


Physiological Stress Responses Associated with Microplastic Ingestion in the Benthic Flatfish Bothus podas
 , ,
, ,  ,
,  ,
,  and
and 
Abstract

1. Introduction
2. Materials and Methods
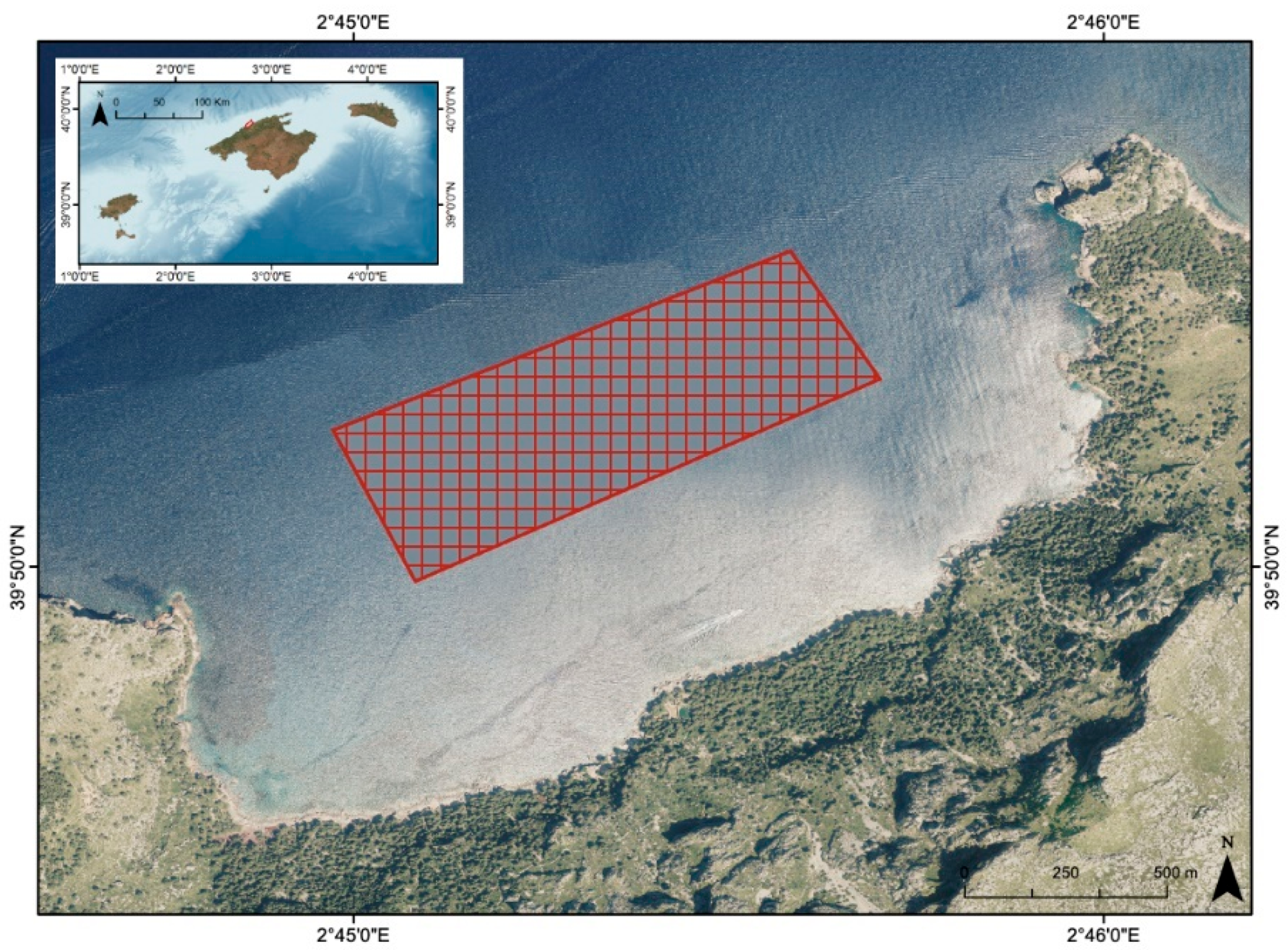
2.1. Area of Study and Fish Sampling
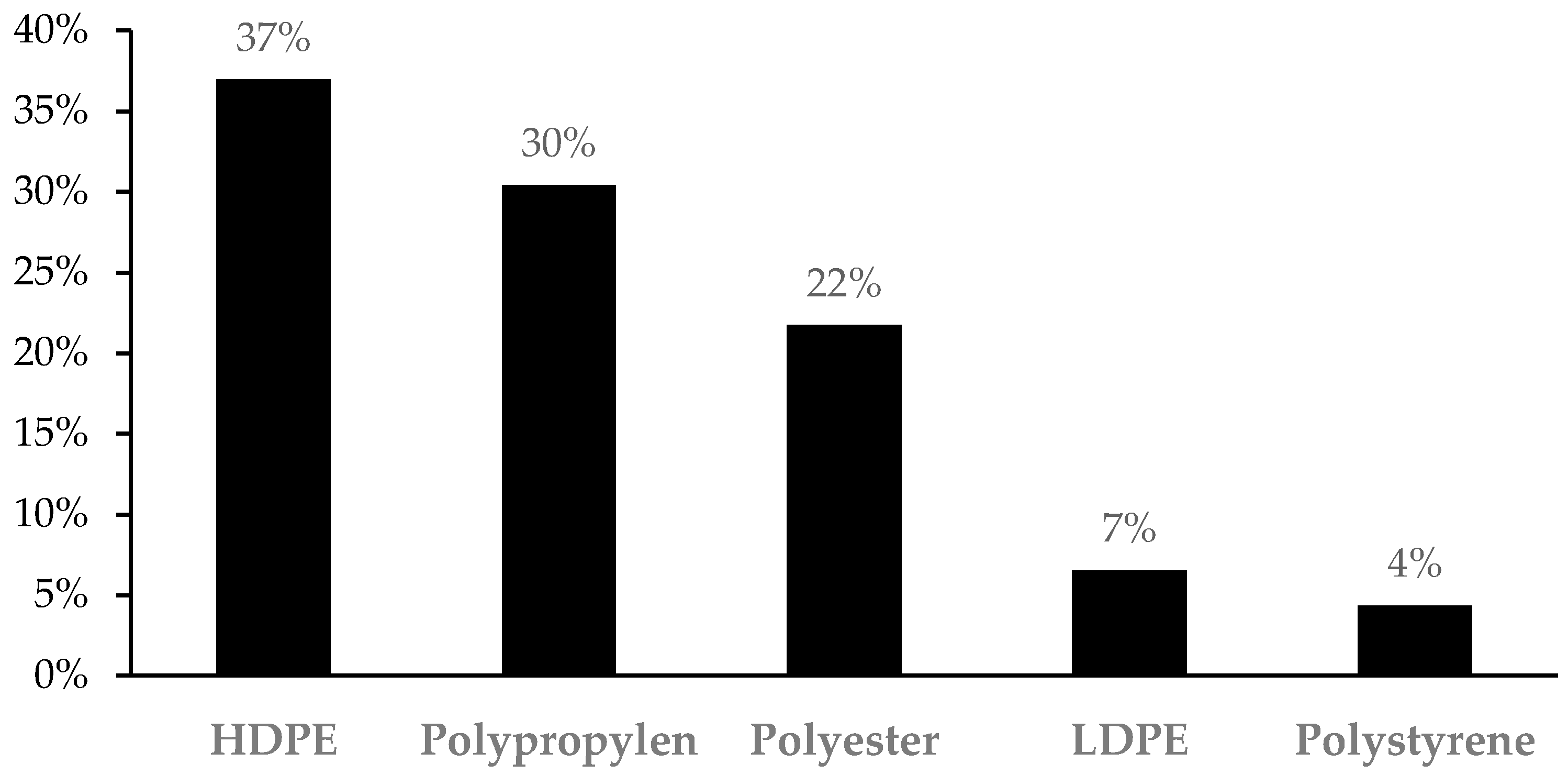
2.2. Microplastic Assessment and Characterization
2.3. Biomarker Analysis
2.4. Statistical Analysis
3. Results
3.1. Biometric Parameters and Microplastic Presence
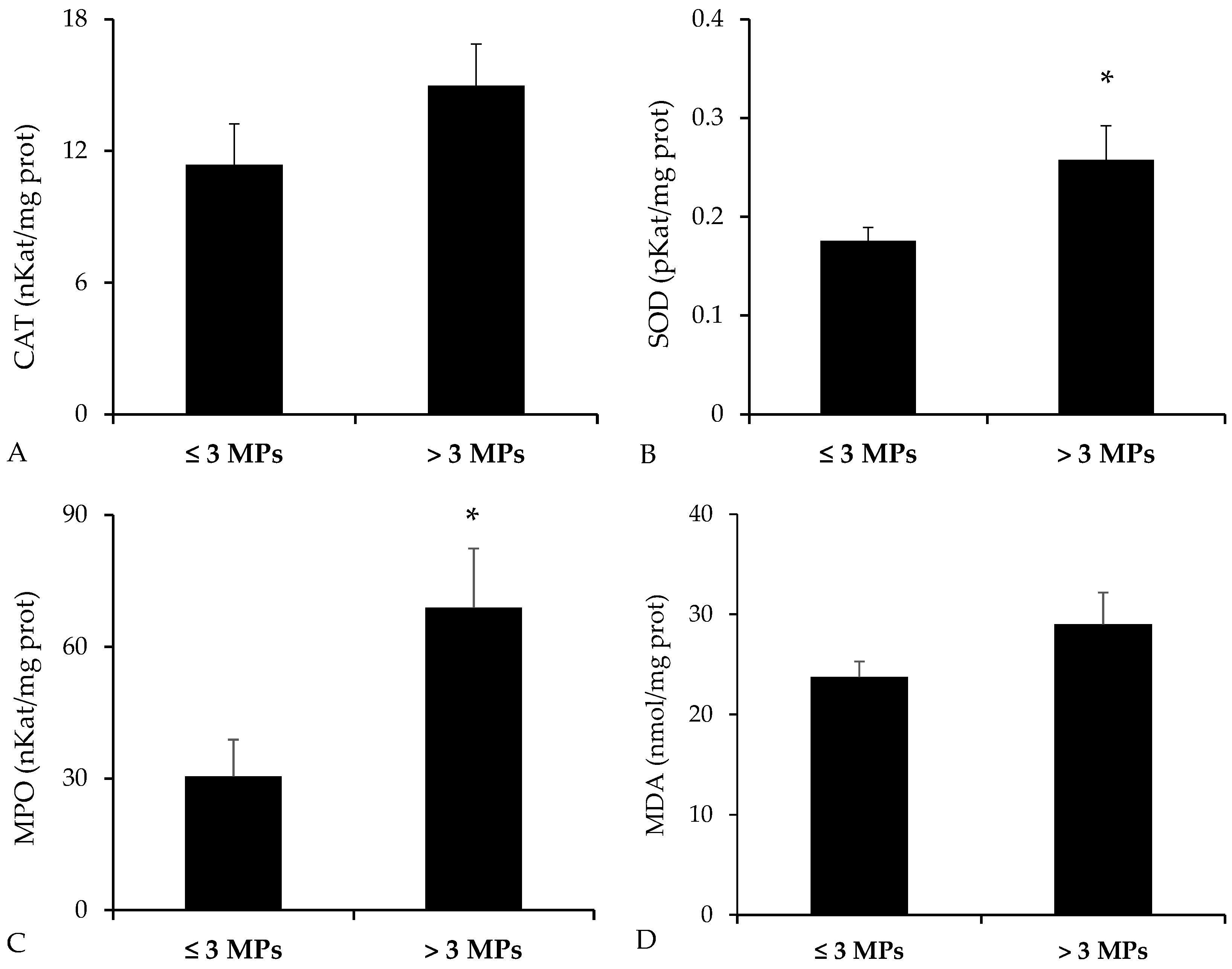
3.2. Biochemical Analysis in the Digestive Tract
3.3. Biomarkers in the Liver
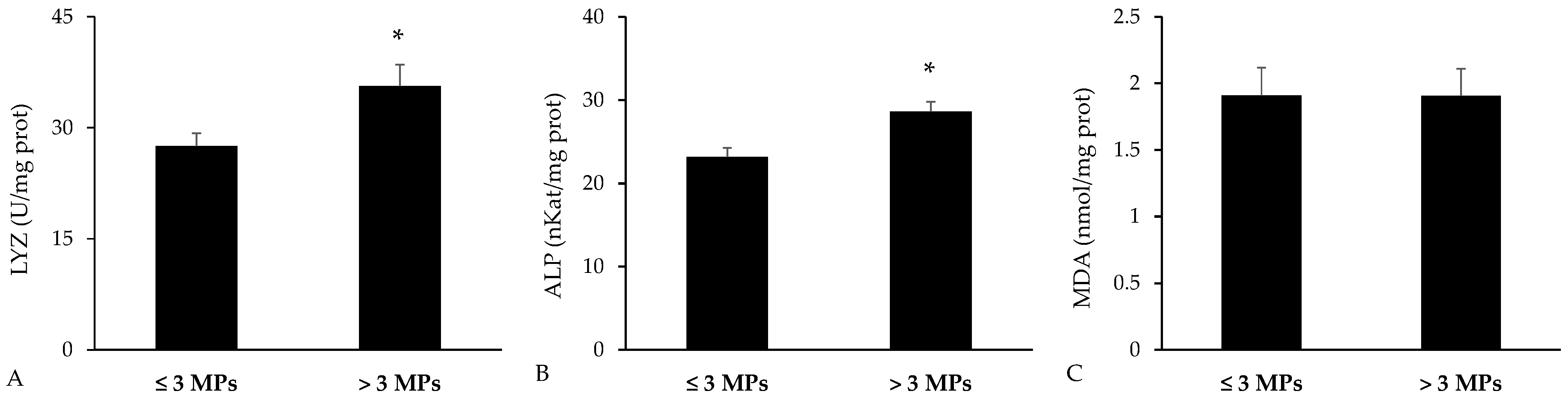
3.4. Biomarkers in the Spleen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| μ-ATR-FTIR | micro-Fourier transform infrared spectroscopy with attenuated total reflection |
| ALP | Alkaline phosphatase |
| CAT | Catalase |
| EROD | Ethoxyresorufin-O-deethylase |
| GST | Glutathione s-transferase |
| HDPE | High-density polyethylene |
| LDPE | Low-density polyethylene |
| MDA | Malondialdehyde |
| MP | Microplastics |
| MPO | Myeloperoxidase |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
References
- Ahmed, M.B.; Rahman, M.S.; Alom, J.; Hasan, M.S.; Johir, M.A.H.; Mondal, M.I.H.; Lee, D.-Y.; Park, J.; Zhou, J.L.; Yoon, M.-H. Microplastic Particles in the Aquatic Environment: A Systematic Review. Sci. Total Environ. 2021, 775, 145793. [Google Scholar] [CrossRef] [PubMed]
- Castro-Castellon, A.T.; Horton, A.A.; Hughes, J.M.R.; Rampley, C.; Jeffers, E.S.; Bussi, G.; Whitehead, P. Ecotoxicity of Microplastics to Freshwater Biota: Considering Exposure and Hazard across Trophic Levels. Sci. Total Environ. 2022, 816, 151638. [Google Scholar] [CrossRef] [PubMed]
- Román-Vega, M.A.; Apún-Molina, J.P.; Escobedo-Urias, D.C.; Muñoz Sevilla, N.P.; Jeyakumar, S.S.L.; Santamaria-Miranda, A. Effects of Plastics and Microplastics on Marine Ecosystems: A Global Review. In Health and Climate Change; Elsevier: Amsterdam, The Netherlands, 2025; pp. 151–173. [Google Scholar]
- Okoye, C.O.; Addey, C.I.; Oderinde, O.; Okoro, J.O.; Uwamungu, J.Y.; Ikechukwu, C.K.; Okeke, E.S.; Ejeromedoghene, O.; Odii, E.C. Toxic Chemicals and Persistent Organic Pollutants Associated with Micro-and Nanoplastics Pollution. Chem. Eng. J. Adv. 2022, 11, 100310. [Google Scholar] [CrossRef]
- Kim, J.-H.; Yu, Y.-B.; Choi, J.-H. Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Neurotoxicity in Fish Exposed to Microplastics: A Review. J. Hazard. Mater. 2021, 413, 125423. [Google Scholar] [CrossRef]
- Alberghini, L.; Truant, A.; Santonicola, S.; Colavita, G.; Giaccone, V. Microplastics in Fish and Fishery Products and Risks for Human Health: A Review. Int. J. Environ. Res. Public Health 2022, 20, 789. [Google Scholar] [CrossRef]
- Xia, X.; Ma, X.; Liang, N.; Qin, L.; Huo, W.; Li, Y. Damage of Polyethylene Microplastics on the Intestine Multilayer Barrier, Blood Cell Immune Function and the Repair Effect of Leuconostoc Mesenteroides DH in the Large-Scale Loach (Paramisgurnus Dabryanus). Fish Shellfish. Immunol. 2024, 147, 109460. [Google Scholar] [CrossRef]
- Ding, R.; Ma, Y.; Li, T.; Sun, M.; Sun, Z.; Duan, J. The Detrimental Effects of Micro-and Nano-Plastics on Digestive System: An Overview of Oxidative Stress-Related Adverse Outcome Pathway. Sci. Total Environ. 2023, 878, 163144. [Google Scholar] [CrossRef]
- Santana, M.S.; de Melo, G.D.; Sandrini-Neto, L.; Di Domenico, M.; Prodocimo, M.M. A meta-analytic review of fish antioxidant defense and biotransformation systems following pesticide exposure. Chemosphere 2022, 291, 132730. [Google Scholar] [CrossRef]
- Grădinariu, L.; Crețu, M.; Vizireanu, C.; Dediu, L. Oxidative Stress Biomarkers in Fish Exposed to Environmental Concentrations of Pharmaceutical Pollutants: A Review. Biology 2025, 14, 472. [Google Scholar] [CrossRef]
- Das, A. The Emerging Role of Microplastics in Systemic Toxicity: Involvement of Reactive Oxygen Species (ROS). Sci. Total Environ. 2023, 895, 165076. [Google Scholar] [CrossRef]
- Li, H.; Liu, H.; Bi, L.; Liu, Y.; Jin, L.; Peng, R. Immunotoxicity of Microplastics in Fish. Fish Shellfish. Immunol. 2024, 150, 109619. [Google Scholar] [CrossRef] [PubMed]
- Cecapolli, E.; Calò, A.; Giakoumi, S.; Di Lorenzo, M.; Greco, S.; Fanelli, E.; Milisenda, G.; Di Franco, A. Sandy Bottoms Have Limited Species Richness but Substantially Contribute to the Regional Coastal Fish β-Diversity: A Case Study of the Central Mediterranean Sea. Mar. Environ. Res. 2024, 201, 106701. [Google Scholar] [CrossRef]
- Govern de les Illes Balears. ES0000225 Sa Costera (ZEC, ZEPA). Available online: https://www.caib.es/sites/xarxanatura/es/es0000225_sa_costera_zec_zepa/ (accessed on 2 June 2025).
- Bahamon, N.; Aguzzi, J.; Ahumada-Sempoal, M.Á.; Bernardello, R.; Reuschel, C.; Company, J.B.; Peters, F.; Gordoa, A.; Navarro, J.; Velásquez, Z.; et al. Stepped Coastal Water Warming Revealed by Multiparametric Monitoring at NW Mediterranean Fixed Stations. Sensors 2020, 20, 2658. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.O.; Gonçalves, A.M.M.; Gonçalves, F.J.M.; Abrantes, N. Improving Cost-Efficiency for MPs Density Separation by Zinc Chloride Reuse. MethodsX 2020, 7, 100785. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in Vitro. Methods Enzym. Anal. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Flohé, L.; Ötting, F. Superoxide Dismutase Assay. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 93–104. [Google Scholar]
- Capeillère-Blandin, C. Oxidation of Guaiacol by Myeloperoxidase: A Two-Electron-Oxidized Guaiacol Transient Species as a Mediator of NADPH Oxidation. Biochem. J. 1998, 336, 395–404. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Ross, N.W.; Firth, K.J.; Wang, A.; Burka, J.F.; Johnson, S.C. Changes in Hydrolytic Enzyme Activities of Naive Atlantic Salmon Salmo Salar Skin Mucus Due to Infection with the Salmon Louse Lepeophtheirus Salmonis and Cortisol Implantation. Dis. Aquat. Organ. 2000, 41, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.D.; Mayer, R.T. Ethoxyresorufin: Direct Fluorimetric Assay of a Microsomal O-Dealkylation Which Is Preferentially Inducible by 3-Methylcholanthrene. Drug Metab. Dispos. 1974, 2, 583–588. [Google Scholar] [CrossRef]
- Lee, Y.C.; Yang, D. Determination of Lysozyme Activities in a Microplate Format. Anal. Biochem. 2002, 310, 223–224. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, V.; Chatterjee, S. Microplastics in the Mediterranean Sea: Sources, Pollution Intensity, Sea Health, and Regulatory Policies. Front. Mar. Sci. 2021, 8, 634934. [Google Scholar] [CrossRef]
- Galgani, F.; Michela, A.; Gérigny, O.; Maes, T.; Tambutté, E.; Harris, P.T. Marine Litter, Plastic, and Microplastics on the Seafloor. In Plastics and the Ocean; Wiley: Hoboken, NJ, USA, 2022; pp. 151–197. [Google Scholar]
- Fagiano, V.; Compa, M.; Alomar, C.; Rios-Fuster, B.; Morató, M.; Capó, X.; Deudero, S. Breaking the Paradigm: Marine Sediments Hold Two-Fold Microplastics than Sea Surface Waters and Are Dominated by Fibers. Sci. Total Environ. 2023, 858, 159722. [Google Scholar] [CrossRef] [PubMed]
- Nayak, R.; Jajpura, L.; Khandual, A. Traditional Fibres for Fashion and Textiles: Associated Problems and Future Sustainable Fibres. Sustain. Fibres Fash. Text. Manuf. 2023, 3–25. [Google Scholar] [CrossRef]
- Maddah, H.A. Polypropylene as a Promising Plastic: A Review. Am. J. Polym. Sci. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Mecozzi, M.; Pietroletti, M.; Monakhova, Y.B. FTIR Spectroscopy Supported by Statistical Techniques for the Structural Characterization of Plastic Debris in the Marine Environment: Application to Monitoring Studies. Mar. Pollut. Bull. 2016, 106, 155–161. [Google Scholar] [CrossRef]
- Pannetier, P.; Vignet, C.; Revel, M.; Le Bihanic, F.; Clérandeau, C.; Mouneyrac, C.; Larcher, T.; Grassl, B.; Gaspéri, J.; Cachot, J.; et al. Juveniles at Risk: Behaviour and Colour Changes in Sole Juveniles (Solea Solea) after Exposure to Estuarine Ragworms (Hediste Diversicolor) Contaminated with Microplastics. Mar. Environ. Res. 2025, 209, 107197. [Google Scholar] [CrossRef]
- Albendín, M.G.; Alarcón, I.; Coello, M.D.; Rodríguez-Barroso, R.; Aranda, V.; Quiroga, J.M.; Arellano, J.M. The Effects of Exposing Solea Senegalensis to Microbeads with and Without Pesticides. Water Air. Soil. Pollut. 2023, 234, 132. [Google Scholar] [CrossRef]
- Esposito, V.; Castriota, L.; Consoli, P.; Romeo, T.; Falautano, M.; Andaloro, F. Feeding Habits and Selectivity of the Wide-Eyed Flounder, Bothus Podas (Delaroche, 1809) (Bothidae) from the Southern Tyrrhenian Sea. Mar. Biol. Res. 2010, 6, 496–502. [Google Scholar] [CrossRef]
- Kılıç, E.; Yücel, N. Microplastic Occurrence in the Gastrointestinal Tract and Gill of Bioindicator Fish Species in the Northeastern Mediterranean. Mar. Pollut. Bull. 2022, 177, 113556. [Google Scholar] [CrossRef]
- Cohen-Sánchez, A.; Solomando, A.; Pinya, S.; Tejada, S.; Valencia, J.M.; Box, A.; Sureda, A. First Detection of Microplastics in Xyrichtys Novacula (Linnaeus 1758) Digestive Tract from Eivissa Island (Western Mediterranean). Environ. Sci. Pollut. Res. 2022, 29, 65077–65087. [Google Scholar] [CrossRef]
- Solomando, A.; Cohen-Sánchez, A.; Box, A.; Montero, I.; Pinya, S.; Sureda, A. Microplastic Presence in the Pelagic Fish, Seriola Dumerili, from Balearic Islands (Western Mediterranean), and Assessment of Oxidative Stress and Detoxification Biomarkers in Liver. Environ. Res. 2022, 212, 113369. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Sánchez, A.; Solomando, A.; Pinya, S.; Tejada, S.; Valencia, J.M.; Box, A.; Sureda, A. Microplastic Presence in the Digestive Tract of Pearly Razorfish Xyrichtys Novacula Causes Oxidative Stress in Liver Tissue. Toxics 2023, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Cocci, P.; Gabrielli, S.; Pastore, G.; Minicucci, M.; Mosconi, G.; Palermo, F.A. Microplastics Accumulation in Gastrointestinal Tracts of Mullus Barbatus and Merluccius Merluccius Is Associated with Increased Cytokine Production and Signaling. Chemosphere 2022, 307, 135813. [Google Scholar] [CrossRef]
- Solomando, A.; Capó, X.; Alomar, C.; Álvarez, E.; Compa, M.; Valencia, J.M.; Pinya, S.; Deudero, S.; Sureda, A. Long-Term Exposure to Microplastics Induces Oxidative Stress and a pro-Inflammatory Response in the Gut of Sparus Aurata Linnaeus, 1758. Environ. Pollut. 2020, 266, 115295. [Google Scholar] [CrossRef]
- Espinosa, C.; Esteban, M.Á.; Cuesta, A. Dietary Administration of PVC and PE Microplastics Produces Histological Damage, Oxidative Stress and Immunoregulation in European Sea Bass (Dicentrarchus Labrax, L.). Fish Shellfish. Immunol. 2019, 95, 574–583. [Google Scholar] [CrossRef]
- Roch, S.; Ros, A.F.H.; Friedrich, C.; Brinker, A. Microplastic Evacuation in Fish Is Particle Size-dependent. Freshw. Biol. 2021, 66, 926–935. [Google Scholar] [CrossRef]
- Torres, S.; Compa, M.; Box, A.; Pinya, S.; Sureda, A. Presence and Potential Effects of Microplastics in the Digestive Tract of Two Small Species of Shark from the Balearic Islands. Fishes 2024, 9, 55. [Google Scholar] [CrossRef]
- Del Piano, F.; Lama, A.; Piccolo, G.; Addeo, N.F.; Iaccarino, D.; Fusco, G.; Riccio, L.; De Biase, D.; Mattace Raso, G.; Meli, R.; et al. Impact of Polystyrene Microplastic Exposure on Gilthead Seabream (Sparus Aurata Linnaeus, 1758): Differential Inflammatory and Immune Response between Anterior and Posterior Intestine. Sci. Total Environ. 2023, 879, 163201. [Google Scholar] [CrossRef]
- Jia, R.; Han, J.; Liu, X.; Li, K.; Lai, W.; Bian, L.; Yan, J.; Xi, Z. Exposure to Polypropylene Microplastics via Oral Ingestion Induces Colonic Apoptosis and Intestinal Barrier Damage through Oxidative Stress and Inflammation in Mice. Toxics 2023, 11, 127. [Google Scholar] [CrossRef]
- Das, B.C.; Ramanan, P.A.; Gorakh, S.S.; Pillai, D.; Vattiringal Jayadradhan, R.K. Sub-Chronic Exposure of Oreochromis Niloticus to Environmentally Relevant Concentrations of Smaller Microplastics: Accumulation and Toxico-Physiological Responses. J. Hazard. Mater. 2023, 458, 131916. [Google Scholar] [CrossRef]
- Liu, X.; Liang, C.; Zhou, M.; Chang, Z.; Li, L. Exposure of Cyprinus Carpio Var. Larvae to PVC Microplastics Reveals Significant Immunological Alterations and Irreversible Histological Organ Damage. Ecotoxicol. Environ. Saf. 2023, 249, 114377. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Chernick, M.; Rittschof, D.; Hinton, D.E. Chronic Dietary Exposure to Polystyrene Microplastics in Maturing Japanese Medaka (Oryzias Latipes). Aquat. Toxicol. 2020, 220, 105396. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarker Type | Name | Tissue/s |
|---|---|---|
| Antioxidants | CAT, SOD | Intestine, liver |
| Oxidative damage | MDA | Intestine, liver, spleen |
| Detoxification | EROD, GST | Intestine, liver |
| Inflammation | MPO | Intestine |
| Immune response | Lysozyme, ALP | Spleen |
| CAT | SOD | MDA | EROD | GST | |
|---|---|---|---|---|---|
| ≥3 MPs | 21.0 ± 2.5 | 0.30 ± 0.03 | 19.4 ± 3.7 | 27.9 ± 2.2 | 106.4 ± 9.7 |
| <3 MPs | 25.7 ± 4.9 | 0.29 ± 0.02 | 19.5 ± 1.9 | 31.6 ± 2.9 | 121.7 ± 10.1 |
| p value | p = 0.630 | p = 0.717 | p = 0.992 | p = 0.324 | p = 0.284 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cohen-Sánchez, A.; Compa, M.; Lombardo, J.; Quetglas-Llabrés, M.M.; Ribas-Taberner, M.d.M.; Jiménez-García, M.; Tejada, S.; Sureda, A. Physiological Stress Responses Associated with Microplastic Ingestion in the Benthic Flatfish Bothus podas. Toxics 2025, 13, 584. https://doi.org/10.3390/toxics13070584
Cohen-Sánchez A, Compa M, Lombardo J, Quetglas-Llabrés MM, Ribas-Taberner MdM, Jiménez-García M, Tejada S, Sureda A. Physiological Stress Responses Associated with Microplastic Ingestion in the Benthic Flatfish Bothus podas. Toxics. 2025; 13(7):584. https://doi.org/10.3390/toxics13070584
Chicago/Turabian StyleCohen-Sánchez, Amanda, Montserrat Compa, Jessica Lombardo, Maria Magdalena Quetglas-Llabrés, Maria del Mar Ribas-Taberner, Manuel Jiménez-García, Silvia Tejada, and Antoni Sureda. 2025. "Physiological Stress Responses Associated with Microplastic Ingestion in the Benthic Flatfish Bothus podas" Toxics 13, no. 7: 584. https://doi.org/10.3390/toxics13070584
APA StyleCohen-Sánchez, A., Compa, M., Lombardo, J., Quetglas-Llabrés, M. M., Ribas-Taberner, M. d. M., Jiménez-García, M., Tejada, S., & Sureda, A. (2025). Physiological Stress Responses Associated with Microplastic Ingestion in the Benthic Flatfish Bothus podas. Toxics, 13(7), 584. https://doi.org/10.3390/toxics13070584