

Using Blood Gas Analysis and Capnography to Determine Oxygenation Status in Bottlenose Dolphins (Tursiops truncatus) Following the Deepwater Horizon Oil Spill

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Field-Based Dolphin Sample Collection
2.2. Managed Dolphin Sample Collection
2.3. Data and Statistical Analysis
3. Results
3.1. Pulmonary Ultrasound
3.2. Physical Exam
3.3. Pulse Oximetry
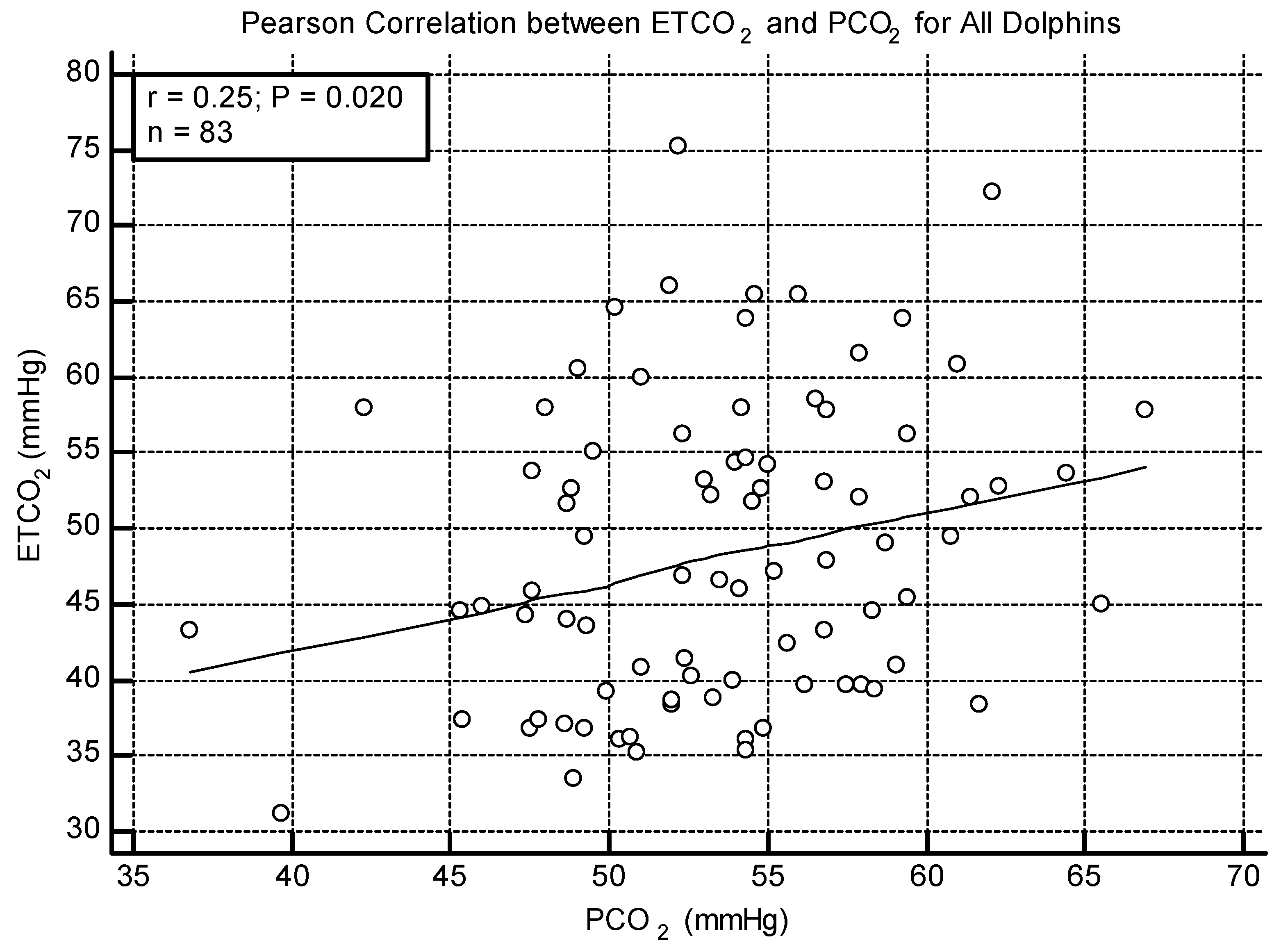
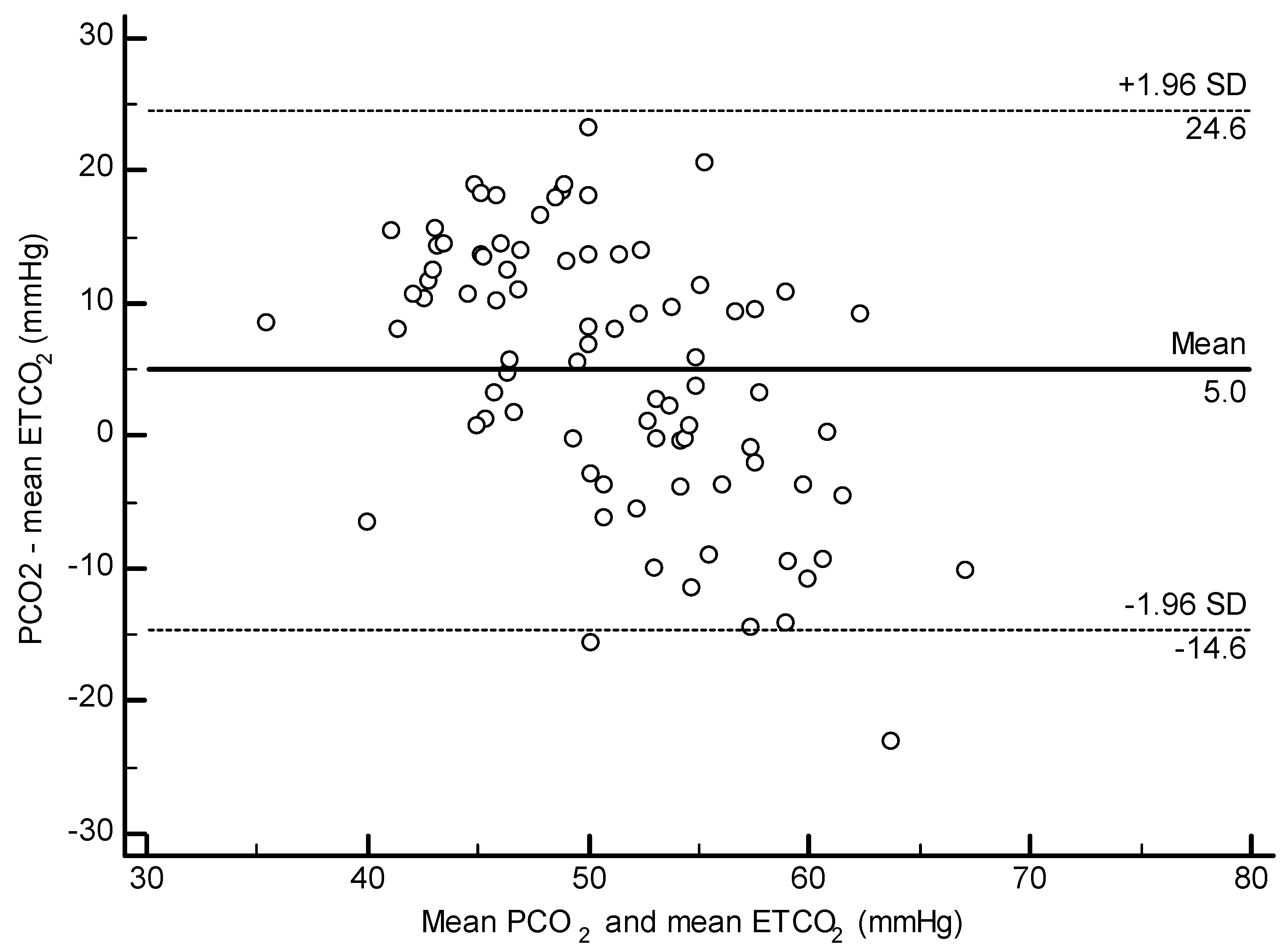
3.4. ETCO2 and PCO2
3.5. All Dolphin Analysis
3.6. Navy Cohort
3.7. Barataria Bay Cohort
3.8. Time Differential Analysis
3.9. Prognostic Score Analysis
4. Discussion
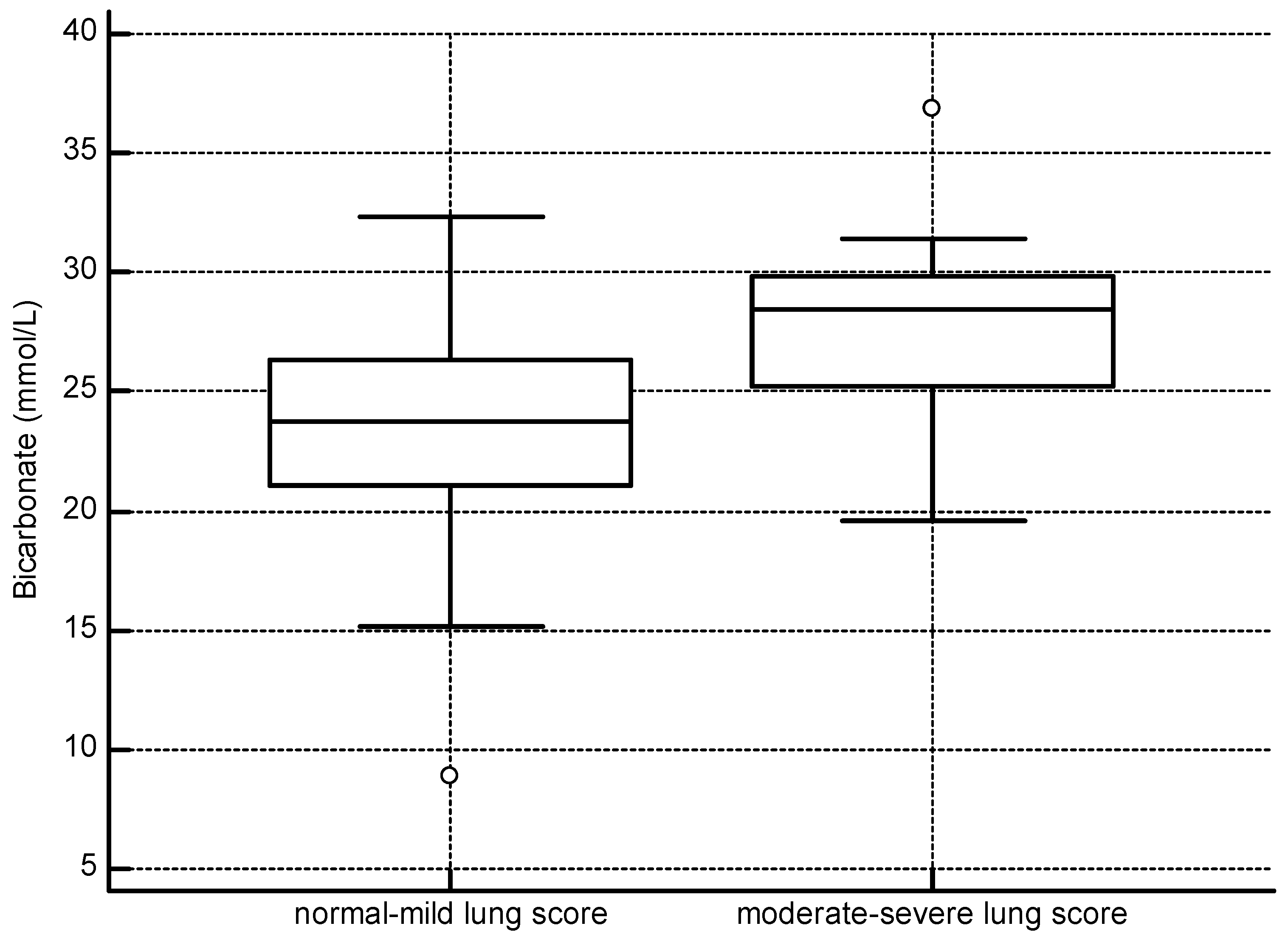
4.1. Pulmonary Disease
4.2. Barataria Bay vs. Navy Dolphin Cohorts
4.3. Sex and Pregnancy
4.4. Capnography
4.5. Blood Gas Interpretation in Dolphins
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United States of America; BP Exploration & Production, Inc. Findings of fact and conclusions of law: Phase two trial. In Re: Oil Spill by the Oil Rig Deepwater Horizons in the Gulf of Mexico, on 20 April 2010; No. MDL 2179, 2015 WL 225421. L.A. E.D. Jan. 15, 2015. Doc 14021; US District Court for the Eastern District of Louisiana: New Orleans, LA, USA, 2015. Available online: http://www.laed.uscourts.gov/sites/default/files/OilSpill/Orders/1152015FindingsPhaseTwo.pdf (accessed on 5 September 2019).
- Schwacke, L.H.; Smith, C.R.; Townsend, F.I.; Wells, R.S.; Hart, L.B.; Balmer, B.C.; Collier, T.K.; De Guise, S.; Fry, M.M.; Guillette, J.L.J.; et al. Health of common bottlenose dolphins (Tursiops truncatus) in Barataria Bay, Louisiana, following the Deepwater Horizon oil spill. Environ. Sci. Technol. 2014, 48, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Lane, S.M.; Smith, C.R.; Mitchell, J.; Balmer, B.C.; Barry, K.P.; McDonald, T.; Mori, C.S.; Rosel, P.E.; Rowles, T.K.; Speakman, T.R.; et al. Reproductive outcome and survival of common bottlenose dolphins sampled in Barataria Bay, Louisiana, USA, following the Deepwater Horizon oil spill. Proc. Biol. Sci. 2015, 282, 20151944. [Google Scholar] [CrossRef] [PubMed]
- Venn-Watson, S.; Colegrove, K.M.; Litz, J.; Kinsel, M.; Terio, K.; Saliki, J.; Fire, S.; Carmichael, R.; Chevis, C.; Hatchett, W.; et al. gland and lung lesions in Gulf of Mexico common bottlenose dolphins (Tursiops truncatus) found dead following the Deepwater Horizon oil spill. PLoS ONE 2015, 10, e0126538. [Google Scholar] [CrossRef] [PubMed]
- Kellar, N.M.; Speakman, T.R.; Smith, C.R.; Lane, S.M.; Balmer, B.C.; Trego, M.L.; Catelani, K.N.; Robbins, M.N.; Allen, C.D.; Wells, R.S.; et al. Low reproductive success rates of common bottlenose dolphins Tursiops truncatus in the northern Gulf of Mexico following the Deepwater Horizon disaster (2010–2015). Endanger. Species Res. 2017, 33, 143–158. [Google Scholar] [CrossRef]
- Smith, C.R.; Rowles, T.K.; Hart, L.B.; Townsend, F.I.; Wells, R.S.; Zolman, E.S.; Balmer, B.C.; Quigley, B.; Ivancic, M.; McKercher, W.; et al. Slow recovery of Barataria Bay dolphin health following the Deepwater Horizon oil spill (2013–2014), with evidence of persistent lung disease and impaired stress response. Endanger. Species Res. 2017, 33, 127–142. [Google Scholar] [CrossRef]
- Colegrove, K.; Venn-Watson, S.; Litz, J.; Kinsel, M.; Terio, K.; Fougeres, E.; Ewing, R.; Pabst, D.; McLellan, W.; Raverty, S.; et al. Fetal distress and in utero pneumonia in perinatal dolphins during the Northern Gulf of Mexico unusual mortality event. Dis. Aquat. Org. 2016, 119, 1–16. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Roy-Engel, A.M.; Baddoo, M.C.; Flemington, E.K.; Wang, G.; Wang, H. The impact of oil spill to lung health—Insights from an RNA-seq study of human airway epithelial cells. Gene 2016, 578, 38–51. [Google Scholar] [CrossRef]
- Nie, H.; Liu, H.; Shi, Y.; Lai, W.; Liu, X.; Xi, Z.; Lin, B. Combined multi-omics analysis reveals oil mist particulate matter-induced lung injury in rats: Pathological damage, proteomics, metabolic disturbances, and lung dysbiosis. Ecotox. Environ. Safe. 2022, 241, 113759. [Google Scholar] [CrossRef]
- Tuder, R.M.; Yun, J.H.; Bhunia, A.; Fijalkowska, I. Hypoxia and chronic lung disease. J. Mol. Med. 2007, 85, 1317–1324. [Google Scholar] [CrossRef]
- Hutter, D.; Kingdom, J.; Jaeggi, E. Causes and Mechanisms of Intrauterine Hypoxia and its Impact on the Fetal Cardiovascular System: A Review. Int. J. Pediatr. 2010, 2010, 401323. [Google Scholar] [CrossRef]
- Getahun, D.; Ananth, C.V.; Oyelese, Y.; Peltier, M.R.; Smulian, J.C.; Vintzileos, A.M. Acute and chronic respiratory disease in pregnancy: Associations with spontaneous premature rupture of membranes. J. Matern. Fetal Neonatal Med. 2007, 20, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Ilkiw, J.E.; Rose, R.J.; Martin, I.C.A. A Comparison of Simultaneously Collected Arterial, Mixed Venous, Jugular Venous and Cephalic Venous Blood Samples in the Assessment of Blood-Gas and Acid-Base Status in the Dog. J. Vet. Intern. Med. 1991, 5, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.D. The physiological basis of pulmonary gas exchange: Implications for clinical interpretation of arterial blood gases. Eur. Respir. J. 2015, 45, 227–243. [Google Scholar] [CrossRef]
- Trulock, E.P. Arterial Blood Gases. In Clinical Methods: The History, Physical and Laboratory Examinations, 3rd ed.; Walker, K.W., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990. [Google Scholar]
- Shaffer, S.A.; Costa, D.P.; Williams, T.M.; Ridgway, S.H. Diving and swimming performance of white whales, Delphinapterus leucas: An assessment of plasma lactate and blood gas levels and respiratory rates. J. Exp. Biol. 1997, 200, 3091–3099. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.M.; Haun, J.E.; Friedl, W.A. The diving physiology of bottlenose dolphins (Tursiops truncatus). I. Balancing the demands of exercise for energy conservation at depth. J. Exp. Biol. 1999, 202, 2739–2748. [Google Scholar] [CrossRef]
- Haulena, M.; Heath, R.B. Marine Mammal Anesthesia. In CRC Handbook of Marine Mammal Medicine, 2nd ed.; Dierauf, L.A., Gulland, F.M.D., Eds.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Ridgway, S.H.; McCormick, J.G. Anesthetization of Porpoises for Major Surgery. Science 1967, 158, 510–512. [Google Scholar] [CrossRef]
- Rieu, M.; Gautheron, B. Preliminary observations concerning a method for introduction of a tube for anesthesia in small delphinids. Life Sci. 1968, 7, 141–146. [Google Scholar] [CrossRef]
- ACVAA (American College of Veterinary Anesthesia and Analgesia). Recommendations for Monitoring Anesthetized Veterinary Patients. 2009. Available online: http://www.acvaa.org/docs/Small_Animal_Monitoring_2009.doc (accessed on 26 January 2017).
- Dold, C.; Ridgway, S. Cetaceans. In Zoo Animal and Wildlife Immobilization and Anesthesia; West, G.W., Heard, D., Caukett, N., Eds.; Blackwell Publishing: Ames, IA, USA, 2007. [Google Scholar]
- Kelmer, E.; Scanson, L.C.; Reed, A.; Love, L.C. Agreement between values for arterial and end-tidal partial pressure of carbon dioxide in spontaneously breathing, critically ill dogs. J. Am. Vet. Med. Assoc. 2009, 235, 1314–1318. [Google Scholar] [CrossRef]
- Mortola, J.P.; Seguin, J. End-tidal CO2 in some aquatic mammals of large size. Zoology 2009, 112, 77–85. [Google Scholar] [CrossRef]
- van Elk, C.E.; Epping, N.; Gans, S.L.M. Pulmonary function measurements in dolphins using capnography. Vet. Rec. 2001, 149, 308–309. [Google Scholar] [CrossRef]
- Wells, R.S.; Rhinehart, H.L.; Hansen, L.J.; Sweeney, J.C.; Townsend, F.I.; Stone, R.; Casper, D.; Scott, M.D.; Hohn, A.A.; Rowles, T.K. Bottlenose dolphins as marine ecosystem sentinels: Developing a health monitoring system. EcoHealth 2004, 1, 246–254. [Google Scholar] [CrossRef]
- Barratclough, A.; Wells, R.S.; Schwacke, L.H.; Rowles, T.K.; Gomez, F.M.; Fauquier, D.A.; Sweeney, J.C.; Townsend, F.I.; Hansen, L.J.; Zolman, E.S.; et al. Health Assessments of Common Bottlenose Dolphins (Tursiops truncatus): Past, Present, and Potential Conservation Applications. Front. Vet. Sci. 2019, 6, 444. [Google Scholar] [CrossRef] [PubMed]
- McFee, W.E.; Schwacke, J.H.; Stolen, M.K.; Mullin, K.D.; Schwacke, L.H. Investigation of growth phases for bottlenose dolphins using a Bayesian modeling approach. Mar. Mammal Sci. 2010, 26, 67–85. [Google Scholar] [CrossRef]
- Smith, C.R.; Solano, M.; Lutmerding, B.A.; Johnson, S.P.; Meegan, J.M.; Le-Bert, C.R.; Emory-Gomez, F.; Cassle, S.; Carlin, K.; Jensen, E.D. Pulmonary ultrasound findings in a bottlenose dolphin Tursiops truncatus population. Dis. Aquat. Org. 2012, 101, 243–255. [Google Scholar] [CrossRef]
- Wells, R.; Smith, C.R.; Sweeney, J.; Townsend, F.I.; Fauquier, D.; Stone, R.; Langan, J.; Schwacke, L.; Rowles, T.K. Fetal survival for bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Aquat. Mamm. 2014, 4, 252–259. [Google Scholar] [CrossRef]
- Everitt, B.S.; Dunn, G. Applied Multivariate Data Analysis; Edward Arnold: London, UK, 1991; pp. 219–220. [Google Scholar]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Abdi, H. Holm’s Sequential Bonferroni Procedure. In Encyclopedia of Research Design; Salkind, N., Ed.; Sage: Thousand Oaks, CA, USA, 2010. [Google Scholar]
- Bland, J.M.; Altman, D.G. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet 1986, 1, 307–310. [Google Scholar] [CrossRef]
- Di Guardo, G.; Agrimi, U.; Morelli, L.; Cardeti, G.; Terracciano, G.; Kennedy, S. Post mortem investigations on cetaceans found stranded on the coasts of Italy between 1990 and 1993. Vet. Rec. 1995, 136, 439–442. [Google Scholar] [CrossRef]
- Cornaglia, E.; Rebora, L.; Gilli, C.; Di Guardo, G. Histopathological and immunohistochemical studies on cetaceans found stranded on the coast of Italy between 1990 and 1997. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2000, 47, 129–142. [Google Scholar] [CrossRef]
- Bogomolni, A.L.; Pugliares, K.R.; Sharp, S.M.; Patchett, K.; Harry, C.T.; LaRocque, J.M.; Touhey, K.M.; Moore, M. Mortality trends of stranded marine mammals on Cape Cod and southeastern Massachusetts, USA, 2000 to 2006. Dis. Aquat. Organ. 2010, 88, 143–155. [Google Scholar] [CrossRef]
- Venn-Watson, S.; Jensen, E.D.; Ridgway, S.H. Effects of age and sex on clinicopathologic reference ranges in a healthy managed Atlantic bottlenose dolphin population. J. Am. Vet. Med Assoc. 2007, 231, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Coles, E.H. Veterinary Clinical Pathology; W.B. Saunders: Philadelphia, PA, USA, 1986; pp. 1–8, 203–213. [Google Scholar]
- Stringer, W.; Casaburi, R.; Wasserman, K. Acid-base regulation during exercise and recovery in humans. J. Appl. Physiol. 1992, 72, 854–961. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, D.E.; D’Arsigny, C.; Fitzpatrick, M.; Webb, K.A. Exercise hypercapnia in advanced chronic obstructive pulmonary disease: The role of lung hyperinflation. Am. J. Respir. Crit. Care Med. 2002, 1566, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Schwacke, L.H.; Marques, T.A.; Thomas, L.; Booth, C.G.; Balmer, B.C.; Barratclough, A.; Colegrove, K.; De Guise, S.; Garrison, L.P.; Gomez, F.M.; et al. Modeling population effects of the Deepwater Horizon oil spill on a long-lived species. Conserv. Biol. 2022, 36, e13878. [Google Scholar] [CrossRef]
- Bennett, D.; Mazzei, M.A.; Squitieri, N.C.; Bargagli, E.; Refini, R.M.; Fossi, A.; Volterrani, L.; Rottoli, P. Familial pulmonary fibrosis: Clinical and radiologic characteristics and progression analysis in different high resolution-CT patterns. Respir. Med. 2017, 126, 75–83. [Google Scholar] [CrossRef]
- Lapinsky, S.E.; Tram, C.; Mehta, S.; Maxwell, C.V. Restrictive lung disease in pregnancy. Chest 2014, 145, 394–398. [Google Scholar] [CrossRef]
- Schmidt, S.L.; Tayob, N.; Han, M.K.; Zappala, C.; Kervitsky, D.; Murray, S.; Wells, A.U.; Brown, K.K.; Martinez, F.J.; Flaherty, K.R. Predicting Pulmonary Fibrosis Disease Course from Past Trends in Pulmonary Function. Chest 2014, 145, 579–585. [Google Scholar] [CrossRef]
- Seeger, W.; Adir, Y.; Barberà, J.A.; Champion, H.; Coghlan, J.G.; Cottin, V.; De Marco, T.; Galiè, N.; Ghio, S.; Gibbs, S.; et al. Pulmonary hypertension in chronic lung diseases. J. Am. Coll. Cardiol. 2013, 62 (Suppl. 25), D109–D116. [Google Scholar] [CrossRef]
- Koenig, S.; Chandra, S.; Alaverdian, A.; Dibello, C.; Mayo, P.H.; Narasimhan, M. Ultrasound Assessment of Pulmonary Embolism in Patients Receiving CT Pulmonary Angiography. Chest 2014, 145, 818–823. [Google Scholar] [CrossRef]
- Bass, C.M.; Sajed, D.R.; Adedipe, A.A.; West, T.E. Pulmonary ultrasound and pulse oximetry versus chest radiography and arterial blood gas analysis for the diagnosis of acute respiratory distress syndrome: A pilot study. Crit. Care 2015, 19, 282. [Google Scholar] [CrossRef]
- Irwin, Z.; Cook, J.O. Advances in Point-of-Care Thoracic Ultrasound. Emerg. Med. Clin. N. Am. 2016, 34, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Terasawa, F.; Ohizumi, H.; Ohshita, I. Effect of Breath-Hold on Blood Gas Analysis in Captive Pacific White-Sided Dolphins (Lagenorhynchus obliquidens). J. Vet. Med. Sci. 2010, 72, 1221–1224. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, F.; Kametas, N.; Costello, J.; Rafferty, G.F.; Greenough, A.; Nicolaides, K. Respiratory function in singleton and twin pregnancy. BJOG 2002, 109, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, S.H.; Scronce, B.L.; Kanwisher, J. Respiration and Deep Diving in the Bottlenose Porpoise. Science 1969, 166, 1651–1654. [Google Scholar] [CrossRef] [PubMed]
- Fahlman, A.; Loring, S.H.; Levine, G.; Rocho-Levine, J.; Austin, T.; Brodsky, M. Lung mechanics and pulmonary function testing in cetaceans. J. Exp. Biol. 2015, 218, 2030–2038. [Google Scholar] [CrossRef] [PubMed]
- Askenov, A.A.; Yeates, L.; Pasamontes, A.; Siebe, C.; Zrodnikov, Y.; Simmons, J.; McCartney, M.M.; Deplanque, J.P.; Wells, R.S.; Davies, C.E. Metabolite Content Profiling of Bottlenose Dolphin Exhaled Breath. Anal. Chem. 2014, 86, 10616–10624. [Google Scholar]
- Yeates, L.C.; Carlin, K.P.; Baird, M.; Venn-Watson, S.; Ridgway, S. Nitric oxide in the breath of bottlenose dolphins: Effects of breath hold duration, feeding and lung disease. Mar. Mammal Sci. 2014, 30, 272–281. [Google Scholar] [CrossRef]
- Kiefmann, M.; Tank, S.; Tritt, M.O.; Keller, P.; Heckel, K.; Schulte-Uentrop, L.; Olotu, C.; Schrepfer, S.; Goetz, A.E.; Kiefmann, R. Dead space ventilation promotes alveolar hypocapnia reducing surfactant secretion by altering mitochondrial function. Thorax 2019, 74, 219–228. [Google Scholar] [CrossRef]
- Varela, R.A.; Schwacke, L.; Fair, P.A.; Bossart, G.D. Effects of duration of capture and sample handling on critical care blood analytes in free-ranging bottlenose dolphins. J. Am. Vet. Med. Assoc. 2006, 226, 1955–1961. [Google Scholar] [CrossRef]
- Sharp, S.M.; Knoll, J.S.; Moore, M.J.; Moore, K.M.; Harry, C.T.; Hoppe, J.M.; Niemeyer, M.E.; Robinson, I.; Rose, K.S.; Sharp, W.B.; et al. Hematological, biochemical, and morphological parameters as prognostic indicators for stranded common dolphins (Delphinus delphis) from Cape Cod, Massachusetts, U.S.A. Mar. Mammal Sci. 2014, 30, 864–887. [Google Scholar] [CrossRef]
- Bossart, G.D.; Reidarson, T.H.; Dierauf, L.A.; Duffield, D.A. Clinical pathology. In CRC Handbook of Marine Mammal Medicine, 2nd ed.; Dierauf, L.A., Gulland, F.M.D., Eds.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Kim, B.R.; Park, S.J.; Shin, H.S.; Jung, Y.S.; Rim, H. Correlation between peripheral venous and arterial blood gas measurements in patients admitted to the intensive care unit: A single center study. Kidney Res. Clin. Pract. 2013, 32, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Bloom, B.M.; Grundlingh, J.; Bestwick, J.; Harris, T. The role of venous blood gas in the Emergency Department: A systematic review and meta-analysis. Eur. J. Emerg. Med. 2014, 21, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, K.; Kutter, A.P.; Schefer, R.J.; Marly-Voquer, C.; Sigrist, N. Determination of reference intervals and comparison of venous blood gas parameters using standard and non-standard collection methods in 24 cats. J. Feline Med. Surg. 2017, 19, 831–840. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BB | Navy | Total | ||
|---|---|---|---|---|
| Sex | Male | 25 | 14 | 39 |
| Female | 34 | 16 | 50 | |
| Age/Length Class | Juvenile | 33 | 8 | 41 |
| Subadult | 13 | 4 | 17 | |
| Adult | 13 | 18 | 31 | |
| Pregnancy | Pregnant Female | 10 | 5 | 15 |
| Non-Pregnant Female | 24 | 11 | 35 | |
| Ultrasound Lung Score | Normal–Mild | 40 | 30 | 70 |
| Moderate–Severe | 19 | 0 | 19 |
| Pulmonary Ultrasound Scores | Normal | Mild | Moderate | Severe |
|---|---|---|---|---|
| Navy | 25 | 5 | 0 | 0 |
| BB | 4 | 36 | 16 | 3 |
| Cohort | BB by Lung Score Category | ||||
|---|---|---|---|---|---|
| All Dolphins | Navy (All Normal–Mild Lung Score) | BB | BB Normal–Mild Lung Score | BB Moderate–Severe Lung Score | |
| Base Excess (mEq/L) | −1.0 | −0.6 | −2.0 | −3.8 # | 1.8 # |
| (−24–12) | (3.2) | (6.0) | (5.7) | (4.7) | |
| n = 89 | n = 30 | n = 59 | n = 40 | n = 19 | |
| Bicarbonate (mEq/L) | 25.6 | 26.0 | 24.9 | 23.6 # | 27.7 # |
| (8.9–36.9) | (2.9) | (4.7) | (4.5) | (3.8) | |
| n = 89 | n = 30 | n = 59 | n = 40 | n = 19 | |
| TCO2 (mmol/L) | 27.0 | 27.6 | 26.3 | 25.2 # | 28.6 # |
| (10–39) | (3.0) | (4.9) | (4.6) | (4.8) | |
| n = 89 | n = 30 | n = 59 | n = 40 | n = 19 | |
| Lactate (mmol/L) | 4.0 | 1.3 * | 7.3 * | 8.1 | 5.0 |
| (0.3–20.0) | (0.7) | (1.2–20.0) | (1.7–20) | (1.2–12.7) | |
| n = 89 | n = 30 | n = 59 | n = 40 | n = 19 | |
| PCO2 (mmHg) | 53.4 | 53.8 | 53.3 | 54.1 | 51.4 |
| (5.7) | (5.2) | (6.0) | (5.8) | (6.2) | |
| n = 89 | n = 30 | n = 59 | n = 40 | n = 19 | |
| PO2 (mmHg) | 62.8 | 51.4 * | 68.0 * | 68.0 | 68.0 |
| (18.0) | (18.1) | (46–116) | (47.0–116.0) | (46.0–97.0) | |
| n = 88 | n = 30 | n = 58 | n = 39 | n = 19 | |
| pH | 7.294 | 7.289 | 7.279 | 7.243 # | 7.333 # |
| (6.903–7.456) | (0.037) | (6.903–7.456) | (6.903–7.393) | (7.162–7.456) | |
| n = 89 | n = 30 | n = 59 | n = 40 | n = 19 | |
| ETCO2 (mmHg) | 46.8 | 40.6 * | 51.9 * | 52.0 | 51.6 |
| (31.2–75.2) | (33.4–65.4) | (9.7) | (9.2) | (10.9) | |
| n = 83 | n = 30 | n = 53 | n = 36 | n = 17 | |
| Lung Score | ||
|---|---|---|
| Normal–Mild (n = 24) | Moderate–Severe (n = 18) | |
| Δ ETCO2 | 3.9 | 2.1 |
| Δ PCO2 | 0.9 | 0 |
| Δ HCO3 | 3.7 | 1.6 |
| Δ Lactate | −4.2 | −2.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharp, S.M.; Gomez, F.M.; Meegan, J.M.; Rowles, T.K.; Townsend, F.; Schwacke, L.H.; Smith, C.R. Using Blood Gas Analysis and Capnography to Determine Oxygenation Status in Bottlenose Dolphins (Tursiops truncatus) Following the Deepwater Horizon Oil Spill. Toxics 2023, 11, 423. https://doi.org/10.3390/toxics11050423
Sharp SM, Gomez FM, Meegan JM, Rowles TK, Townsend F, Schwacke LH, Smith CR. Using Blood Gas Analysis and Capnography to Determine Oxygenation Status in Bottlenose Dolphins (Tursiops truncatus) Following the Deepwater Horizon Oil Spill. Toxics. 2023; 11(5):423. https://doi.org/10.3390/toxics11050423
Chicago/Turabian StyleSharp, Sarah M., Forrest M. Gomez, Jenny M. Meegan, Teresa K. Rowles, Forrest Townsend, Lori H. Schwacke, and Cynthia R. Smith. 2023. "Using Blood Gas Analysis and Capnography to Determine Oxygenation Status in Bottlenose Dolphins (Tursiops truncatus) Following the Deepwater Horizon Oil Spill" Toxics 11, no. 5: 423. https://doi.org/10.3390/toxics11050423
APA StyleSharp, S. M., Gomez, F. M., Meegan, J. M., Rowles, T. K., Townsend, F., Schwacke, L. H., & Smith, C. R. (2023). Using Blood Gas Analysis and Capnography to Determine Oxygenation Status in Bottlenose Dolphins (Tursiops truncatus) Following the Deepwater Horizon Oil Spill. Toxics, 11(5), 423. https://doi.org/10.3390/toxics11050423