
Differential Cell Metabolic Pathways in Gills and Liver of Fish (White Seabream Diplodus sargus) Coping with Dietary Methylmercury Exposure

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
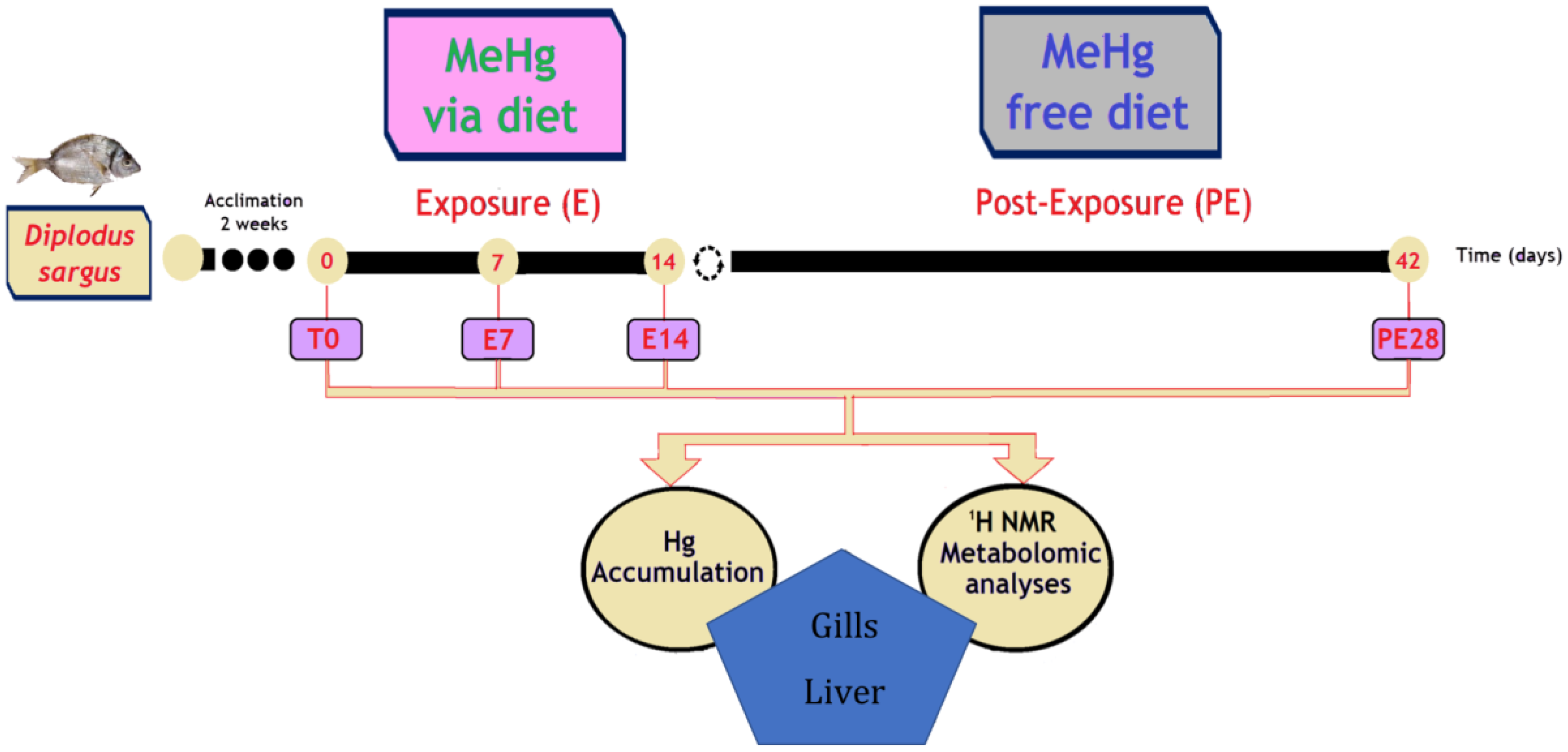
2.2. Experimental Set-Up and Sampling
2.3. Total Hg Determination
2.4. Metabolomics Analysis
2.4.1. Extraction of Metabolites
2.4.2. 1H NMR-Based Metabolomics and Spectral Pre-Processing
2.5. Data Statistical Analysis
3. Results
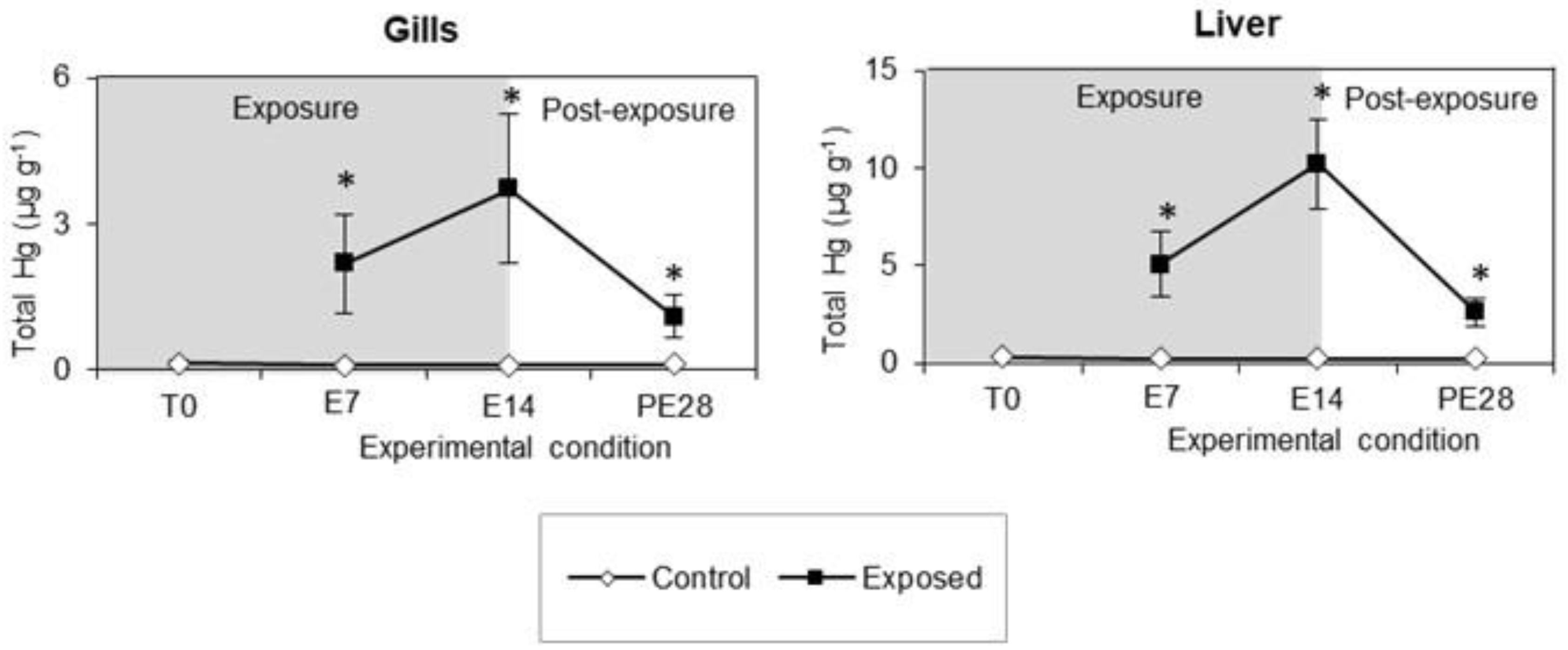
3.1. Total Hg Accumulation
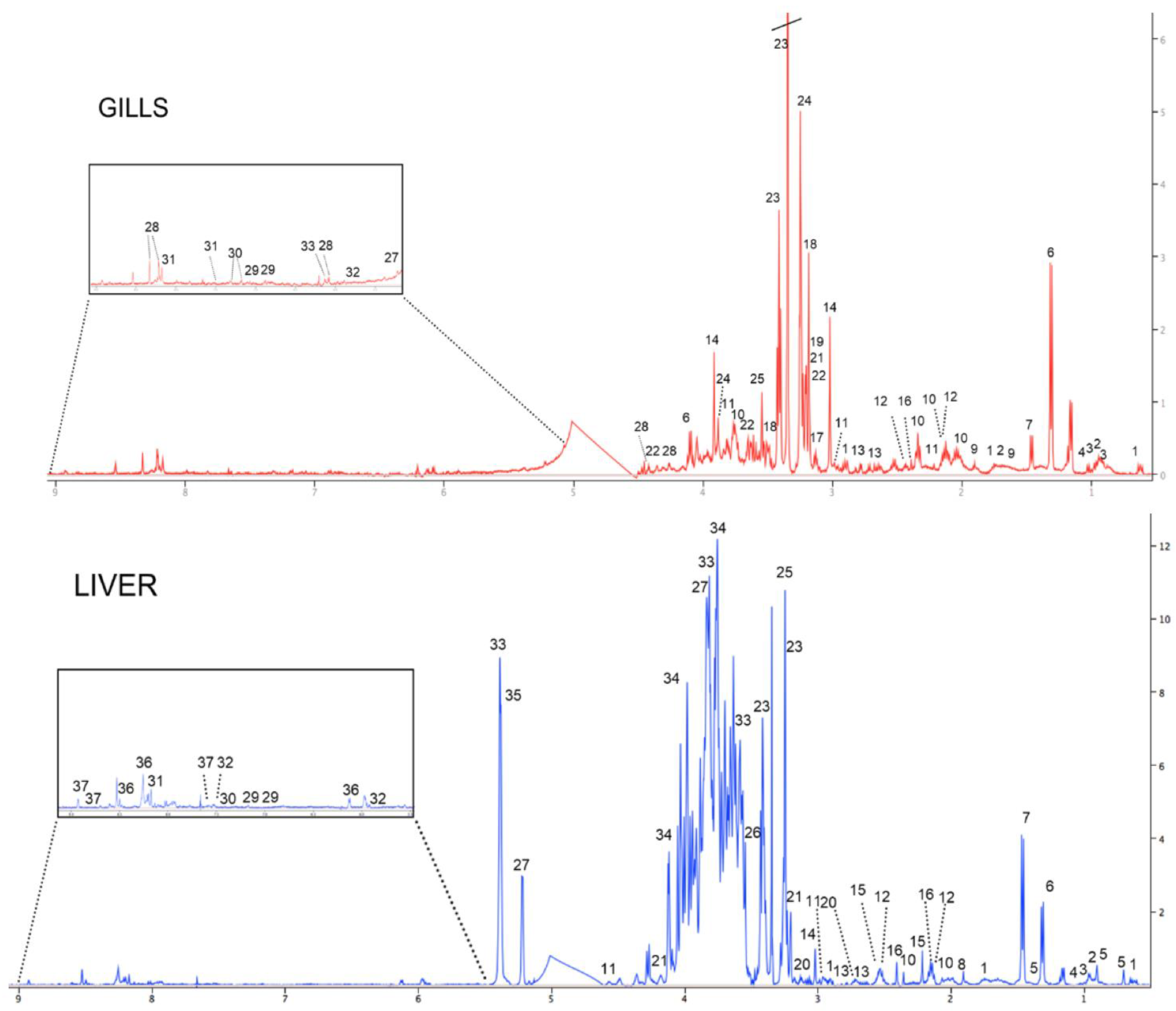
3.2. Metabolomics
3.2.1. 1H NMR Spectroscopy of Gill and Liver Extracts of Unexposed Fish
3.2.2. Metabolome of Gills in MeHg-Exposed Fish
3.2.3. Metabolome of Liver in MeHg-Exposed Fish
4. Discussion
4.1. MeHg-Induced Metabolome Changes in Fish Gills
4.2. MeHg-Induced Metabolome Changes in Fish Liver
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cao, L.; Liu, J.; Dou, S.; Huang, W. Biomagnification of methylmercury in a marine food web in Laizhou Bay (North China) and associated potential risks to public health. Mar. Pollut. Bull. 2020, 150, 110762. [Google Scholar] [CrossRef] [PubMed]
- Paduraru, E.; Iacob, D.; Rarinca, V.; Rusu, A.; Jijie, R.; Ilie, O.-D.; Ciobica, A.; Nicoara, M.; Doroftei, B. Comprehensive review regarding mercury poisoning and its complex involvement in Alzheimer’s disease. Int. J. Mol. Sci. 2022, 23, 1992. [Google Scholar] [CrossRef] [PubMed]
- Kerek, E.; Hassanin, M.; Prenner, E.J. Inorganic mercury and cadmium induce rigidity in eukaryotic lipid extracts while mercury also ruptures red blood cells. BBA–Biomembranes 2018, 1860, 710–717. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadegh, M. Toxic mechanisms of five heavy metals: Mercury, lead, chromium, cadmium, and arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef] [PubMed]
- Obrist, D.; Kirk, J.L.; Zhang, L.; Sunderland, E.M.; Jiskra, M.; Selin, N.E. A review of global environmental mercury processes in response to human and natural perturbations: Changes of emissions, climate, and land use. Ambio 2018, 47, 116–140. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Korbas, M.; Pereira, V.; Cappello, T.; Maisano, M.; Canario, J.; Almeida, A.; Pacheco, M. A multidimensional concept for mercury neuronal and sensory toxicity in fish—From toxicokinetics and biochemistry to morphometry and behavior. BBA–Gen. Subj. 2019, 1863, 129298. [Google Scholar] [CrossRef] [PubMed]
- Parrino, V.; Costa, G.; Giannetto, A.; De Marco, G.; Cammilleri, G.; Acar, Ü.; Piccione, G.; Fazio, F. Trace elements (Al, Cd, Cr, Cu, Fe, Mn, Ni, Pb and Zn) in Mytilus galloprovincialis and Tapes decussatus from Faro and Ganzirri Lakes (Sicily, Italy): Flow cytometry applied for hemocytes analysis. J. Trace Elem. Med. Biol. 2021, 68, 126870. [Google Scholar] [CrossRef]
- Polak-Juszczak, L. Distribution of organic and inorganic mercury in the tissues and organs of fish from the southern Baltic Sea. Environ. Sci. Pollut. Res. 2018, 25, 34181–34189. [Google Scholar] [CrossRef]
- Łuczyńska, J.; Łuczyński, M.J.; Nowosad, J.; Kowalska-Góralska, M.; Senze, M. Total mercury and fatty acids in selected fish species on the Polish market: A risk to human health. Int. J. Environ. Res. Public Health 2022, 19, 10092. [Google Scholar] [CrossRef]
- Mason, R.P.; Coulibaly, M.; Hansen, G.; Inman, H.; Myer, P.K.; Yao, K.M. An examination of mercury levels in the coastal environment and fish of Cote d’Ivoire. Chemosphere 2022, 300, 134609. [Google Scholar] [CrossRef]
- Olson, C.I.; Fakhraei, H.; Driscoll, C.T. Mercury emissions, atmospheric concentrations, and wet deposition across the conterminous United States: Changes over 20 years of monitoring. Environ. Sci. Technol. Lett. 2020, 7, 376–381. [Google Scholar] [CrossRef]
- Arcagni, M.; Juncos, R.; Rizzo, A.; Pavlin, M.; Fajon, V.; Arribere, M.A.; Horvat, M.; Guevara, S.R. Species- and habitat-specific bioaccumulation of total mercury and methylmercury in the food web of a deep oligotrophic lake. Sci. Total Environ. 2018, 612, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Lopez, L.; Bhavsar, S.P.; Sharma, S. What’s hot about mercury? Examining the influence of climate on mercury levels in Ontario top predator fishes. Environ. Res. 2018, 162, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, T.W. The pharmacology of mercury compounds. Annu. Rev. Pharmacol. 1972, 12, 375–406. [Google Scholar] [CrossRef]
- Taylor, D.L.; Calabrese, N.M. Mercury content of blue crabs (Callinectes sapidus) from southern New England coastal habitats: Contamination in an emergent fishery and risks to human consumers. Mar. Pollut. Bull. 2018, 126, 166–178. [Google Scholar] [CrossRef]
- Genchi, G.; Sinicropi, M.S.; Carocci, A.; Lauria, G.; Catalano, A. Mercury exposure and heart diseases. Int. J. Environ. Res. Public Health 2017, 14, 74. [Google Scholar] [CrossRef]
- Raihan, S.M.; Moniruzzaman, M.; Park, Y.; Lee, S.; Bai, S.C. Evaluation of dietary organic and inorganic mercury threshold levels on induced mercury toxicity in a marine fish model. Animals 2020, 10, 405. [Google Scholar] [CrossRef]
- Zulkipli, S.Z.; Liew, H.J.; Ando, M.; Lim, L.S.; Wang, M.; Sung, Y.Y.; Mok, W.J. A review of mercury pathological effects on organs specific of fishes. Environ. Pollut. Bioavailab. 2021, 33, 76–87. [Google Scholar] [CrossRef]
- Strungaru, S.A.; Robea, M.A.; Plavan, G.; Todirascu-Ciornea, E.; Ciobica, A.; Nicoara, M. Acute exposure to methylmercury chloride induces fast changes in swimming performance, cognitive processes and oxidative stress of zebrafish (Danio rerio) as reference model for fish community. J. Trace Elem. Med. Biol. 2018, 47, 115–123. [Google Scholar] [CrossRef]
- Brandão, F.; Cappello, T.; Raimundo, J.; Santos, M.A.; Maisano, M.; Mauceri, A.; Pacheco, M.; Pereira, P. Unravelling the mechanisms of mercury hepatotoxicity in wild fish (Liza aurata) through a triad approach: Bioaccumulation, metabolomic profiles and oxidative stress. Metallomics 2015, 7, 1352–1363. [Google Scholar] [CrossRef]
- Khadra, M.; Caron, A.; Dolors, P.; Ponton, D.E.; Rosabal, M.; Amyot, M. The fish or the egg: Maternal transfer and subcellular partitioning of mercury and selenium in Yellow Perch (Perca flavescens). Sci. Total Environ. 2019, 675, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Liu, X.; Wang, Z.; Wang, B.; Lin, C.; Xin, M.; Zhang, B.T.; Wu, T.; He, M.; Ouyang, W. Trophic transfer and dietary exposure risk of mercury in aquatic organisms from urbanized coastal ecosystems. Chemosphere 2021, 281, 130836. [Google Scholar] [CrossRef] [PubMed]
- Noureen, A.; De Marco, G.; Rehman, N.; Jabeen, F.; Cappello, T. Ameliorative hematological and histomorphological effects of dietary Trigonella foenum-graecum seeds in common carp (Cyprinus carpio) exposed to copper oxide nanoparticles. Int. J. Environ. Res. Public Health 2022, 19, 13462. [Google Scholar] [CrossRef] [PubMed]
- Parrino, V.; De Marco, G.; Minutoli, R.; Lo Paro, G.; Giannetto, A.; Cappello, T.; De Plano, L.M.; Cecchini, S.; Fazio, F. Effects of pesticides on Chelon labrosus (Risso, 1827) evaluated by enzymatic activities along the north eastern Sicilian coastlines (Italy). Eur. Zool. J. 2021, 88, 540–548. [Google Scholar] [CrossRef]
- Yuan, L.; Shi, X.; Tang, B.Z.; Wang, W.X. Real-time in vitro monitoring of the subcellular toxicity of inorganic Hg and methylmercury in zebrafish cells. Aquat. Toxicol. 2021, 236, 105859. [Google Scholar] [CrossRef]
- Zitouni, N.; Cappello, T.; Missawi, O.; Boughattas, I.; De Marco, G.; Belbekhouche, S.; Mokni, M.; Alphonse, V.; Guerbej, H.; Bousserrhine, N.; et al. Metabolomic disorders unveil hepatotoxicity of environmental microplastics in wild fish Serranus scriba (Linnaeus 1758). Sci. Total Environ. 2022, 838, 155872. [Google Scholar] [CrossRef]
- Giacalone, V.M.; Pipitone, C.; Abecasis, D.; Badalamenti, F.; D’Anna, G. Movement ecology of the white seabream Diplodus sargus across its life cycle: A review. Environ. Biol. Fish. 2022, 105, 1809–1823. [Google Scholar] [CrossRef]
- Merciai, R.; Rodriguez-Prieto, C.; Torres, J.; Casadevall, M. Bioaccumulation of mercury and other trace elements in bottom-dwelling omnivorous fishes: The case of Diplodus sargus (L.) (Osteichthyes: Sparidae). Mar. Pollut. Bull. 2018, 136, 10–21. [Google Scholar] [CrossRef]
- Cardoso, O.; Puga, S.; Brandão, F.; Canário, J.; O’Driscoll, N.J.; Santos, M.A.; Pacheco, M.; Pereira, P. Oxidative stress profiles in brain point out a higher susceptibility of fish to waterborne divalent mercury compared to dietary organic mercury. Mar. Pollut. Bull. 2017, 122, 110–121. [Google Scholar] [CrossRef]
- Puga, S.; Pereira, P.; Pinto-Ribeiro, F.; O’Driscoll, N.J.; Mann, E.; Barata, M.; Pousão-Ferreira, P.; Canário, J.; Almeida, A.; Pacheco, M. Unveiling the neurotoxicity of methylmercury in fish (Diplodus sargus) through a regional morphometric analysis of brain and swimming behavior assessment. Aquat. Toxicol. 2016, 180, 320–333. [Google Scholar] [CrossRef]
- Cappello, T. NMR-based metabolomics of aquatic organisms. eMagRes 2020, 9, 81–100. [Google Scholar] [CrossRef]
- Lin, C.Y.; Viant, M.R.; Tjeerdema, R.S. Metabolomics: Methodologies and applications in the environmental sciences. J. Pestic. Sci. 2006, 31, 245–251. [Google Scholar] [CrossRef]
- Caliani, I.; De Marco, G.; Cappello, T.; Giannetto, A.; Mancini, G.; Ancora, S.; Maisano, M.; Parrino, V.; Cappello, S.; Bianchi, N.; et al. Assessment of the effectiveness of a novel BioFilm-Membrane BioReactor oil-polluted wastewater treatment technology by applying biomarkers in the mussel Mytilus galloprovincialis. Aquat. Toxicol. 2022, 243, 106059. [Google Scholar] [CrossRef]
- Cappello, T.; Vitale, V.; Oliva, S.; Villari, V.; Mauceri, A.; Fasulo, S.; Maisano, M. Alteration of neurotransmission and skeletogenesis in sea urchin Arbacia lixula embryos exposed to copper oxide nanoparticles. Comp. Biochem. Physiol. C 2017, 199, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T.; De Marco, G.; Oliveri Conti, G.; Giannetto, A.; Ferrante, M.; Mauceri, A.; Maisano, M. Time-dependent metabolic disorders induced by short-term exposure to polystyrene microplastics in the Mediterranean mussel Mytilus galloprovincialis. Ecotoxicol. Environ. Saf. 2021, 209, 111780. [Google Scholar] [CrossRef] [PubMed]
- De Marco, G.; Brandão, F.; Pereira, P.; Pacheco, M.; Cappello, T. Organ-specific metabolome deciphering cell pathways to cope with mercury in wild fish (golden grey mullet Chelon auratus). Animals 2022, 12, 79. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, T.; Li, M.; Xu, H.; Jia, A.; Zhang, J.; Wang, J. 1H NMR based metabolomics approach to study the toxic effects of dichlorvos on goldfish (Carassius auratus). Chemosphere 2015, 138, 537–545. [Google Scholar] [CrossRef]
- Missawi, O.; Venditti, M.; Cappello, T.; Zitouni, N.; Marco, G.D.E.; Boughattas, I.; Bousserrhine, N.; Belbekhouche, S.; Minucci, S.; Maisano, M.; et al. Autophagic event and metabolomic disorders unveil cellular toxicity of environmental microplastics on marine polychaete Hediste diversicolor. Environ. Pollut. 2022, 302, 119106. [Google Scholar] [CrossRef]
- Xu, H.-D.; Wang, J.-S.; Li, M.-H.; Liu, Y.; Chen, T.; Jia, A.-Q. 1H NMR based metabolomics approach to study the toxic effects of herbicide butachlor on goldfish (Carassius auratus). Aquat. Toxicol. 2015, 159, 69–80. [Google Scholar] [CrossRef]
- Cappello, T.; Brandão, F.; Guilherme, S.; Santos, M.A.; Maisano, M.; Mauceri, A.; Canário, J.; Pacheco, M.; Pereira, P. Insights into the mechanisms underlying mercury-induced oxidative stress in gills of wild fish (Liza aurata) combining 1H NMR metabolomics and conventional biochemical assays. Sci. Total Environ. 2016, 548–549, 13–24. [Google Scholar] [CrossRef]
- Cappello, T.; Pereira, P.; Maisano, M.; Mauceri, A.; Pacheco, M.; Fasulo, S. Advances in understanding the mechanisms of mercury toxicity in wild golden grey mullet (Liza aurata) by 1H NMR-based metabolomics. Environ. Pollut. 2016, 219, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Pablo, H.D.; Vale, C.; Pacheco, M. Combined use of environmental data and biomarkers in fish (Liza aurata) inhabiting a eutrophic and metal contaminated coastal system—Gills reflect environmental contamination. Mar. Environ. Res. 2010, 69, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.H. The fish gill: Site of action and model for toxic effects of environmental pollutants. Environ. Health Perspect. 1987, 71, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Leaner, J.J.; Mason, R.P. Methylmercury uptake and distribution kinetics in sheepshead minnows, Cyprinodon variegatus, after exposure to CH3Hg-spiked food. Environ. Toxicol. Chem. 2004, 23, 2138–2146. [Google Scholar] [CrossRef]
- Ardeshir, R.A.; Movahedinia, A.A.; Rastgar, S. Fish liver biomarkers for heavy metal pollution: A review article. Am. J. Toxicol. 2017, 2, 1–8. [Google Scholar]
- EPA. Method 7473 (SW-846): Mercury in Solids and Solutions by Thermal Decomposition, Amalgamation, and Atomic Absorption Spectrophotometry; EPA: Washington, DC, USA, 1998. [Google Scholar]
- Costley, C.T.; Mossop, K.F.; Dean, J.R.; Garden, L.M.; Marshall, J.; Carrol, J. Determination of mercury in environmental and biological samples using pyrolysis atomic absorption spectrometry with gold amalgamation. Anal. Chim. Acta 2000, 405, 179–183. [Google Scholar] [CrossRef]
- Cappello, T.; Maisano, M.; Giannetto, A.; Natalotto, A.; Parrino, V.; Mauceri, A.; Spanò, N. Pen shell Pinna nobilis L. (Mollusca, Bivalvia) from different peculiar environments: Adaptive mechanisms of osmoregulation and neurotransmission. Eur. Zool. J. 2019, 86, 333–342. [Google Scholar] [CrossRef]
- Vignet, C.; Cappello, T.; Fu, Q.; Lajoie, K.; De Marco, G.; Clerandeau, C.; Mottaz, H.; Maisano, M.; Hollender, J.; Schirmer, K.; et al. Imidacloprid induces adverse effects on fish early life stages that are more severe in Japanese medaka (Oryzias latipes) than in zebrafish (Danio rerio). Chemosphere 2019, 225, 470–478. [Google Scholar] [CrossRef]
- Kostopoulou, S.; Ntatsi, G.; Arapis, G.; Aliferis, K.A. Assessment of the effects of metribuzin, glyphosate, and their mixtures on the metabolism of the model plant Lemna minor L. applying metabolomics. Chemosphere 2020, 239, 124582. [Google Scholar] [CrossRef]
- Tian, S.; Teng, M.; Meng, Z.; Yan, S.; Jia, M.; Li, R.; Liu, L.; Yan, J.; Zhou, Z.; Zhu, W. Toxicity effects in zebrafish embryos (Danio rerio) induced by prothioconazole. Environ. Pollut. 2019, 255, 113269. [Google Scholar] [CrossRef]
- Cappello, T.; Giannetto, A.; Parrino, V.; Maisano, M.; Oliva, S.; De Marco, G.; Guerriero, G.; Mauceri, A.; Fasulo, S. Baseline levels of metabolites in different tissues of mussel Mytilus galloprovincialis (Bivalvia: Mytilidae). Comp. Biochem. Physiol. D 2018, 26, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose metabolism in fish: A review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef] [PubMed]
- Ung, C.Y.; Lam, S.H.; Hlaing, M.M.; Winata, C.L.; Korzh, S.; Mathavan, S.; Gong, Z. Mercury-induced hepatotoxicity in zebrafish: In vivo mechanistic insights from transcriptome analysis, phenotype anchoring and targeted gene expression validation. BMC Genom. 2010, 11, 212. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Puga, S.; Cardoso, V.; Pinto-Ribeiro, F.; Raimundo, J.; Barata, M.; Pousão-Ferreira, P.; Pacheco, M.; Almeida, A. Inorganic mercury accumulation in brain following waterborne exposure elicits a deficit on the number of brain cells and impairs swimming behavior in fish (white seabream—Diplodus sargus). Aquat. Toxicol. 2016, 170, 400–412. [Google Scholar] [CrossRef]
- Xu, Q.; Zhao, L.; Wang, Y.; Xie, Q.; Yin, D.; Feng, X.; Wang, D. Bioaccumulation characteristics of mercury in fish in the Three Gorges Reservoir, China. Environ. Pollut. 2018, 243, 115–126. [Google Scholar] [CrossRef]
- Zheng, N.; Wang, S.; Dong, W.; Hua, X.; Li, Y.; Song, X.; Chu, Q.; Hou, S.; Li, Y. The toxicological effects of mercury exposure in marine fish. Bull. Environ. Contam. Toxicol. 2019, 102, 714–720. [Google Scholar] [CrossRef]
- Pereira, P.; Raimundo, J.; Barata, M.; Araujo, O.; Pousao-Ferreira, P.; Canario, J.; Almeida, A.; Pahceco, M. A new page on the road book of inorganic mercury in fish body—Tissue distribution and elimination following waterborne exposure and post-exposure periods. Metallomics 2015, 7, 525–535. [Google Scholar] [CrossRef]
- Calder, P.C. Branched-chain amino acids and immunity. J. Nutr. 2006, 136, 288S–293S. [Google Scholar] [CrossRef]
- Bucking, C. A broader look at ammonia production, excretion, and transport in fish: A review of impacts of feeding and the environment. J. Comp. Physiol. B 2017, 187, 1–18. [Google Scholar] [CrossRef]
- Reed, M.; Jonz, M.G. Neurochemical signalling associated with gill oxygen sensing and ventilation: A receptor focused mini-review. Front. Physiol. 2022, 13, 1396. [Google Scholar] [CrossRef]
- Pan, W.; Godoy, R.S.; Cook, D.P.; Scott, A.L.; Nurse, C.A.; Jonz, M.G. Single-cell transcriptomic analysis of neuroepithelial cells and other cell types of the gills of zebrafish (Danio rerio) exposed to hypoxia. Sci. Rep. 2022, 12, 10144. [Google Scholar] [CrossRef] [PubMed]
- Zachar, P.C.; Pan, W.; Jonz, M.G. Distribution and morphology of cholinergic cells in the branchial epithelium of zebrafish (Danio rerio). Cell Tissue Res. 2017, 367, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Basu, N.; Scheuhammer, A.M.; Rouvinen-Watt, K.; Grochowina, N.; Klenavic, K.; Evans, R.D.; Man Chan, H. Methylmercury impairs components of the cholinergic system in captive mink (Mustela vison). Toxicol. Sci. 2006, 91, 202–209. [Google Scholar] [CrossRef] [PubMed]
- De Jesus, T.B.; Colombi, J.S.; Ribeiro, C.A.O.; De Assis, H.C.S.; De Carvalho, C.E.V. Cholinesterase activity in methylmercury and mercury chloride exposure fish. Ecotoxicol. Environ. Contam. 2013, 8, 147–148. [Google Scholar] [CrossRef]
- Chen, H.; Diao, X.; Wang, H.; Zhou, H. An integrated metabolomic and proteomic study of toxic effects of benzo[a]pyrene on gills of the pearl oyster Pinctada martensii. Ecotoxicol. Environ. Saf. 2018, 156, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Sullivan, H.; Seligman, C.; Gilchrist, S.; Wang, B. An NMR-based metabolomics study on sea anemones Exaiptasia diaphana (Rapp, 1829) with atrazine exposure. Mol. Omi. 2021, 17, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; You, L.; Zhou, D.; Wang, Q.; Li, F.; Cong, M.; Li, L.; Zhao, J.; Liu, D.; et al. Benzo(a)pyrene-induced metabolic responses in Manila clam Ruditapes philippinarum by proton nuclear magnetic resonance (1H NMR) based metabolomics. Environ. Toxicol. Pharmacol. 2011, 32, 218–225. [Google Scholar] [CrossRef]
- Karakach, T.K.; Huenupi, E.C.; Soo, E.C.; Walter, J.A.; Afonso, L.O.B. 1H-NMR and mass spectrometric characterization of the metabolic response of juvenile Atlantic salmon (Salmo salar) to long-term handling stress. Metabolomics 2009, 5, 123–137. [Google Scholar] [CrossRef]
- Qin, H.; Yu, Z.; Zhu, Z.; Lin, Y.; Xia, J.; Jia, Y. The integrated analyses of metabolomics and transcriptomics in gill of GIFT tilapia in response to long term salinity challenge. Aquac. Fish. 2022, 7, 131–139. [Google Scholar] [CrossRef]
- Li, Q.-Q.; Xiang, Q.-Q.; Lian, L.-H.; Chen, Z.-Y.; Luo, X.; Ding, C.-Z.; Chen, L.-Q. Metabolic profiling of nanosilver toxicity in the gills of common carp. Ecotoxicol. Environ. Saf. 2021, 222, 112548. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Y.; Wang, F.; Luo, Z.; Guo, S.; Strähle, U. Toxicity of mercury: Molecular evidence. Chemosphere 2020, 245, 125586. [Google Scholar] [CrossRef] [PubMed]
- Kerper, L.E.; Ballatori, N.; Clarkson, T.W. Methylmercury transport across the blood-brain barrier by an amino acid carrier. Am. J. Physiol. Integr. Comp. Physiol. 1992, 262, R761–R765. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Unoki, T.; Yoshida, E.; Ding, Y.; Yamakawa, H.; Shinkai, Y.; Ishii, I.; Kumagai, Y. Repression of mercury accumulation and adverse effects of methylmercury exposure is mediated by cystathionine γ-lyase to produce reactive sulfur species in mouse brain. Toxicol. Lett. 2020, 330, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Kazak, L.; Cohen, P. Creatine metabolism: Energy homeostasis, immunity and cancer biology. Nat. Rev. Endocrinol. 2020, 16, 421–436. [Google Scholar] [CrossRef]
- Borchel, A.; Verleih, M.; Rebl, A.; Kühn, C.; Goldammer, T. Creatine metabolism differs between mammals and rainbow trout (Oncorhynchus mykiss). Springerplus 2014, 3, 510. [Google Scholar] [CrossRef]
- Gronczewska, J.; Niedźwiecka, N.; Grzyb, K.; Skorkowski, E.F. Bioenergetics of fish spermatozoa with focus on some herring (Clupea harengus) enzymes. Fish Physiol. Biochem. 2019, 45, 1615–1625. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Metabolites | Gills | ||
|---|---|---|---|
| E7 | E14 | PE28 | |
| Amino acids | |||
| Leucine | ↑ 12% | ↑ 59% * | ↓ 12% |
| Isoleucine | ↑ 10% | ↑ 37% * | ↓ 21% * |
| Valine | ↑ 20% * | ↑ 43% * | ↓ 16% |
| Alanine | ↑ 31% * | ↑ 65% * | ↓ 4% |
| Tyrosine | ↑ 9% | ↑ 27% * | ↓ 37% * |
| Phenylalanine | ↑ 34% * | ↑ 54% * | ↓ 24% * |
| Energy-related | |||
| Lactate | ↑ 102% * | ↑ 166% * | ↑ 99% * |
| Malonate | ↑ 32% * | ↑ 64% * | ↓ 16% * |
| Glycogen | ↓ 69% * | ↓ 31% * | ↑ 5% |
| Glucose | ↓ 18% * | ↑ 17% * | no change |
| Osmolytes | |||
| Taurine | ↑ 58% * | ↑ 64% * | ↑ 156% * |
| Glycerophosphocholine | ↑ 122% * | ↑ 162% * | ↑ 228% * |
| Neutransmitters | |||
| Acetylcholine | ↑ 68% * | ↑ 50% * | ↑ 149% * |
| Miscellaneous | |||
| Phosphocholine | ↑ 83% * | ↑ 126% * | ↑ 116% * |
| Inosine | ↑ 67% * | ↑ 195% * | ↑ 111% * |
| Metabolites | Liver | ||
|---|---|---|---|
| E7 | E14 | PE28 | |
| Amino acids | |||
| Alanine | ↑ 32% * | ↑ 30% * | ↑ 13% |
| Glutamate | ↓ 32% * | ↓ 73% * | ↓ 42% * |
| Glutamine | ↓ 33% * | ↓ 24% * | ↓ 7% |
| Energy-related | |||
| Glucose | ↓ 41% * | ↓ 52% * | ↓ 41% * |
| ATP/ADP | ↑ 18% | ↑ 40% * | ↑ 31% * |
| Glucose | ↓ 41% * | ↓ 52% * | ↓ 41% * |
| ATP/ADP | ↑ 18% | ↑ 40% * | ↑ 31% * |
| Miscellaneous | |||
| Creatine | ↓ 20% * | ↓ 39% * | ↓ 27% * |
| Cystathionine | ↑ 2% | ↑ 79% * | ↑ 112% * |
| Glutathione | ↑ 6% | ↓ 4% | ↓ 16% * |
| Niacinamide | ↑ 55% * | ↑ 58% * | ↑ 73% * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Marco, G.; Billè, B.; Brandão, F.; Galati, M.; Pereira, P.; Cappello, T.; Pacheco, M. Differential Cell Metabolic Pathways in Gills and Liver of Fish (White Seabream Diplodus sargus) Coping with Dietary Methylmercury Exposure. Toxics 2023, 11, 181. https://doi.org/10.3390/toxics11020181
De Marco G, Billè B, Brandão F, Galati M, Pereira P, Cappello T, Pacheco M. Differential Cell Metabolic Pathways in Gills and Liver of Fish (White Seabream Diplodus sargus) Coping with Dietary Methylmercury Exposure. Toxics. 2023; 11(2):181. https://doi.org/10.3390/toxics11020181
Chicago/Turabian StyleDe Marco, Giuseppe, Barbara Billè, Fátima Brandão, Mariachiara Galati, Patrícia Pereira, Tiziana Cappello, and Mário Pacheco. 2023. "Differential Cell Metabolic Pathways in Gills and Liver of Fish (White Seabream Diplodus sargus) Coping with Dietary Methylmercury Exposure" Toxics 11, no. 2: 181. https://doi.org/10.3390/toxics11020181
APA StyleDe Marco, G., Billè, B., Brandão, F., Galati, M., Pereira, P., Cappello, T., & Pacheco, M. (2023). Differential Cell Metabolic Pathways in Gills and Liver of Fish (White Seabream Diplodus sargus) Coping with Dietary Methylmercury Exposure. Toxics, 11(2), 181. https://doi.org/10.3390/toxics11020181