmus subspicatus and Daphnia magna. J. Environ. Sci. Health Part A 2011, 46, 1311–1319. [Google Scholar] [CrossRef]
- Giraldo, A.; Montes, R.; Rodil, R.; Quintana, J.B.; Vidal-Liñán, L.; Beiras, R. Ecotoxicological evaluation of the UV filters ethylhexyl dimethyl p-aminobenzoic acid and octocrylene using marine organisms Isochrysis galbana, Mytilus galloprovincialis and Paracentrotus lividus. Arch. Environ. Contam. Toxicol. 2017, 72, 606–611. [Google Scholar] [CrossRef]
- Torres, T.; Cunha, I.; Martins, R.; Santos, M.M. Screening the toxicity of selected personal care products using embryo bioassays: 4-MBC, propylparaben and triclocarban. Int. J. Mol. Sci. 2016, 17, 1762. [Google Scholar] [CrossRef]
- Esperanza, M.; Seoane, M.; Rioboo, C.; Herrero, C.; Cid, Á. Differential toxicity of the UV-filters BP-3 and BP-4 in Chlamydomonas reinhardtii: A flow cytometric approach. Sci. Total Environ. 2019, 669, 412–420. [Google Scholar] [CrossRef]
- Seoane, M.; Rioboo, C.; Herrero, C.; Cid, Á. Toxicity induced by three antibiotics commonly used in aquaculture on the marine microalga Tetraselmis suecica (Kylin) Butch. Mar. Environ. Res. 2014, 101, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Stien, D.; Clergeaud, F.; Rodrigues, A.M.S.; Lebaron, K.; Pillot, R.; Romans, P.; Fagervold, S.; Lebaron, P. Metabolomics reveal that octocrylene accumulates in Pocillopora damicornis tissues as fatty acid conjugates and triggers coral cell mitochondrial dysfunction. Anal. Chem. 2019, 91, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Yuan, T.; Zhou, C.; Cheng, P.; Bai, Q.; Ao, J.; Wang, W.; Zhang, H. Effects of four commonly used UV filters on the growth, cell viability and oxidative stress responses of the Tetrahymena thermophila. Chemosphere 2013, 93, 2507–2513. [Google Scholar] [CrossRef] [PubMed]
- Zucchi, S.; Oggier, D.M.; Fent, K. Global gene expression profile induced by the UV-filter 2-ethyl-hexyl-4-trimethoxycinnamate (EHMC) in zebrafish (Danio rerio). Environ. Pollut. 2011, 159, 3086–3096. [Google Scholar] [CrossRef] [PubMed]
- Coronado, M.; De Haro, H.; Deng, X.; Rempel, M.A.; Lavado, R.; Schlenk, D. Estrogenic activity and reproductive effects of the UV-filter oxybenzone (2-hydroxy-4-methoxyphenyl-methanone) in fish. Aquat. Toxicol. 2008, 90, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, D.; Sieratowicz, A.; Zielke, H.; Oetken, M.; Hollert, H.; Oehlmann, J. Ecotoxicological effect characterisation of widely used organic UV filters. Environ. Pollut. 2012, 163, 84–90. [Google Scholar] [CrossRef]
- European Commission. Chemicals Legislation | Internal Market, Industry, Entrepreneurship and SMEs. 2007. Available online: https://ec.europa.eu/growth/sectors/chemicals/legislation_en (accessed on 12 March 2019).
- Caldwell, G.S.; Bentley, M.G.; Olive, P.J.W. The use of a brine shrimp (Artemia salina) bioassay to assess the toxicity of diatom extracts and short chain aldehydes. Toxicon 2003, 42, 301–306. [Google Scholar] [CrossRef]
- Libralato, G. The case of Artemia spp. in nanoecotoxicology. Mar. Environ. Res. 2014, 101, 38–43. [Google Scholar] [CrossRef]
- Nunes, B.S.; Carvalho, F.D.; Guilhermino, L.M.; Van Stappen, G. Use of the genus Artemia in ecotoxicity testing. Environ. Pollut. 2006, 144, 453–462. [Google Scholar] [CrossRef]
- Pecqueur, D.; Vidussi, F.; Fouilland, E.; Le Floc’h, E.; Mas, S.; Roques, C.; Salles, C.; Tournoud, M.-G.; Mostajir, B. Dynamics of microbial planktonic food web components during a river flash flood in a Mediterranean coastal lagoon. Hydrobiologia 2011, 673, 13–27. [Google Scholar] [CrossRef]
- Parthuisot, N.; Catala, P.; Lemarchand, K.; Baudart, J.; Lebaron, P. Evalautionof ChemChrome V6 for bactierial viability assessment in waters. J. Appl. Microbiol. 2000, 89, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Fagervold, S.K.; Rodrigues, A.M.S.; Rohée, C.; Roe, R.; Bourrain, M.; Stien, D.; Lebaron, P. Occurrence and environmental distribution of 5 UV filters during the summer season in different water bodies. Water Air Soil Pollut. 2019, 230, 172. [Google Scholar] [CrossRef]
- Ramos, S.; Homem, V.; Alves, A.; SAntos, L. Advances in analytical methods and occuence of organic UV-filters in the environment—A review. Sci. Total. Environ. 2015, 526, 278–311. [Google Scholar] [CrossRef] [PubMed]
- Langford, K.H.; Thomas, K.V. Inputs of chemicals from recreational activities into the Norwegian coastal zone. J. Environ. Monit. 2008, 10, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Blüthgen, N.; Meili, N.; Chew, G.; Odermatt, A.; Fent, K. Accumulation and effects of the UV-filter octocrylene in adult and embryonic zebrafish (Danio rerio). Sci. Total Environ. 2014, 476, 476–477. [Google Scholar] [CrossRef]
- Gago-Ferrero, P.; Díaz-Cruz, M.S.; Barceló, D. Fast pressurized liquid extraction with in-cell purification and analysis by liquid chromatography tandem mass spectrometry for the determination of UV filters and their degradation products in sediments. Anal. Bioanal. Chem. 2011, 400, 2195–2204. [Google Scholar] [CrossRef]
- Kameda, Y.; Kimura, K.; Miyazaki, M. Occurrence and profiles of organic sun-blocking agents in surface waters and sediments in Japanese rivers and lakes. Environ. Pollut. 2011, 159, 1570–1576. [Google Scholar] [CrossRef]
- Sankoda, K.; Murata, K.; Tanihata, M.; Suzuki, K.; Nomiyama, K.; Shinohara, R. Seasonal and Diurnal variation of organic ultraviolet filters from personal care products used along the Japanese coast. Arch. Environ. Contam. Toxicol. 2015, 68, 217–224. [Google Scholar] [CrossRef]
- Pablos, M.V.; García-Hortigüela, P.; Fernández, C. Acute and chronic toxicity of emerging contaminants, alone or in combination, in Chlorella vulgaris and Daphnia magna. Environ. Sci. Pollut. Res. 2015, 22, 5417–5424. [Google Scholar] [CrossRef]
- Zhong, X.; Downs, C.A.; Che, X.; Zhang, Z.; Li, Y.; Liu, B.; Li, Q.; Li, Y.; Gao, H. The toxicological effects of oxybenzone, an active ingredient in suncream personal care products, on prokaryotic alga Arthrospira sp. and eukaryotic alga Chlorella sp. Aquat. Toxicol. 2019, 216, 105295. [Google Scholar] [CrossRef]
- Seoane, M.; Esperanza, M.; Rioboo, C.; Herrero, C.; Cid, Á. Flow cytometric assay to assess short-term effects of personal care products on the marine microalga Tetraselmis suecica. Chemosphere 2017, 171, 339–347. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thorel, E.; Clergeaud, F.; Jaugeon, L.; Rodrigues, A.M.S.; Lucas, J.; Stien, D.; Lebaron, P. Effect of 10 UV Filters on the Brine Shrimp Artemia salina and the Marine Microalga Tetraselmis sp. Toxics 2020, 8, 29. https://doi.org/10.3390/toxics8020029
Thorel E, Clergeaud F, Jaugeon L, Rodrigues AMS, Lucas J, Stien D, Lebaron P. Effect of 10 UV Filters on the Brine Shrimp Artemia salina and the Marine Microalga Tetraselmis sp. Toxics. 2020; 8(2):29. https://doi.org/10.3390/toxics8020029
Chicago/Turabian StyleThorel, Evane, Fanny Clergeaud, Lucie Jaugeon, Alice M. S. Rodrigues, Julie Lucas, Didier Stien, and Philippe Lebaron. 2020. "Effect of 10 UV Filters on the Brine Shrimp Artemia salina and the Marine Microalga Tetraselmis sp." Toxics 8, no. 2: 29. https://doi.org/10.3390/toxics8020029
APA StyleThorel, E., Clergeaud, F., Jaugeon, L., Rodrigues, A. M. S., Lucas, J., Stien, D., & Lebaron, P. (2020). Effect of 10 UV Filters on the Brine Shrimp Artemia salina and the Marine Microalga Tetraselmis sp. Toxics, 8(2), 29. https://doi.org/10.3390/toxics8020029