
Metabolic Disruptions in Zebrafish Induced by α-Cypermethrin: A Targeted Metabolomics Study

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Fish
2.3. Acute Toxicity Assessment of α-Cypermethrin in Zebrafish
2.4. Chemical Exposure and Sample Collection
2.5. Water Sample Analysis
2.6. Sample Preparation for Metabolomics Analysis
2.7. Instrumental Conditions for Targeted Metabolites Using GC-MS/MS
2.8. Statistical Analysis
3. Results and Discussion
3.1. Determination of Acute Toxicity of α-Cypermethrin in Zebrafish
3.2. Exposure Concentration Verification
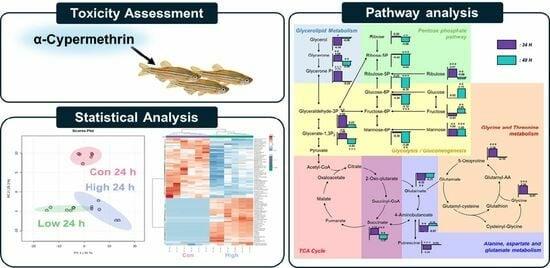
3.3. Metabolomic Alternation Induced by α-Cypermethrin
3.4. Metabolic Pathway Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MSTFA | N-methyl-N-(trimethylsilyl)trifluoroacetamide reagent containing 1% trimethylsilyl chloride |
| UHPLC–MS/MS | Ultra-high performance liquid chromatography tandem mass spectrometry |
| MRM | Multiple reaction monitoring |
| PCA | Principal component analysis |
| PLS-DA | Partial least squares-discriminant analysis |
| GABA | 4-Aminobutanoate |
References
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of pesticides on environment. In Plant, Soil and Microbes: Volume 1: Implications in Crop Science; Springer: Berlin/Heidelberg, Germany, 2016; pp. 253–269. [Google Scholar]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Ewald, J.A.; Wheatley, C.J.; Aebischer, N.J.; Moreby, S.J.; Duffield, S.J.; Crick, H.Q.; Morecroft, M.B. Influences of extreme weather, climate and pesticide use on invertebrates in cereal fields over 42 years. Glob. Change Biol. 2015, 21, 3931–3950. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, Y.; Arshad, M.; Mahmood, N.; Kächele, H.; Kong, R. Occupational hazards, health costs, and pesticide handling practices among vegetable growers in Pakistan. Environ. Res. 2021, 200, 111340. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.; Gawlik, B.M.; Locoro, G.; Rimaviciute, E.; Contini, S.; Bidoglio, G. EU-wide survey of polar organic persistent pollutants in European river waters. Environ. Pollut. 2009, 157, 561–568. [Google Scholar] [CrossRef]
- Brown, C.D.; Van Beinum, W. Pesticide transport via sub-surface drains in Europe. Environ. Pollut. 2009, 157, 3314–3324. [Google Scholar] [CrossRef]
- Mitra, A.; Chatterjee, C.; Mandal, F.B. Synthetic chemical pesticides and their effects on birds. Res. J. Environ. Toxicol. 2011, 5, 81–96. [Google Scholar] [CrossRef]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1. [Google Scholar] [CrossRef]
- van der Werf, H.M. Assessing the impact of pesticides on the environment. Agric. Ecosyst. Environ. 1996, 60, 81–96. [Google Scholar] [CrossRef]
- Sharma, A.; Shukla, A.; Attri, K.; Kumar, M.; Kumar, P.; Suttee, A.; Singh, G.; Barnwal, R.P.; Singla, N. Global trends in pesticides: A looming threat and viable alternatives. Ecotoxicol. Environ. Saf. 2020, 201, 110812. [Google Scholar] [CrossRef]
- Maggi, F.; Tang, F.H.; Black, A.J.; Marks, G.B.; McBratney, A. The pesticide health risk index-An application to the world’s countries. Sci. Total Environ. 2021, 801, 149731. [Google Scholar] [CrossRef]
- Yao, G.; Gao, J.; Zhang, C.; Jiang, W.; Wang, P.; Liu, X.; Liu, D.; Zhou, Z. Enantioselective degradation of the chiral alpha-cypermethrin and detection of its metabolites in five plants. Environ. Sci. Pollut. Res. 2019, 26, 1558–1564. [Google Scholar] [CrossRef]
- Kaur, R.; Singh, J. Toxicity, monitoring, and biodegradation of cypermethrin insecticide: A review. Nat. Environ. Pollut. Technol. 2021, 20, 1997–2005. [Google Scholar] [CrossRef]
- Kansal, I.; Kapoor, A.; Solanki, S.; Singh, R. Cypermethrin toxicity in the environment: Analytical insight into detection methods and microbial degradation pathways. J. Appl. Microbiol. 2023, 134, lxad105. [Google Scholar] [CrossRef]
- Farag, M.R.; Alagawany, M.; Bilal, R.M.; Gewida, A.G.; Dhama, K.; Abdel-Latif, H.M.; Amer, M.S.; Rivero-Perez, N.; Zaragoza-Bastida, A.; Binnaser, Y.S. An overview on the potential hazards of pyrethroid insecticides in fish, with special emphasis on cypermethrin toxicity. Animals 2021, 11, 1880. [Google Scholar] [CrossRef]
- Gonçalves, Í.F.S.; Souza, T.M.; Vieira, L.R.; Marchi, F.C.; Nascimento, A.P.; Farias, D.F. Toxicity testing of pesticides in zebrafish—A systematic review on chemicals and associated toxicological endpoints. Environ. Sci. Pollut. Res. 2020, 27, 10185–10204. [Google Scholar] [CrossRef]
- Jin, Y.; Zheng, S.; Pu, Y.; Shu, L.; Sun, L.; Liu, W.; Fu, Z. Cypermethrin has the potential to induce hepatic oxidative stress, DNA damage and apoptosis in adult zebrafish (Danio rerio). Chemosphere 2011, 82, 398–404. [Google Scholar] [CrossRef]
- Nema, S.; Bhargava, Y. Quantitative assessment of cypermethrin induced behavioural and biochemical anomalies in adult zebrafish. Neurotoxicol. Teratol. 2018, 68, 57–65. [Google Scholar] [CrossRef]
- Mu, X.; Shen, G.; Huang, Y.; Luo, J.; Zhu, L.; Qi, S.; Li, Y.; Wang, C.; Li, X. The enantioselective toxicity and oxidative stress of beta-cypermethrin on zebrafish. Environ. Pollut. 2017, 229, 312–320. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.; Ren, L.; Feng, W.; Lv, P.; Wu, W.; Yan, Y. The single and joint toxicity effects of chlorpyrifos and beta-cypermethrin in zebrafish (Danio rerio) early life stages. J. Hazard. Mater. 2017, 334, 121–131. [Google Scholar] [CrossRef]
- Bradberry, S.M.; Cage, S.A.; Proudfoot, A.T.; Vale, J.A. Poisoning due to pyrethroids. Toxicol. Rev. 2005, 24, 93–106. [Google Scholar] [CrossRef]
- Robertson, D.G.; Watkins, P.B.; Reily, M.D. Metabolomics in toxicology: Preclinical and clinical applications. Toxicol. Sci. 2011, 120, S146–S170. [Google Scholar] [CrossRef]
- Roberts, L.D.; Souza, A.L.; Gerszten, R.E.; Clish, C.B. Targeted metabolomics. Curr. Protoc. Mol. Biol. 2012, 98, 30.32.1–30.32.24. [Google Scholar] [CrossRef]
- Zhang, W.; Li, F.; Nie, L. Integrating multiple ‘omics’ analysis for microbial biology: Application and methodologies. Microbiology 2010, 156, 287–301. [Google Scholar] [CrossRef]
- Chen, C.; Wang, J.; Pan, D.; Wang, X.; Xu, Y.; Yan, J.; Wang, L.; Yang, X.; Yang, M.; Liu, G.P. Applications of multi-omics analysis in human diseases. MedComm 2023, 4, e315. [Google Scholar] [CrossRef]
- Bouhifd, M.; Hartung, T.; Hogberg, H.T.; Kleensang, A.; Zhao, L. toxicometabolomics. J. Appl. Toxicol. 2013, 33, 1365–1383. [Google Scholar] [CrossRef]
- Bernhard, A.; Poulsen, R.; Brun Hansen, A.M.; Hansen, M. Toxicometabolomics as a tool for next generation environmental risk assessment. EFSA J. 2023, 21, e211005. [Google Scholar] [CrossRef]
- Araújo, A.M.; Carvalho, F.; Guedes de Pinho, P.; Carvalho, M. Toxicometabolomics: Small molecules to answer big toxicological questions. Metabolites 2021, 11, 692. [Google Scholar] [CrossRef]
- Radilov, A.S.; Ukolov, A.I. Toxicometabolomics—Integration of preventive and analytical toxicology. Toxicol. Rev. 2022, 30, 286–296. [Google Scholar] [CrossRef]
- Lai, K.P.; Gong, Z.; Tse, W.K.F. Zebrafish as the toxicant screening model: Transgenic and omics approaches. Aquat. Toxicol. 2021, 234, 105813. [Google Scholar] [CrossRef]
- Verma, S.K.; Nandi, A.; Sinha, A.; Patel, P.; Mohanty, S.; Jha, E.; Jena, S.; Kumari, P.; Ghosh, A.; Jerman, I. The posterity of Zebrafish in paradigm of in vivo molecular toxicological profiling. Biomed. Pharmacother. 2024, 171, 116160. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, Y.; Hu, N.; Long, D.; Cao, Y. The uses of zebrafish (Danio rerio) as an in vivo model for toxicological studies: A review based on bibliometrics. Ecotoxicol. Environ. Saf. 2024, 272, 116023. [Google Scholar] [CrossRef]
- OECD, T.N. 203: Fish, Acute Toxicity Test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 1992; Volume 2. [Google Scholar]
- Chin, W.W. The partial least squares approach to structural equation modeling. Mod. Methods Bus. Res. 1998, 295, 295–336. [Google Scholar]
- McCombie, G.; Browning, L.M.; Titman, C.M.; Song, M.; Shockcor, J.; Jebb, S.A.; Griffin, J.L. ω-3 oil intake during weight loss in obese women results in remodelling of plasma triglyceride and fatty acids. Metabolomics 2009, 5, 363–374. [Google Scholar] [CrossRef]
- Henseler, J.; Ringle, C.M.; Sinkovics, R.R. The use of partial least squares path modeling in international marketing. In New Challenges to International Marketing; Emerald Group Publishing Limited: Leeds, UK, 2009; Volume 20, pp. 277–319. [Google Scholar]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of energy metabolism by long-chain fatty acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Wu, G.; Lupton, J.R.; Turner, N.D.; Fang, Y.-Z.; Yang, S. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef]
- Hellou, J.; Ross, N.W.; Moon, T.W. Glutathione, glutathione S-transferase, and glutathione conjugates, complementary markers of oxidative stress in aquatic biota. Environ. Sci. Pollut. Res. 2012, 19, 2007–2023. [Google Scholar] [CrossRef]
- Seo, J.H.; Pyo, S.; Shin, Y.-K.; Nam, B.-G.; Kang, J.W.; Kim, K.P.; Lee, H.Y.; Cho, S.-R. The effect of environmental enrichment on glutathione-mediated xenobiotic metabolism and antioxidation in normal adult mice. Front. Neurol. 2018, 9, 425. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J.L. Zebrafish get connected: Investigating neurotransmission targets and alterations in chemical toxicity. Toxics 2016, 4, 19. [Google Scholar] [CrossRef]
- Molina-Rueda, J.; Garrido-Aranda, A.; Gallardo, F. Glutamate decarboxylase. In Amino Acids in Higher Plants; CAB International: Wallingford, UK, 2015; pp. 129–141. [Google Scholar]
- Verma, R.; Choudhary, P.R.; Nirmal, N.K.; Syed, F.; Verma, R. Neurotransmitter systems in zebrafish model as a target for neurobehavioural studies. Mater. Today Proc. 2022, 69, 1565–1580. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Time | 24 h | 48 h | 72 h | 96 h |
|---|---|---|---|---|
| LC50 (µg/L) | 1.54 | 1.54 | 1.28 | 1.28 |
| 95% Confidence limits (µg/L) | 1.16–2.05 | 1.16–2.05 | 1.07–1.54 | 1.07–1.54 |
| Exposure Time | 0.15 µg/L | 1.5 µg/L | ||
|---|---|---|---|---|
| Conc.(µg/L) | Ratio (%) * | Conc. (µg/L) | Ratio (%) * | |
| 0 h | 0.147 | 98 | 1.47 | 98 |
| 24 h | 0.140 | 93 | 1.44 | 96 |
| 48 h | 0.138 | 92 | 1.42 | 95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ok, H.-J.; Yu, J.-W.; Lee, J.-H.; Choi, E.-S.; Kim, J.-H.; Jeon, Y.; Noh, W.; Choi, S.-G.; Kim, J.-H.; Song, M.-H.; et al. Metabolic Disruptions in Zebrafish Induced by α-Cypermethrin: A Targeted Metabolomics Study. Toxics 2025, 13, 529. https://doi.org/10.3390/toxics13070529
Ok H-J, Yu J-W, Lee J-H, Choi E-S, Kim J-H, Jeon Y, Noh W, Choi S-G, Kim J-H, Song M-H, et al. Metabolic Disruptions in Zebrafish Induced by α-Cypermethrin: A Targeted Metabolomics Study. Toxics. 2025; 13(7):529. https://doi.org/10.3390/toxics13070529
Chicago/Turabian StyleOk, Hang-Ji, Ji-Woo Yu, Jung-Hoon Lee, Eun-Song Choi, Jong-Hwan Kim, Yoonjeong Jeon, Won Noh, Sung-Gil Choi, Jeong-Han Kim, Min-Ho Song, and et al. 2025. "Metabolic Disruptions in Zebrafish Induced by α-Cypermethrin: A Targeted Metabolomics Study" Toxics 13, no. 7: 529. https://doi.org/10.3390/toxics13070529
APA StyleOk, H.-J., Yu, J.-W., Lee, J.-H., Choi, E.-S., Kim, J.-H., Jeon, Y., Noh, W., Choi, S.-G., Kim, J.-H., Song, M.-H., & Lee, J.-H. (2025). Metabolic Disruptions in Zebrafish Induced by α-Cypermethrin: A Targeted Metabolomics Study. Toxics, 13(7), 529. https://doi.org/10.3390/toxics13070529