Chlorpyrifos- and Dichlorvos-Induced Oxidative and Neurogenic Damage Elicits Neuro-Cognitive Deficits and Increases Anxiety-Like Behavior in Wild-Type Rats

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Drugs
2.2. Animals and Experimental Design
2.3. Treatment Schedule
- Group 1 (control)—given normal saline (1 mL/kg orally) daily for 14 days
- Group 3—given CPF (14.9 mg/kg orally) daily for 14 days [9]
- Group 4—given DDVP (8.8 mg/kg orally) plus CPF (14 mg/kg orally) daily for 14 days
2.4. Ethical Approval
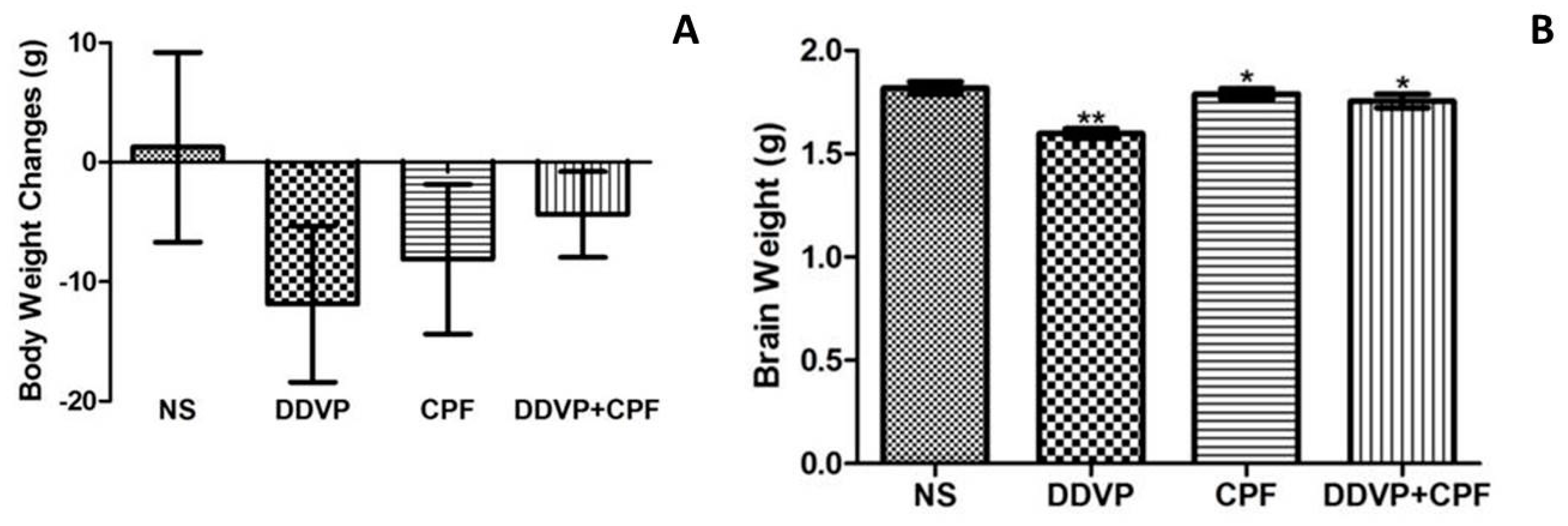
2.5. Evaluationof Brain and Body Weight
2.6. Behavioral Evaluations
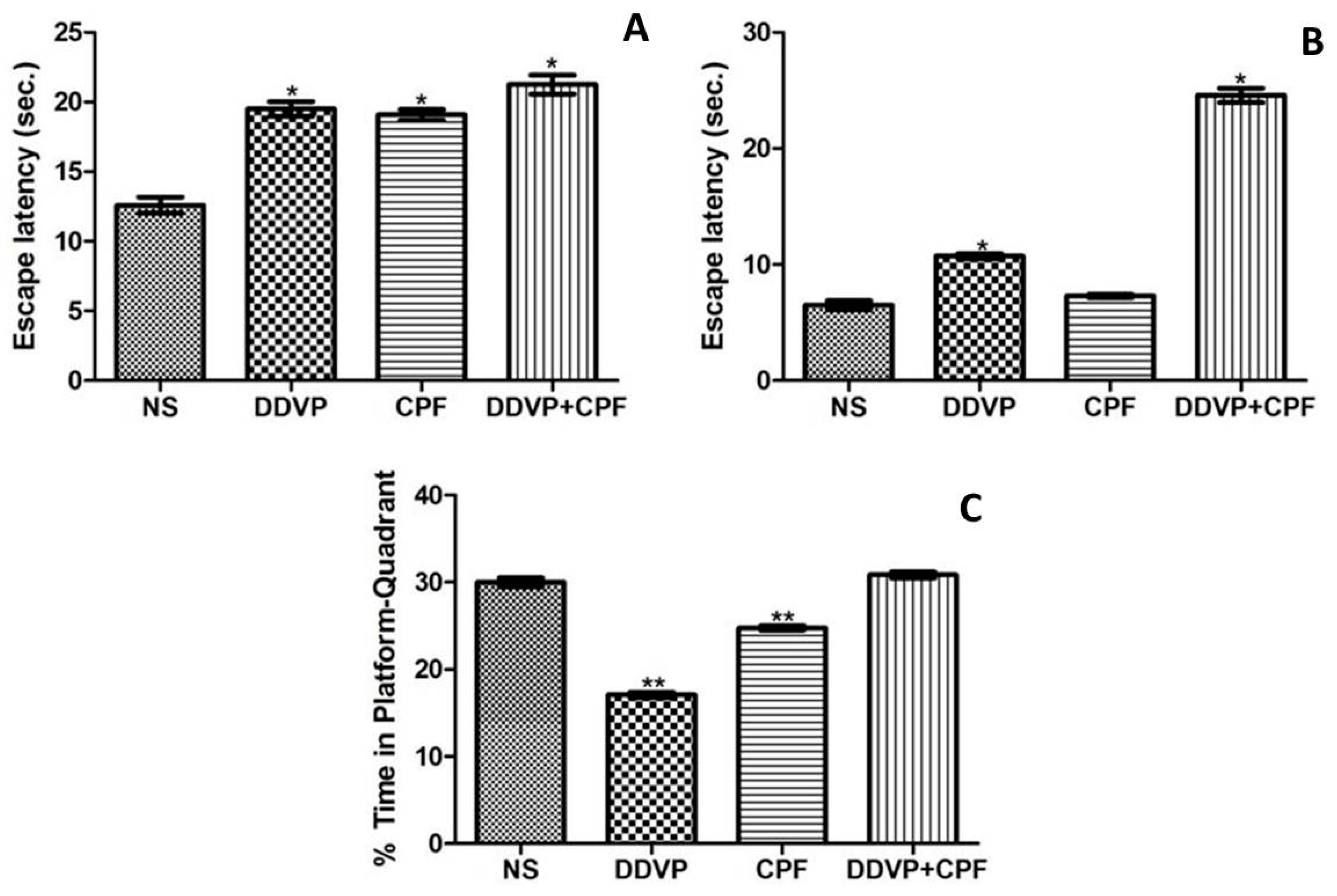
2.6.1. Morris Water Maze Procedure
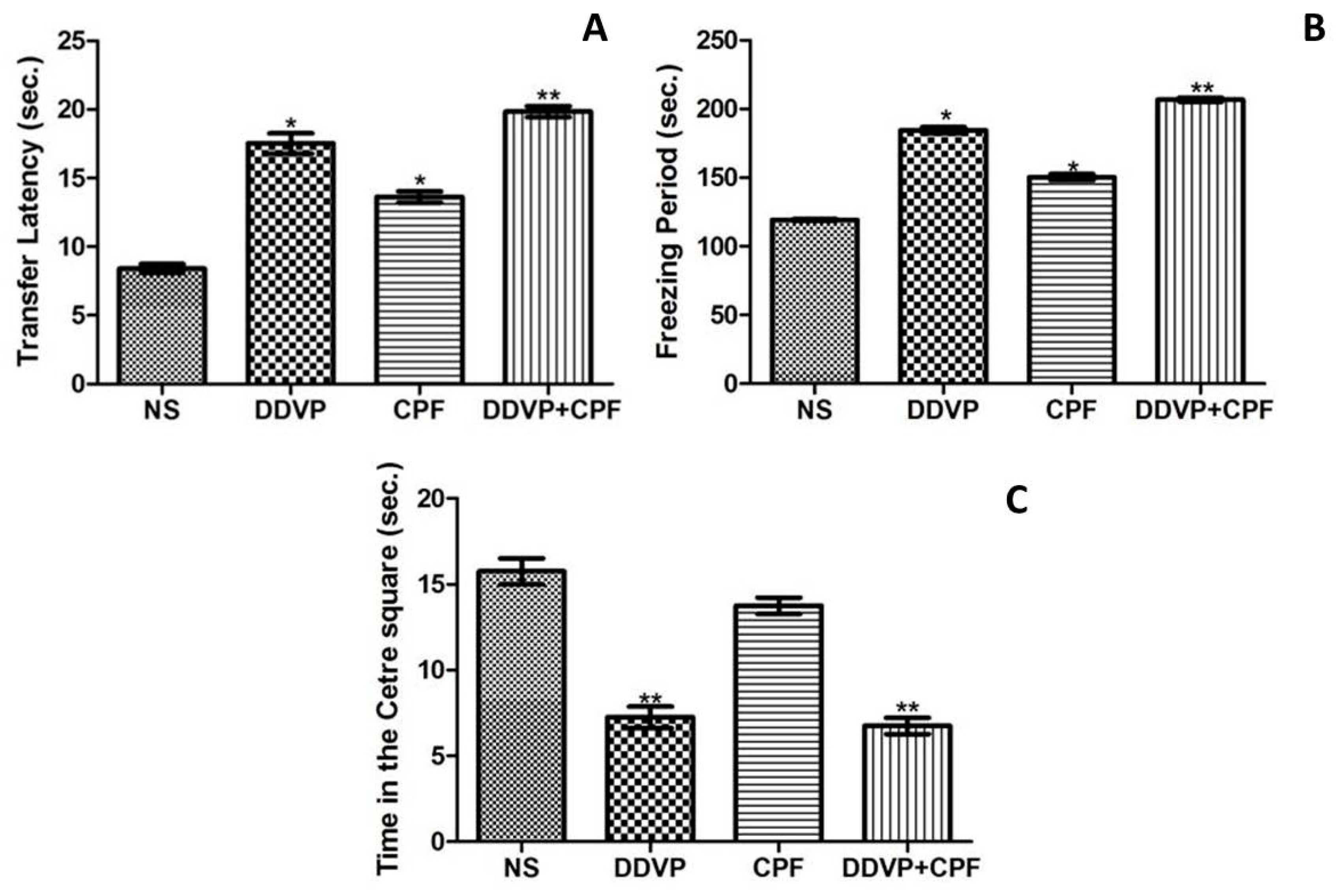
2.6.2. Anxiety-Like Behaviors and Fear Learning
OFT Procedure
EPM Procedure
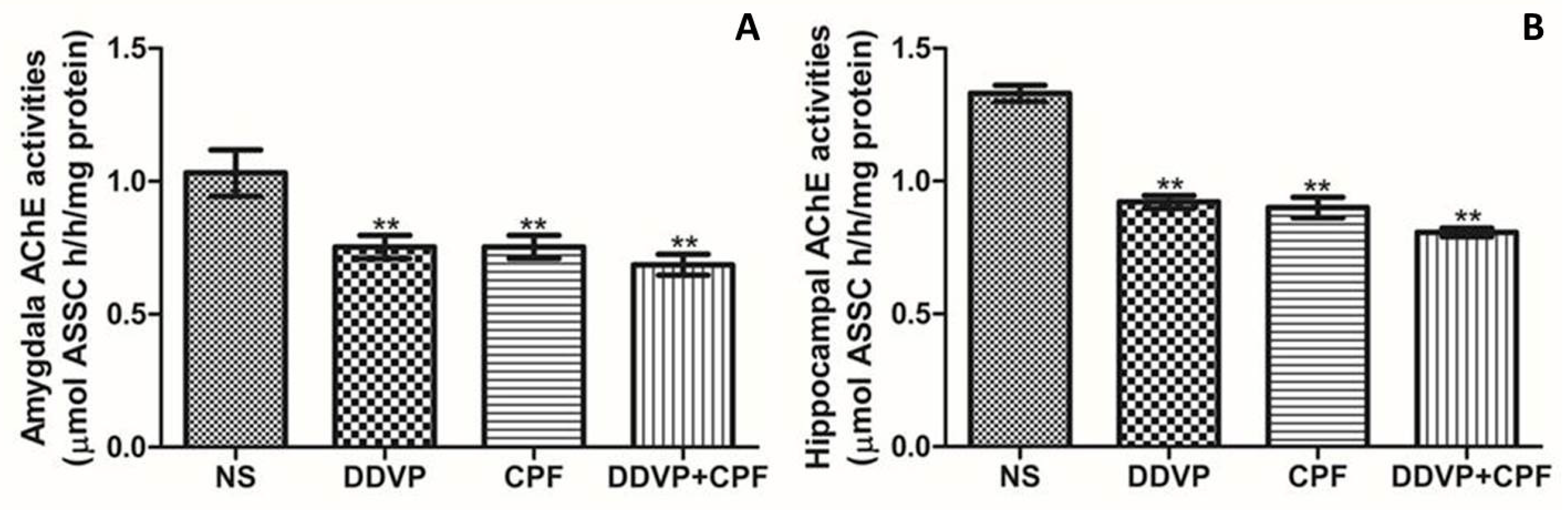
2.7. Biochemical Evaluation
2.8. Tissue Processing and Histopathology
Immunohistochemistry for Ki-67
2.9. Statistical Analysis
3. Results
3.1. Morphometric Changes Following Exposure to DDVP and CPF
3.2. Effects of DDVP and CPF Exposures on Spatial Working Memory
3.3. DDVP and CPF Exposures Increased Anxiety-Like Behavious
3.4. DDVP and CPF Exposures Inhibit Anticholinesterase in the Amygdala and Hippocampus
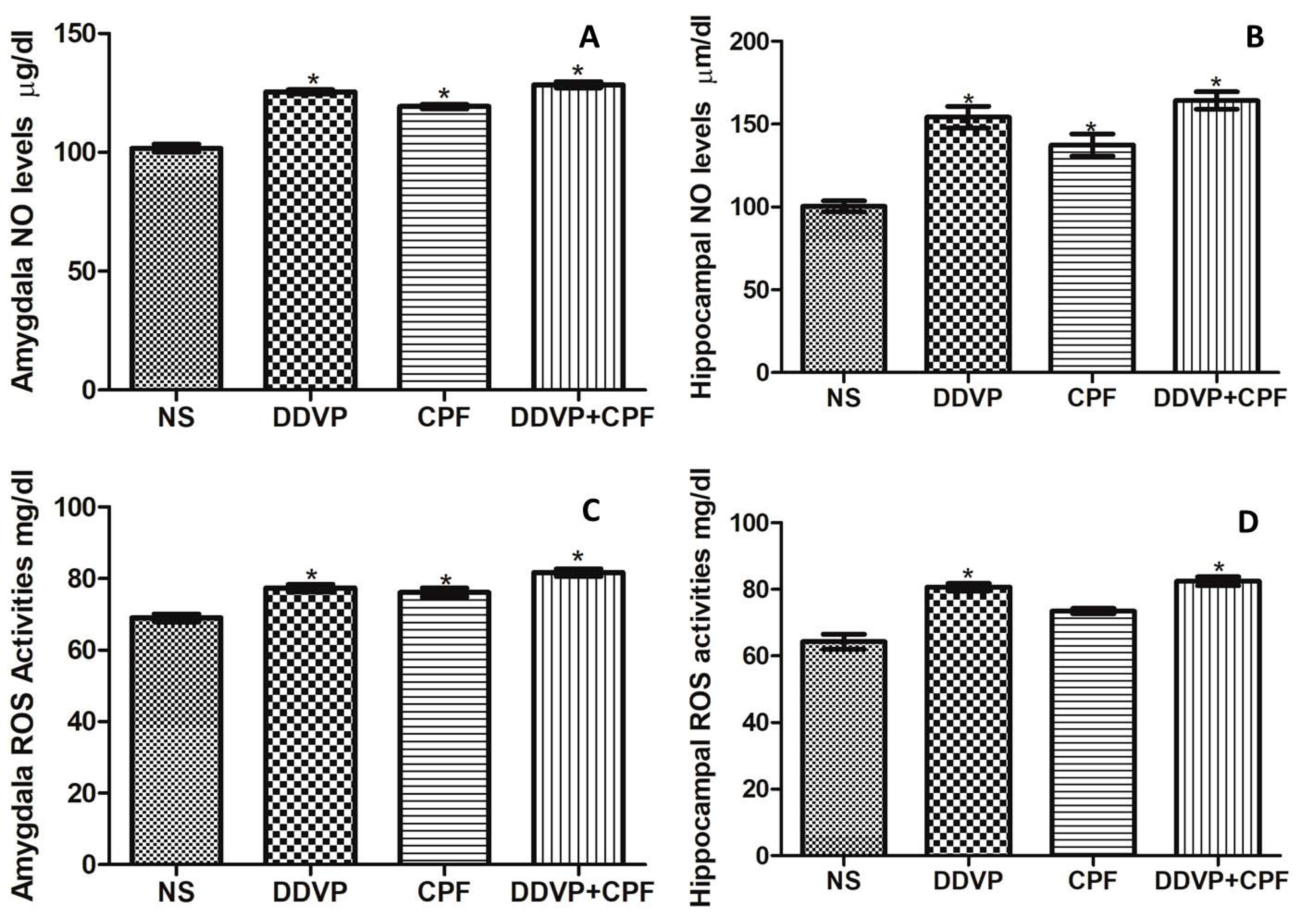
3.5. Effects of DDVP and CPF Exposures on Oxidative Stress Markers (ROS and NO) in the Amygdala and Hippocampus
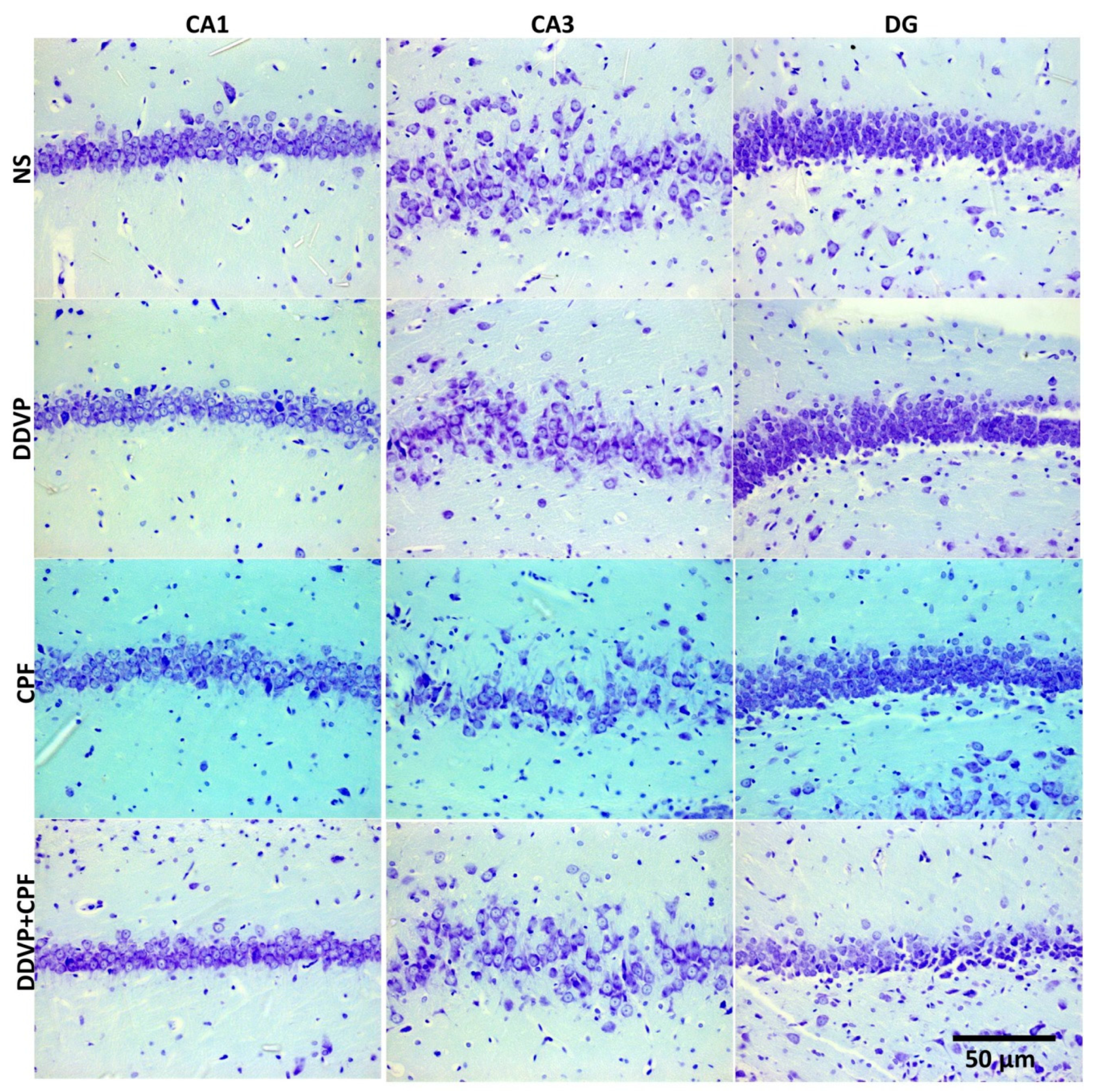
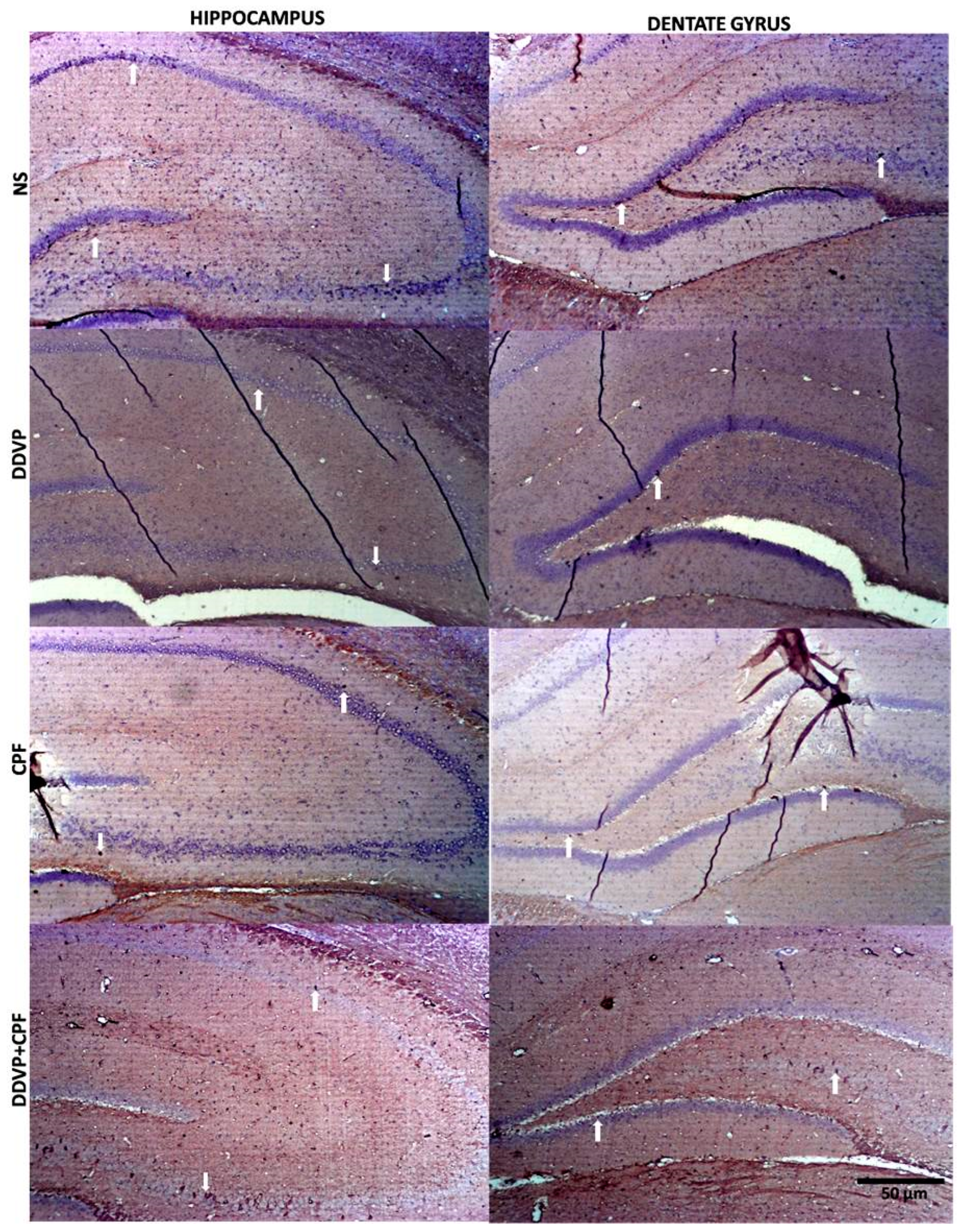
3.6. Effects of DDVP and CPF Exposures on the Distributions of Proliferative Nuclei (Ki67) in the Hippocampus and the Histoarchitecture
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Butterfield, D.A.; Lauderback, C.M. Lipid peroxidation and protein oxidation in Alzheimer’s disease brain: Potential causes and consequences involving amyloid beta-peptide-associated free radical oxidative stress. Free Radic. Biol. Med. 2002, 32, 1050–1060. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rainba, A.; Shadnia, S.; Nikfar, S.; Rezaie, A. Pesticide and oxidative stress: A review. Med. Sci. Monit. 2004, 10, RA141–RA147. [Google Scholar] [PubMed]
- Mostafalou, S.; Abdollahi, M. Pesticides and human chronic diseases: Evidences, mechanisms, and perspectives. Toxicol. Appl. Pharmacol. 2013, 268, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska-Trypuć, A. Pesticides as inducers of oxidative stress. React. Oxyg. Species 2017, 3, 96–110. [Google Scholar] [CrossRef]
- Pereira, E.F.; Aracava, Y.; DeTolla, L.J.; Beecham, E.J.; Basinger, G.W.; Wakayama, E.J.; Albuquerque, E.X. Animal models that best reproduce the clinical manifestations of human intoxication with organophosphorus compounds. J. Pharmacol. Exp. Ther. 2014, 350, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.D.; Todd, S.W.; Lumsden, E.; Mullins, R.J.; Mamczarz, J.; Fawcett, W.P.; Gullapalli, R.P.; Randall, W.R.; Pereira, E.F.R.; Albuquerque, E.X. Developmental neurotoxicity of the organophosphorus insecticide chlorpyrifos: From clinical findings to preclinical models and potential mechanisms. J. Neurochem. 2017, 142, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G. Organophosphorus compounds at 80: Some old and new issues. Toxicol. Sci. 2018, 162, 24–35. [Google Scholar] [CrossRef]
- Imam, A.; Ogunniyi, A.; Ibrahim, A.; Abdulmajeed, W.I.; Oyewole, L.A.; Lawan, A.H.; Sulaimon, F.A.; Adana, M.Y.; Ajao, M.S. Dichlorvos induced Oxidative and Neuronal responses in rats: Mitigative Efficacy of Nigella sativa (black cumin). Niger. J. Physiol. Sci. 2018, 33, 83–88. [Google Scholar]
- Imam, A.; Sulaiman, N.A.; Oyewole, A.L.; Amin, A.; Shittu, S.-T.T.; Ajao, M.S. Pro-Neurogenic and Antioxidant Efficacy of Nigella sativa Oil reduced Vulnerability to Cholinesterase Dysfunction and Disruption in Amygdala Dependent Behaviors in CPF Exposure. JKIMSU 2018, 7, 1–12. [Google Scholar]
- Pearson, J.N.; Patel, M. The role of oxidative stress in organophosphate and nerve agent toxicity. Ann. N. Y. Acad. Sci. 2016, 1378, 17–24. [Google Scholar] [CrossRef]
- Wang, X.; Martínez, M.A.; Dai, M.; Chen, D.; Ares, I.; Romero, A.; Castellano, V.; Martínez, M.; Rodríguez, J.L.; Martínez-Larrañaga, M.R.; et al. Permethrin-induced oxidative stress and toxicity and metabolism. A review. Environ. Res. 2016, 149, 86–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Anadón, A.; Wu, Q.; Qiao, F.; Ares, I.; Martínez-Larrañaga, M.R.; Yuan, Z.; Martínez, M.A. Mechanism of neonicotinoid toxicity: Impact on oxidative stress and metabolism. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 471–507. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, T.H.; Apland, J.P.; Braga, M.F.M.; Marini, A.M. Acute and long-term consequences of exposure to organophosphate nerve agents in humans. Epilepsia 2018, 59, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Harrison, V.; Mackenzie Ross, S.J. An emerging concern: Toxic fumes in airplane cabins. Cortex 2016, 74, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, S.; Burdon, J.; Vyvyan Howard, C. Aerotoxic Syndrome: A New Occupational Disease? Public Health Panorama 2017, 3, 198–211. [Google Scholar]
- Fatma, M.E. Lipid peroxidation, oxidative stress and acetylcholinesterase in rat brain exposed to organophosphate and pyrethroid insecticides. Food Chem. Toxicol. 2011, 49, 1346–1352. [Google Scholar] [CrossRef]
- Kodali, M.; Hattiangady, B.; Shetty, G.A.; Bates, A.; Shuai, B.; Shetty, A.K. Curcumin treatment leads to better cognitive and mood function in a model of Gulf War Illness with enhanced neurogenesis, and alleviation of inflammation and mitochondrial dysfunction in the hippocampus. Brain Behav. Immun. 2018, 69, 499–514. [Google Scholar] [CrossRef]
- Sánchez-Santed, F.; Colomina, M.T.; Herrero Hernández, E. Organophosphate pesticide exposure and neurodegeneration. Cortex 2016, 74, 417–426. [Google Scholar] [CrossRef]
- Yan, D.; Zhang, Y.; Liu, L.; Yan, H. Pesticide exposure and risk of Alzheimer’s disease: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 32222. [Google Scholar] [CrossRef]
- Ajao, M.S.; Adebayo, S.; Imam, A.; Ibrahim, A.; Adana, M.Y.; Alli-Oluwafuyi, A.; Kareem, S.B. Protective Effect of Nigella Sativa (Black Caraway) Oil on Oral Dichlorrvos Induced Hematological, Renal and Nonspecific Immune System Toxicity in Wistar rats. Iran. J. Toxic. 2017, 11, 1–5. [Google Scholar] [CrossRef]
- Ajao, M.S.; Abdussalam, W.A.; Imam, A.; Amin, A.; Ibrahim, A.; Adana, M.A.; Sulaimon, F.A.; Atata, J.A. Histopathological and Biochemical evaluations of the antidotal efficacy y of Nigella sativa oil on organophosphate induced hepatotoxicity. Res. J. Health Sci. 2017, 5, 18–20. [Google Scholar] [CrossRef][Green Version]
- Kingsley, C.K.; Solomon, N.I.; Odudu, A. Haematological, Biochemical and Antioxidant Changes in Wistar Rats Exposed to Dichlorvos Based Insecticide Formulation Used in Southeast Nigeria. Toxics 2016, 4, 28. [Google Scholar] [CrossRef]
- Arthur, S.; Ran, M.; Sigal, E.; Noah, L.-C.; Yossi, R.; Shai, S.; Michael, K.; Amos, K.; Yoram, E. QT Prolongation as an Isolated Long-Term Cardiac Manifestation of Dichlorvos Organophosphate Poisoning in Rats. Cardiovasc. Toxicol. 2017. [Google Scholar] [CrossRef]
- Suleiman, F.A.; Annas, O.M.; Muftau, S.; Stephen, A.A.; Favour, O.M. Alleviating Effect of Phyllanthusniruri on Sensorimotor and Cognitive Changes Induced by SubacuteChlorpyrifos Exposure in Wistar Rats. Am. J. Med. Med. Sci. 2012, 2, 50–58. [Google Scholar] [CrossRef]
- Hayden, K.M.; Norton, M.C.; Darcey, D.; Østbye, T.; Zandi, P.P.; Breitner, J.C.; Welsh-Bohmer, K.A. Cache County Study Investigators. Occupational exposure to pesticides increases the risk of incident AD the Cache County Study. Neurology 2010, 74, 1524–1530. [Google Scholar] [CrossRef] [PubMed]
- Naughton, S.X.; Terry, A.V., Jr. Neurotoxicity in acute and repeated organophosphate exposure. Toxicology 2018, 408, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Newell, D.R.; Burtles, S.S.; Fox, B.W.; Jodrell, D.I.; Connors, T.A. Evaluation of rodent-only toxicology for early clinical trials with novel cancer therapeutics. Br. J. Cancer 1999, 81, 760. [Google Scholar] [CrossRef] [PubMed]
- Imam, A.; Ajao, M.S.; Ajibola, M.I.; Amin, A.; Abdulmajeed, W.I.; Lawal, A.Z.; Alli-Oluwafuyi, A.; Akinola, O.B.; Oyewopo, A.O.; Olajide, O.J.; et al. Black seed oil reversed scopolamine-induced Alzheimer and corticohippocampal neural alterations in male Wistar rats. Bull. Fac. Pharm. Cairo Univ. 2016, 54, 45–57. [Google Scholar] [CrossRef]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Imam, A.; Ajao, M.S.; Akinola, B.O.; Ajibola, M.I.; Ibrahim, A.; Amin, A.; Abdulmajeed, A.I.; Lawal, Z.A.; Ali-Oluwafuyi, A. Repeated Acute Oral Exposure to Cannabis Sativa Impaired Neurocognitive Behaviours and Cortico-hippocampal Architectonics in Wistar Rats. Niger. J. Physiol. Sci. 2016, 31, 153–159. [Google Scholar]
- Imam, A.; Ajao, M.S.; Amin, A.; Abdulmajeed, W.I.; Ajibola, M.I.; Ibrahim, A.; Olajide, O.J.; Balogun, W.G. Cannabis Induced Moto-Cognitive Dysfunctions in Wistar Rats: Ameliorative efficacy of Nigella sativa. Malays. J. Med. Sci. 2016, 23, 17–28. [Google Scholar] [PubMed]
- Walsh, R.N. Cummins, R.A. The open-field test: A critical review. Psychol. Bull. 1976, 83, 482. [Google Scholar] [CrossRef] [PubMed]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 1985, 14, 149–167. [Google Scholar] [CrossRef]
- Ahadpour, M.; Eskandari, M.R.; Mashayekhi, V. Mitochondrial oxidativestress and dysfunction induced by isoniazid: Study on isolated rat liver and brain mitochondria. Drug Chem. Toxicol. 2016, 39, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Bargi, R.; Asgharzadeh, F.; Beheshti, F.; Hosseini, M.; Sadeghnia, H.R.; Khazaei, M. The effects of thymoquinone on hippocampal cytokine level, brain oxidative stress status and memory deficits induced by lipopolysaccharide in rats. Cytokine 2017, 96, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Jamal, F.; Quazi, S.; Haque, Q.S.; Singh, S. Interrelation of glycemic status and neuropsychiatric disturbances in farmers with organophosphorus pesticide toxicity. Open Biochem. J. 2016, 10, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Quazi, S.H.; Jamal, F.; Rastogi, S.K. Effect of organo-phosphorus on biochemical parameters on agricultural workers. Asian J. Biochem. 2012, 7, 37–45. [Google Scholar] [CrossRef]
- Imam, A.; Adeboye, M.; Abdulmajeed, W.I.; Alli-Oluwafuyi, A.; Amin, A.; Ibrahim, A.; Gwadabe, S.; Popoola, N.G. Dichlorvos Induced AChE Inhibition in Discrete Brain Regions and the Neuro-Cognitive Implications: Ameliorative Effectof Nigella Sativa. Iran. J. Toxic. 2018, 12, 11–16. [Google Scholar]
- Flora, S.J.S.; Dwivedi, N.; Deb, U.; Kushwaha, P.; Lomash, V. Effects of co-exposure to arsenic and dichlorvos on glutathione metabolism, neurological, hepatic variables and tissue histopathology in rats. Toxicol. Res. 2014, 3, 23. [Google Scholar] [CrossRef]
- Wang, H.P.; Liang, Y.J.; Sun, Y.J.; Hou, W.Y.; Chen, J.X.; Long, D.X.; Xu, M.Y.; Wu, Y.J. Subchronic Neurotoxicity of Chlorpyrifos, Carbaryl, and Their Combination in Rats. Environ. Toxicol. 2013, 29, 1193–1200. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Ojo, M.; Afolabi, T.T.; Arowoogun, M.D.; Nwawolor, D.; Farombi, E.O. Protective properties of 6-gingerol-rich fraction from Zingiberofficinale (Ginger) on chlorpyrifos-induced oxidative damage and inflammation in the brain, ovary and uterus of rats. Chem. Biol. Interact. 2017, 25, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Neeraj, S.; Vivek, L.; Jie, L.; Phang, P.; Ahmed, A.; Bharathi, P.; Dharmin, R.; Souvarish, S.; Huajun, J.; Vellareddy, A.; et al. Organophosphate pesticide chlorpyrifos impairs STAT1 signaling to induce dopaminergic neurotoxicity: Implications for mitochondria mediated. Neurobiol. Dis. 2018, 117, 82–113. [Google Scholar] [CrossRef]
- Rasha, M.H.; Wafaa, R.M.; Hany, A.O. A neuroprotective role of kaempferol against chlorpyrifos-induced oxidative stress and memory deficits in rats via GSK3β-Nrf2 signaling pathway. Pestic. Biochem. Physiol. 2018. [Google Scholar] [CrossRef]
- Hariri, A.T.; Moallem, S.A.; Mahmoudi, M.; Hosseinzadeh, H. The effect of crocin and safranal, constituents of saffron, against subacute effect of diazinon on hematologicalandgenotoxicity indices in rats. Phytomedicine 2011, 18, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Varsha, S.; Rupali, P. In vivo antioxidative and neuroprotective effect of 4-Allyl-2-methoxyphenol against chlorpyrifos-induced neurotoxicity in rat brain. Mol. Cell. Biochem. 2014, 388, 61–74. [Google Scholar] [CrossRef]
- Xu, M.-Y.; Wang, P.; Sun, Y.-J.; Wu, Y.-J. Metabolomic analysis for combined hepatotoxicity of chlorpyrifos and cadmium in rats. Toxicology 2017, 384, 50–58. [Google Scholar] [CrossRef]
- Nixon, K.; Crews, F.T. Binge ethanol exposure decreases neurogenesis in adult rat hippocampus. J. Neurochem. 2002, 83, 1087–1093. [Google Scholar] [CrossRef]
- Taffe, M.A.; Kotzebue, R.W.; Crean, R.D.; Crawford, E.F.; Edwards, S.; Mandyam, C.D. Long-lasting reduction in hippocampal neurogenesis by alcohol consumption in adolescent nonhuman primates. Proc. Natl. Acad. Sci. USA 2010, 107, 11104–11109. [Google Scholar] [CrossRef]
- Van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef]
- Parihar, V.K.; Hattiangady, B.; Shuai, B.; Shetty, A.K. Mood and memory deficits in a model of Gulf War Illness are linked with reduced neurogenesis, partial neuron loss, and mild inflammation in the hippocampus. Neuropsychopharmacology 2013, 38, 2348–2362. [Google Scholar] [CrossRef]
- Kohman, R.A.; Rhodes, J.S. Neurogenesis, inflammation and behavior. Brain Behav. Immun. 2013, 27, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Jokanović, M. Neurotoxic effects of organophosphorus pesticides and possible association with neurodegenerative diseases in man: A review. Toxicology 2018, 410, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Aly, N.; Kawther, E.G.; Mahmoud, F.; El-Sebae, A.K. Protective effect of vitamin C against chlorpyrifos oxidative stress in male mice. Pestic. Biochem. Physiol. 2010, 97, 7–12. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, R. Dichlorvos and lindane induced oxidative stress in rat brain: Protective effects of ginger. Pharmacogn. Res. 2012, 4, 27–32. [Google Scholar]
- Nasr, H.M.; El-Demerdash, F.M.; El-Nagar, W.A. Neuro and renal toxicity induced by chlorpyrifos and abamectin in rats: Toxicity of insecticide mixture. Environ. Sci. Pollut. Res. Int. 2016, 23, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- Deveci, H.A.; Karapehlivan, M. Chlorpyrifos-induced parkinsonian model in mice: Behavior, histopathology and biochemistry. Pestic. Biochem. Physiol. 2017, 144, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Taghavian, F.; Vaezi, G.; Abdollahi, M.; Malekirad, A.A. A comparative study of the quality of life, depression, anxiety and stress in farmers exposed to organophosphate pesticides with those in a control group. J. Chem. Health Risks 2016, 6, 143–151. [Google Scholar]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef]
- Nie, Y.; Luo, D.; Yang, M.; Wang, Y.; Xiong, L.; Gao, L.; Liu, Y.; Liu, H. A meta-analysis on the relationship of the PON genes and alzheimer disease. J. Geriatr. Psychiatry Neurol. 2017, 30, 303–310. [Google Scholar] [CrossRef]
- Paul, K.C.; Sinsheimer, J.S.; Cockburn, M.; Bronstein, J.M.; Bordelon, Y.; Ritz, B. Organophosphate pesticides and PON1 L55M in Parkinson’s disease progression. Environ. Int. 2017, 107, 75–81. [Google Scholar] [CrossRef]
- Matthew, N.; Jason, R.R. Time to get personal: A framework for personalized targeting of oxidative stress in neurotoxicity and neurodegenerative disease. Curr. Opin. Toxicol. 2018, 7, 127–132. [Google Scholar]
- Sara, M.; Mohammad, A. The link of organophosphorus pesticides with neurodegenerative and neurodevelopmental diseases based on evidence and mechanisms. Toxicology 2018, 409, 44–45. [Google Scholar]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imam, A.; Sulaiman, N.A.; Oyewole, A.L.; Chengetanai, S.; Williams, V.; Ajibola, M.I.; Folarin, R.O.; Muhammad, A.S.; Shittu, S.-T.T.; Ajao, M.S. Chlorpyrifos- and Dichlorvos-Induced Oxidative and Neurogenic Damage Elicits Neuro-Cognitive Deficits and Increases Anxiety-Like Behavior in Wild-Type Rats. Toxics 2018, 6, 71. https://doi.org/10.3390/toxics6040071
Imam A, Sulaiman NA, Oyewole AL, Chengetanai S, Williams V, Ajibola MI, Folarin RO, Muhammad AS, Shittu S-TT, Ajao MS. Chlorpyrifos- and Dichlorvos-Induced Oxidative and Neurogenic Damage Elicits Neuro-Cognitive Deficits and Increases Anxiety-Like Behavior in Wild-Type Rats. Toxics. 2018; 6(4):71. https://doi.org/10.3390/toxics6040071
Chicago/Turabian StyleImam, Aminu, Nafeesah Abdulkareem Sulaiman, Aboyeji Lukuman Oyewole, Samson Chengetanai, Victoria Williams, Musa Iyiola Ajibola, Royhaan Olamide Folarin, Asma’u Shehu Muhammad, Sheu-Tijani Toyin Shittu, and Moyosore Salihu Ajao. 2018. "Chlorpyrifos- and Dichlorvos-Induced Oxidative and Neurogenic Damage Elicits Neuro-Cognitive Deficits and Increases Anxiety-Like Behavior in Wild-Type Rats" Toxics 6, no. 4: 71. https://doi.org/10.3390/toxics6040071
APA StyleImam, A., Sulaiman, N. A., Oyewole, A. L., Chengetanai, S., Williams, V., Ajibola, M. I., Folarin, R. O., Muhammad, A. S., Shittu, S.-T. T., & Ajao, M. S. (2018). Chlorpyrifos- and Dichlorvos-Induced Oxidative and Neurogenic Damage Elicits Neuro-Cognitive Deficits and Increases Anxiety-Like Behavior in Wild-Type Rats. Toxics, 6(4), 71. https://doi.org/10.3390/toxics6040071