
Effects of Decamethylcyclopentasiloxane on Reproductive Systems in Female Rats




Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animal and Chemical Treatments
2.3. Cell Culture
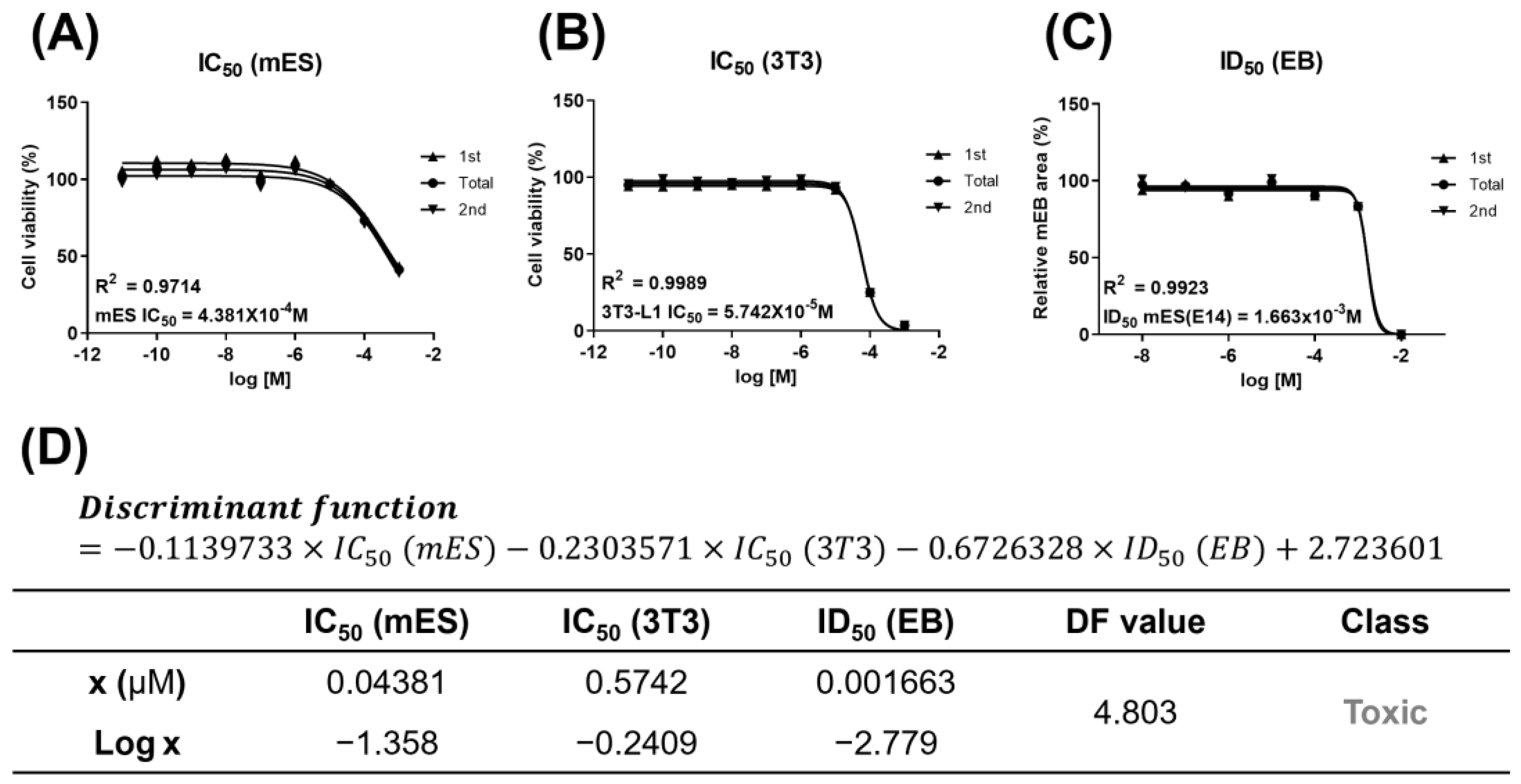
2.4. Embryoid Body Test (EBT)
2.5. Tissue Fixation and Section
2.6. RNA Extraction, Complementary DNA Synthesis, and Quantitative Real-Time PCR
2.7. Blood Collection and Serum Separation
2.8. Statistical Analyses
3. Results
3.1. Developmental Toxicity of D5
3.2. Organ Weights and Polycystic Ovarian Morphology Induced by Long-Term Exposure to D5
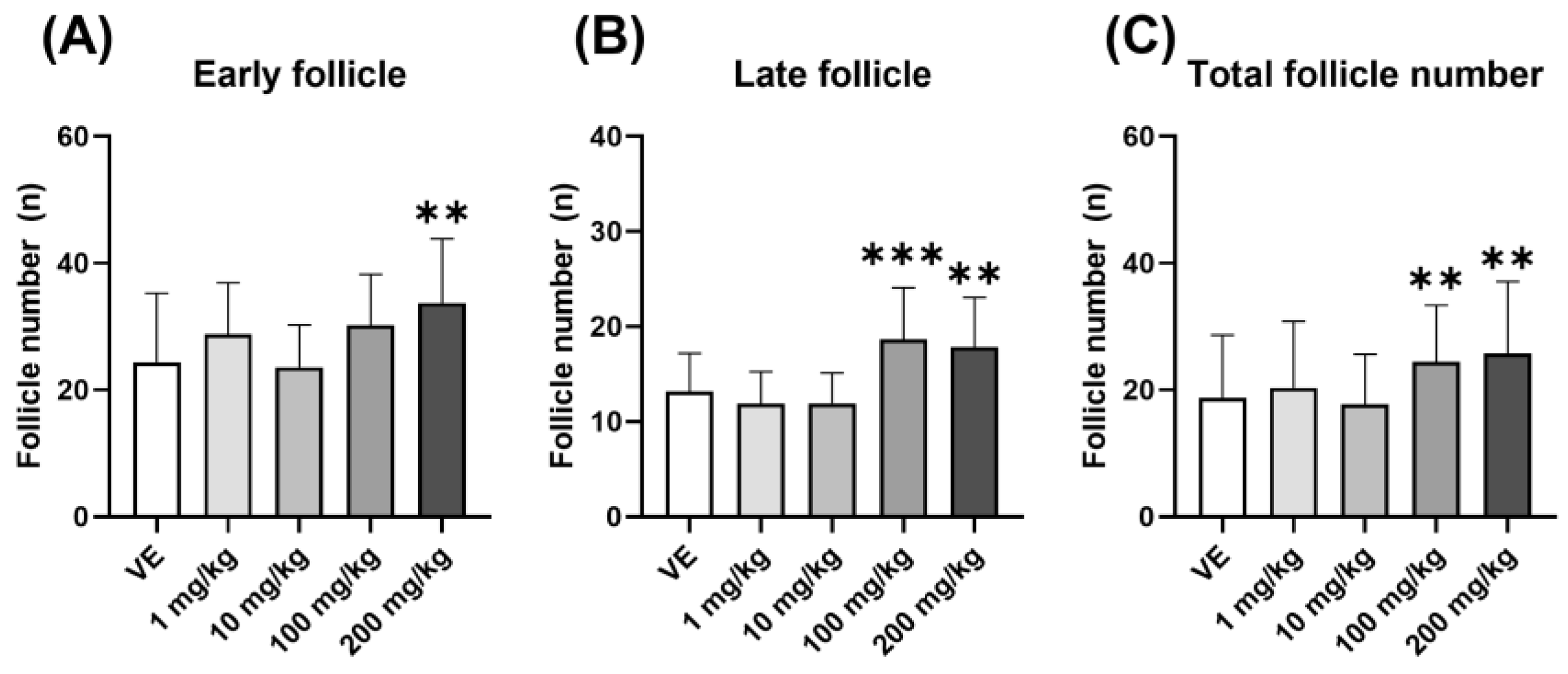
3.2.1. Polycystic Ovarian Morphology Induced by D5
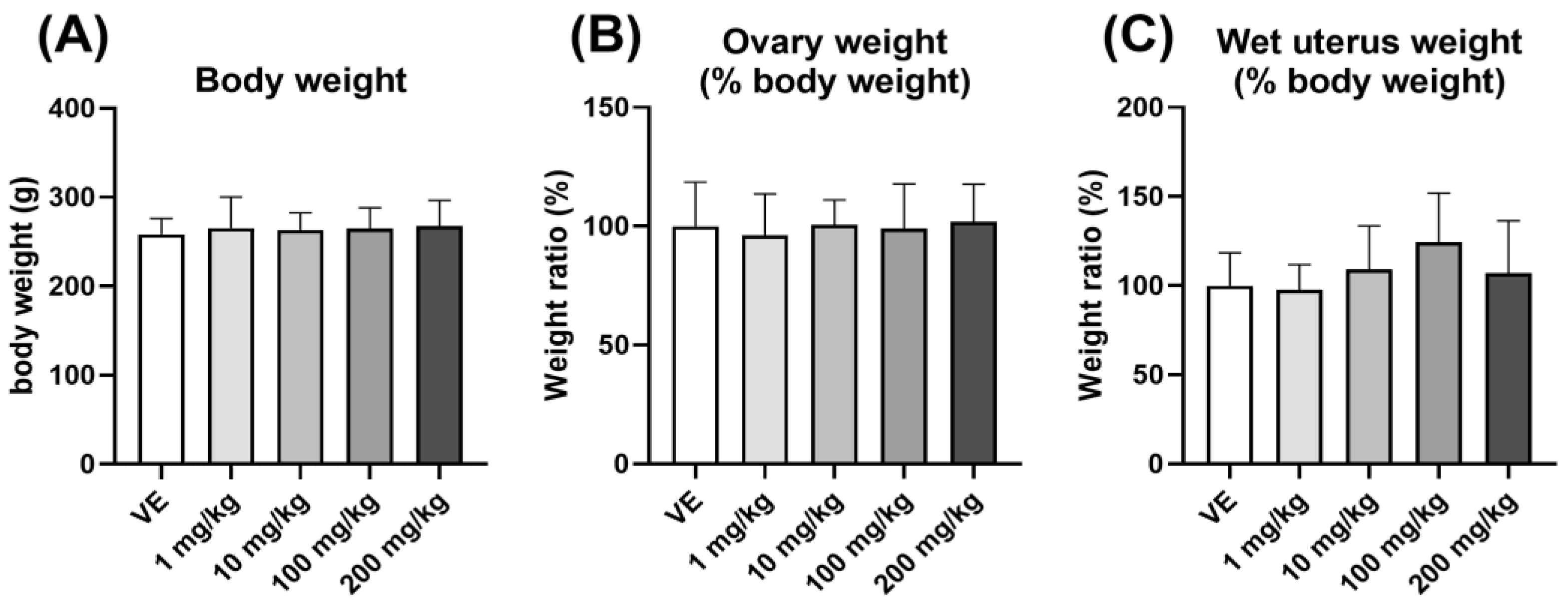
3.2.2. Weight of Uterus and Ovary Compared to Body Weight
3.3. Hormonal Changes in Serum When Exposed to D5
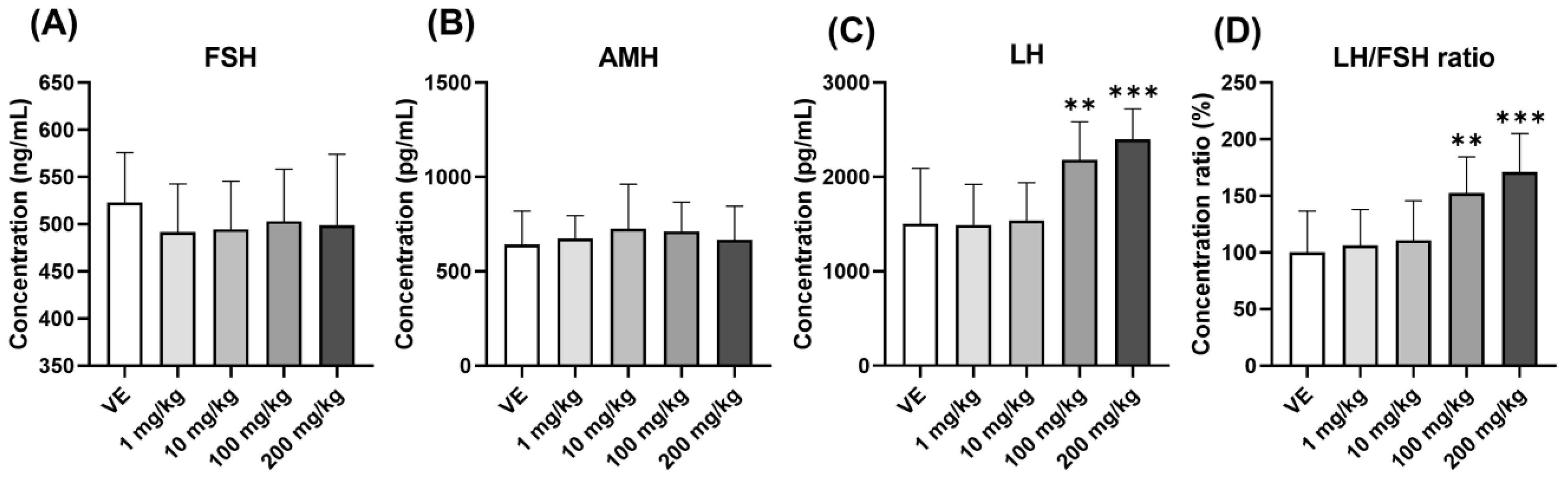
3.3.1. Changes in Gonadotropin Hormones and Anti-Müllerian Hormone
3.3.2. Changes in Steroid Hormones
3.4. Expression Levels of the Gonadotropin Hormone Receptor and the Steroid Hormone Receptor in Reproductive Organs
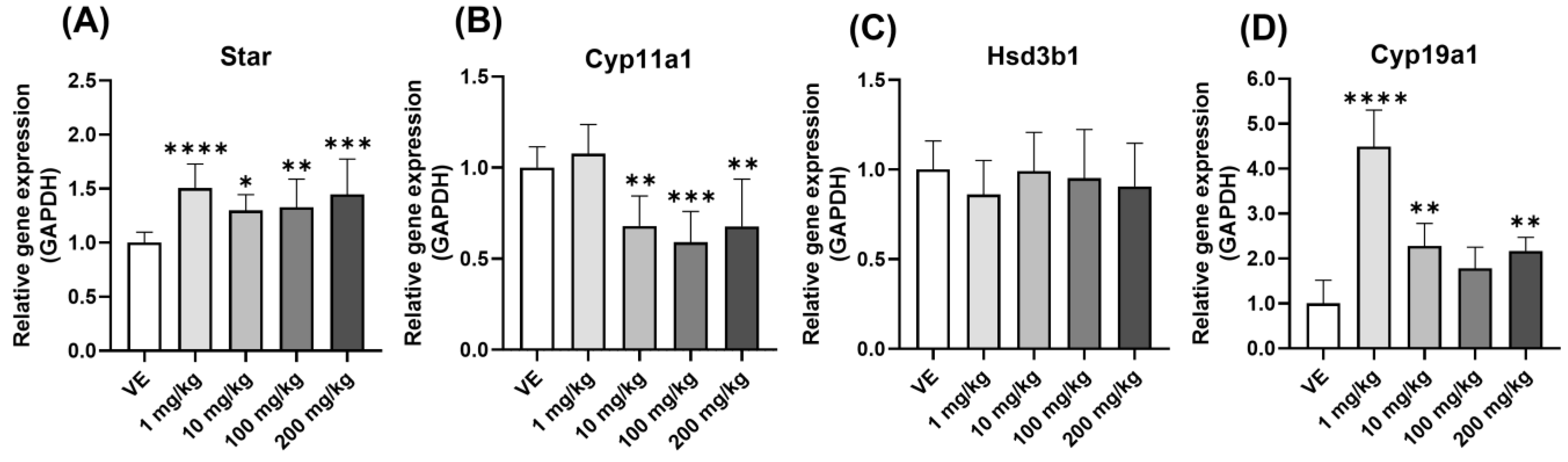
3.5. Comparison of Gene Expression in Ovary
3.5.1. Steroidogenesis Gene
3.5.2. Folliculogenesis Gene
3.6. Confirmation of Implantation-Related Gene Expression in the Uterus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silva, M.S.B.; Giacobini, P. New insights into anti-Müllerian hormone role in the hypothalamic-pituitary-gonadal axis and neuroendocrine development. Cell Mol. Life Sci. 2021, 78, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jaime, J.; Moenter, S.M. GnRH neuron excitability and action potential properties change with development but are not affected by prenatal androgen exposure. eNeuro 2022, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.W.; Sung, Y.-W.; Hsueh, Y.-W.; Chen, Y.-Y.; Ho, M.; Hsu, H.-C.; Yang, T.-C.; Lin, W.-C.; Chang, H.-M. Growth hormone in fertility and infertility: Mechanisms of action and clinical applications. Front. Endocrinol. 2022, 13, 1040503. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhai, J.; Xu, B.; Liu, J.; Chu, W.; Wang, D.; Geng, X.; Chen, Z.-J.; Du, Y. Erythropoietin-producing hepatocellular receptor A7 restrains estrogen negative feedback of luteinizing hormone via ephrin A5 in the hypothalamus of female rats. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E81–E90. [Google Scholar] [CrossRef]
- Lin, W.Y.; Tu, C.-P.; Kuo, H.-H.; Kuo, H.-W. Urinary Malondialdehyde (MDA) and N-Acetyl-β-D-Glucosaminidase (NAG) Associated with Exposure to Trichloroethylene (TCE) in Underground Water. Toxics 2022, 10, 293. [Google Scholar] [CrossRef]
- Otieno, J.; Kowal, P.; Mąkinia, J. Monitoring Lead Concentration in the Surrounding Environmental Components of a Lead Battery Company: Plants, Air and Effluents-Case Study, Kenya. Int. J. Environ. Res. Public Health 2022, 19, 5195. [Google Scholar] [CrossRef]
- Reutova, N.V.; Reutova, T.; Dreeva, F.R.; Shevchenko, A.A. Long-term impact of the Tyrnyauz tungsten-molybdenum mining and processing factory waste on environmental pollution and children’s population. Environ. Geochem. Health 2022, 44, 4557–4568. [Google Scholar] [CrossRef]
- Dong, H.; Lu, G.; Wang, X.; Zhang, P.; Yang, H.; Yan, Z.; Liu, J.; Jiang, R. Tissue-specific accumulation, depuration, and effects of perfluorooctanoic acid on fish: Influences of aqueous pH and sex. Sci. Total Environ. 2022, 861, 160567. [Google Scholar] [CrossRef]
- Gkotsis, G.; Nika, M.-C.; Nikolopoulou, V.; Alygizakis, N.; Bizani, E.; Aalizadeh, R.; Badry, A.; Chadwick, E.; Cincinelli, A.; Claßen, D.; et al. Assessment of contaminants of emerging concern in European apex predators and their prey by LC-QToF MS wide-scope target analysis. Environ. Int. 2022, 170, 107623. [Google Scholar] [CrossRef]
- Tabezar, N.; Sadeghi, P.; Fariman, G.A. Monsoon Effect on Heavy Metal and Chemical Composition in Parastromateus niger of the Oman Sea: Health Risk Assessment of Fish Consumption. Biol. Trace Elem. Res. 2022. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, H.; Namat, A.; Liu, J.; Ruan, F.; Xu, S.; Li, R.; Xia, W. Association between trimester-specific exposure to thirteen endocrine disrupting chemicals and preterm birth: Comparison of three statistical models. Sci. Total Environ. 2022, 851 Pt 2, 158236. [Google Scholar] [CrossRef]
- Lee, I.; Ji, K. Identification of combinations of endocrine disrupting chemicals in household chemical products that require mixture toxicity testing. Ecotoxicol. Environ. Saf. 2022, 240, 113677. [Google Scholar] [CrossRef]
- Mirza, F.G.; Tahlak, M.A.; Rjeili, R.B.; Hazari, K.; Ennab, F. Polycystic Ovarian Syndrome (PCOS): Does the Challenge End at Conception? Int. J. Environ. Res. Public Health 2022, 19, 14914. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Huo, P.; Liu, S.; Huang, H.; Zhang, S. Posttranslational modifications in pathogenesis of PCOS. Front. Endocrinol. 2022, 13, 1024320. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, Q.; Hu, R.; Li, R.; Yang, T. Five immune-related genes as diagnostic markers for endometriosis and their correlation with immune infiltration. Front. Endocrinol. 2022, 13, 1011742. [Google Scholar] [CrossRef]
- van Hoesel, M.H.; Chen, Y.L.; Zheng, A.; Wan, Q.; Mourad, S.M. Selective oestrogen receptor modulators (SERMs) for endometriosis. Cochrane Database Syst. Rev. 2021, 5, Cd011169. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; Mateen, S.; Ahmad, R.; Moin, S. A brief insight into the etiology, genetics, and immunology of polycystic ovarian syndrome (PCOS). J. Assist. Reprod. Genet. 2022, 39, 2439–2473. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, H.; Kong, S.; Wang, S.; Wang, H.; Wang, H.; Armant, D.R. Physiological and molecular determinants of embryo implantation. Mol. Asp. Med. 2013, 34, 939–980. [Google Scholar] [CrossRef]
- Balasch, J. Gonadotrophin ovarian stimulation and intrauterine insemination for unexplained infertility. Reprod. Biomed. Online 2004, 9, 664–672. [Google Scholar] [CrossRef]
- Marquardt, R.M.; Kim, T.H.; Shin, J.-H.; Jeong, J.-W. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef]
- Omura, T.; Morohashi, K. Gene regulation of steroidogenesis. J. Steroid Biochem. Mol. Biol. 1995, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Viru, A. Adaptive regulation of hormone interaction with receptor. Exp. Clin. Endocrinol. 1991, 97, 13–28. [Google Scholar] [CrossRef]
- Wang, D.G.; Norwood, W.; Alaee, M.; Byer, J.D.; Brimble, S. Review of recent advances in research on the toxicity, detection, occurrence and fate of cyclic volatile methyl siloxanes in the environment. Chemosphere 2013, 93, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Dudzina, T.; von Goetz, N.; Bogdal, C.; Biesterbos, J.W.H.; Hungerühler, K. Concentrations of cyclic volatile methylsiloxanes in European cosmetics and personal care products: Prerequisite for human and environmental exposure assessment. Environ. Int. 2014, 62, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Badjagbo, K.; Héroux, M.; Alaee, M.; Moore, S.; Sauvé, S. Quantitative analysis of volatile methylsiloxanes in waste-to-energy landfill biogases using direct APCI-MS/MS. Environ. Sci. Technol. 2010, 44, 600–605. [Google Scholar] [CrossRef]
- Wang, D.G.; Steer, H.; Tait, T.; Williams, Z.; Pacepavicius, G.; Young, T.; Ng, T.; Smyth, S.A.; Kinsman, L.; Alaee, M. Concentrations of cyclic volatile methylsiloxanes in biosolid amended soil, influent, effluent, receiving water, and sediment of wastewater treatment plants in Canada. Chemosphere 2013, 93, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Tan, L.; Xie, L.; Wang, Y.; Ellis, T. Investigation of volatile methyl siloxanes in biogas and the ambient environment in a landfill. J. Environ. Sci. 2020, 91, 54–61. [Google Scholar] [CrossRef]
- Riley, P.; Anson-Cartwright, L.; Cross, J.C. The Hand1 bHLH transcription factor is essential for placentation and cardiac morphogenesis. Nat. Genet. 1998, 18, 271–275. [Google Scholar] [CrossRef]
- Srisupundit, K.; Luewan, S.; Tongsong, T. Prenatal Diagnosis of Fetal Heart Failure. Diagnostics 2023, 13, 779. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.Y.; Choi, Y.-K.; Jo, N.R.; Lee, J.-H.; Ahn, C.; Ahn, I.Y.; Kim, T.S.; Kim, K.-S.; Choi, K.-C.; Lee, J.K.; et al. Advanced developmental toxicity test method based on embryoid body’s area. Reprod. Toxicol. 2017, 72, 74–85. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, S.Y.; Ahn, C.; Kim, C.-Y.; Kim, J.-E.; Jo, N.R.; Kang, H.Y.; Yoo, Y.-M.; Jung, E.-M.; Kim, M.K.; et al. Pre-validation study of alternative developmental toxicity test using mouse embryonic stem cell-derived embryoid bodies. Food Chem. Toxicol. 2019, 123, 50–56. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, S.Y.; Ahn, C.; Yoo, Y.-M.; Kim, C.-W.; Kim, J.-E.; Jo, N.R.; Kang, H.Y.; Jung, E.-M.; Kim, K.-S.; et al. Second-phase validation study of an alternative developmental toxicity test using mouse embryonic stem cell-derived embryoid bodies. J. Physiol. Pharmacol. 2020, 71, 223–233. [Google Scholar] [CrossRef]
- Rousselle, D.C. Opinion of the Scientific Committee on Consumer Safety (SCCS)—Final version of the opinion on decamethylcyclopentasiloxane (cyclopentasiloxane, D5) in cosmetic products. Regul. Toxicol. Pharmacol. 2017, 83, 117–118. [Google Scholar] [CrossRef] [PubMed]
- Meeks, R.G.; Stump, D.G.; Siddiqui, W.H.; Holson, J.F.; Plotzke, K.P.; Reynolds, V.L. An inhalation reproductive toxicity study of octamethylcyclotetrasiloxane (D4) in female rats using multiple and single day exposure regimens. Reprod. Toxicol. 2007, 23, 192–201. [Google Scholar] [CrossRef]
- Quinn, A.L.; Dalu, A.; Meeker, L.S.; Jean, P.A.; Meeks, R.G.; Crissman, J.W.; Gallavan, R.H., Jr.; Plotzke, K.P. Effects of octamethylcyclotetrasiloxane (D4) on the luteinizing hormone (LH) surge and levels of various reproductive hormones in female Sprague-Dawley rats. Reprod. Toxicol. 2007, 23, 532–540. [Google Scholar] [CrossRef]
- Siddiqui, W.H.; Stump, D.G.; Plotzke, K.P.; Holson, J.F.; Meeks, R.G. A two-generation reproductive toxicity study of octamethylcyclotetrasiloxane (D4) in rats exposed by whole-body vapor inhalation. Reprod. Toxicol. 2007, 23, 202–215. [Google Scholar] [CrossRef]
- Takizawa, K.; Mattison, D.R. Female reproduction. Am. J. Ind. Med. 1983, 4, 17–30. [Google Scholar] [CrossRef]
- Varaprath, S.; McMahon, J.M.; Plotzke, K.P. Metabolites of hexamethyldisiloxane and decamethylcyclopentasiloxane in Fischer 344 rat urine--a comparison of a linear and a cyclic siloxane. Drug Metab. Dispos. 2003, 31, 206–214. [Google Scholar] [CrossRef]
- Domoradzki, J.Y.; Sushynski, C.M.; Sushynski, J.M.; McNett, D.A.; Van Landingham, C.; Plotzke, K.P. Metabolism and disposition of [(14)C]-methylcyclosiloxanes in rats. Toxicol. Lett. 2017, 279 (Suppl. 1), 98–114. [Google Scholar] [CrossRef]
- Lim, J.J.; Lima, P.D.A.; Salehi, R.; Lee, D.R.; Tsang, B.K. Regulation of androgen receptor signaling by ubiquitination during folliculogenesis and its possible dysregulation in polycystic ovarian syndrome. Sci. Rep. 2017, 7, 10272. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Rajareddy, S.; Liu, L.; Jagarlamudi, K.; Boman, K.; Selstam, G.; Reddy, P. Control of mammalian oocyte growth and early follicular development by the oocyte PI3 kinase pathway: New roles for an old timer. Dev. Biol. 2006, 299, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Emori, C.; Kobayashi, M.; Maruyama, N.; Fujii, W.; Naito, K.; Sugiura, K. Cooperative effects of oocytes and estrogen on the forkhead box L2 expression in mural granulosa cells in mice. Sci. Rep. 2022, 12, 20158. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, D.; Ovitt, C.E.; Anlag, K.; Fehsenfeld, S.; Gredsted, L.; Treier, A.-C.; Treier, M. The murine winged-helix transcription factor Foxl2 is required for granulosa cell differentiation and ovary maintenance. Development 2004, 131, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Yuan, D.; Rajkovic, A. Germ cell-specific transcriptional regulator sohlh2 is essential for early mouse folliculogenesis and oocyte-specific gene expression. Biol. Reprod. 2008, 79, 1176–1182. [Google Scholar] [CrossRef]
- Murata, H.; Tsuzuki, T.; Kido, T.; Kakita-Kobayashi, M.; Kida, N.; Hisamatsu, Y.; Okada, H. Progestin-induced heart and neural crest derivatives-expressed transcript 2 inhibits angiopoietin 2 via fibroblast growth factor 9 in human endometrial stromal cells. Reprod. Biol. 2019, 19, 14–21. [Google Scholar] [CrossRef]
- Li, Q.; Davila, J.; Bagchi, M.K.; Bagchi, I.C. Chronic exposure to bisphenol a impairs progesterone receptor-mediated signaling in the uterus during early pregnancy. Recept. Clin. Investig. 2016, 3, e1369. [Google Scholar] [CrossRef]
- Ekanayake, D.L.; Maloplska, M.M.; Schwarz, T.; Tuz, R.; Bartlewski, P.M. The roles and expression of HOXA/Hoxa10 gene: A prospective marker of mammalian female fertility? Reprod. Biol. 2022, 22, 100647. [Google Scholar] [CrossRef]
- Wang, N.; Geng, L.; Zhang, S.; He, B.; Wang, J. Expression of PRB, FKBP52 and HB-EGF relating with ultrasonic evaluation of endometrial receptivity. PLoS ONE 2012, 7, e34010. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, Y.; Edelshain, B.; Schatz, F.; Lockwood, C.J.; Taylor, H.S. FKBP4 is regulated by HOXA10 during decidualization and in endometriosis. Reproduction 2012, 143, 531–538. [Google Scholar] [CrossRef]
- Chappel, S.C.; Howles, C. Reevaluation of the roles of luteinizing hormone and follicle-stimulating hormone in the ovulatory process. Hum. Reprod. 1991, 6, 1206–1212. [Google Scholar] [CrossRef]
- Johansson, J.; Stener-Victorin, E. Polycystic ovary syndrome: Effect and mechanisms of acupuncture for ovulation induction. Evid. Based Complement Altern. Med. 2013, 2013, 762615. [Google Scholar] [CrossRef] [PubMed]
- Beato, M.; Klug, J. Steroid hormone receptors: An update. Hum. Reprod. Update 2000, 6, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Kanamarlapudi, V.; Gordon, U.D.; Bernal, A.L. Luteinizing hormone/chorionic gonadotrophin receptor overexpressed in granulosa cells from polycystic ovary syndrome ovaries is functionally active. Reprod. Biomed. Online 2016, 32, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Schniewind, H.A.; Sattler, L.-M.; Haudum, C.W.; Műnzker, J.; Waldemar, B.M.; Obermayer-Pietsch, B.; Schomburg, L. Autoimmunity to the Follicle-Stimulating Hormone Receptor (FSHR) and Luteinizing Hormone Receptor (LHR) in Polycystic Ovarian Syndrome. Int. J. Mol. Sci. 2021, 22, 13667. [Google Scholar] [CrossRef] [PubMed]
- Stocco, D.M. The role of the StAR protein in steroidogenesis: Challenges for the future. J. Endocrinol. 2000, 164, 247–253. [Google Scholar] [CrossRef]
- Lin, Y.C.; Papadopoulos, V. Neurosteroidogenic enzymes: CYP11A1 in the central nervous system. Front. Neuroendocrinol. 2021, 62, 100925. [Google Scholar] [CrossRef] [PubMed]
- Sechman, A.; Grzegorzewska, A.; Grzesiak, M.; Kozubek, A.; Katarzyńska-Banasik, D.; Kowalik, K.; Hrabia, A. Nitrophenols suppress steroidogenesis in prehierarchical chicken ovarian follicles by targeting STAR, HSD3B1, and CYP19A1 and downregulating LH and estrogen receptor expression. Domest. Anim. Endocrinol. 2020, 70, 106378. [Google Scholar] [CrossRef]
- Huang, X.; Wu, B.; Chen, M.; Hong, L.; Kong, P.; Wei, Z.; Teng, X. Depletion of exosomal circLDLR in follicle fluid derepresses miR-1294 function and inhibits estradiol production via CYP19A1 in polycystic ovary syndrome. Aging 2020, 12, 15414–15435. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, E.M.; An, B.S.; Hwang, I.; Vo, T.T.; Lee, S.M.; Choi, K.C.; Jeung, E.B. Additional effects of bisphenol A and paraben on the induction of calbindin-D(9K) and progesterone receptor via an estrogen receptor pathway in rat pituitary GH3 cells. J. Physiol. Pharmacol. 2012, 63, 445–455. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| Pgr | TGGTTCCGCCACTCATCA | TGGTCAGCAAAGAGCTGGAAG |
| Esr1 | GACTTGAATCTCCACGATCA | CTTCAAGGTGCTGGATAGAA |
| Esr2 | TCCGGCTCTTAGAAAGCTGC | CCCCTCATCCCTGTCCAGAA |
| Fshr | CTTGAAGCGGCAAATCTCTG | GAGCAGGTCACATCAACAAC |
| Lhcgr | CTCACTGAAAACACTGCCCT | ATGGCGGAATAAAGCGTCTC |
| Star | GCGGAACATGAAAGGACTGA | TCCTTGCTGGATGTAGGACA |
| Cyp11a1 | GCTTTGCCTTTGAGTCCATC | CATGGTCCTTCCAGGTCTTA |
| Hsd3b1 | TGCCACTTGGTCACACTGTCA | CCCTGTGCTGCTCCACTAGTGT |
| Cyp19a1 | GGCAAGCACTCCTTATCAAACC | TCCACGTCTCTCAGCGAAAA |
| Amh | CTGGCTGAAGTGATATGGGA | CACAGTCAGCACCAAATAGC |
| Sohlh2 | AGCCAGCTCCAGTTGTCTGT | GATGCTGGATGAGGCAGT |
| Kitlg | TTCAAGGACTTCATGGTGGC | GCGGCTTTCCTATTACTGCT |
| Foxl2 | TCGCTAAGTTCCCGTTCTAC | GTAATTGCCCTTCTCGAACA |
| Hand2 | CCACCAGCTACATCGCCTAC | CTGTCCGGCCTTTGGTTTTC |
| Fkbp4 | TTCCAGATCCCACCACATGC | TCCTTGAAGTACACGGTGCC |
| Hoxa10 | GCTGGGGTGCACATCATAAA | TGCCTCAAAGTGGCAGTCG |
| Gapdh * | GGCAAGTTCAATGGCACAGT | TGGTGAAGACGCCAGTAGACTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Kim, K.; Park, S.-M.; Kwon, J.-S.; Jeung, E.-B. Effects of Decamethylcyclopentasiloxane on Reproductive Systems in Female Rats. Toxics 2023, 11, 302. https://doi.org/10.3390/toxics11040302
Lee J, Kim K, Park S-M, Kwon J-S, Jeung E-B. Effects of Decamethylcyclopentasiloxane on Reproductive Systems in Female Rats. Toxics. 2023; 11(4):302. https://doi.org/10.3390/toxics11040302
Chicago/Turabian StyleLee, Jimin, Kangmin Kim, Seon-Mi Park, Jin-Sook Kwon, and Eui-Bae Jeung. 2023. "Effects of Decamethylcyclopentasiloxane on Reproductive Systems in Female Rats" Toxics 11, no. 4: 302. https://doi.org/10.3390/toxics11040302
APA StyleLee, J., Kim, K., Park, S.-M., Kwon, J.-S., & Jeung, E.-B. (2023). Effects of Decamethylcyclopentasiloxane on Reproductive Systems in Female Rats. Toxics, 11(4), 302. https://doi.org/10.3390/toxics11040302