
Comparing Ocular Toxicity of Legacy and Alternative Per- and Polyfluoroalkyl Substances in Zebrafish Larvae


Abstract
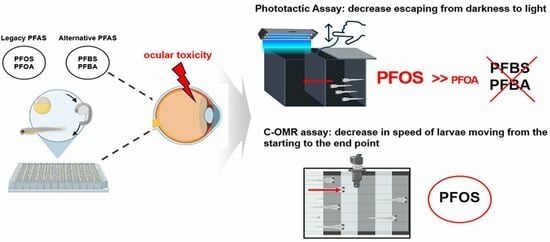
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Embryo Collection and Developmental Toxicity Testing
2.3. Phototactic Response Assay
2.4. C-OMR (Contrast-Optomotor) Assay
2.5. Gene Expression Measurement
2.6. Statistical Analysis
3. Results and Discussion
3.1. Developmental Toxicity after PFOS, PFOA, PFBS, and PFBA Exposure
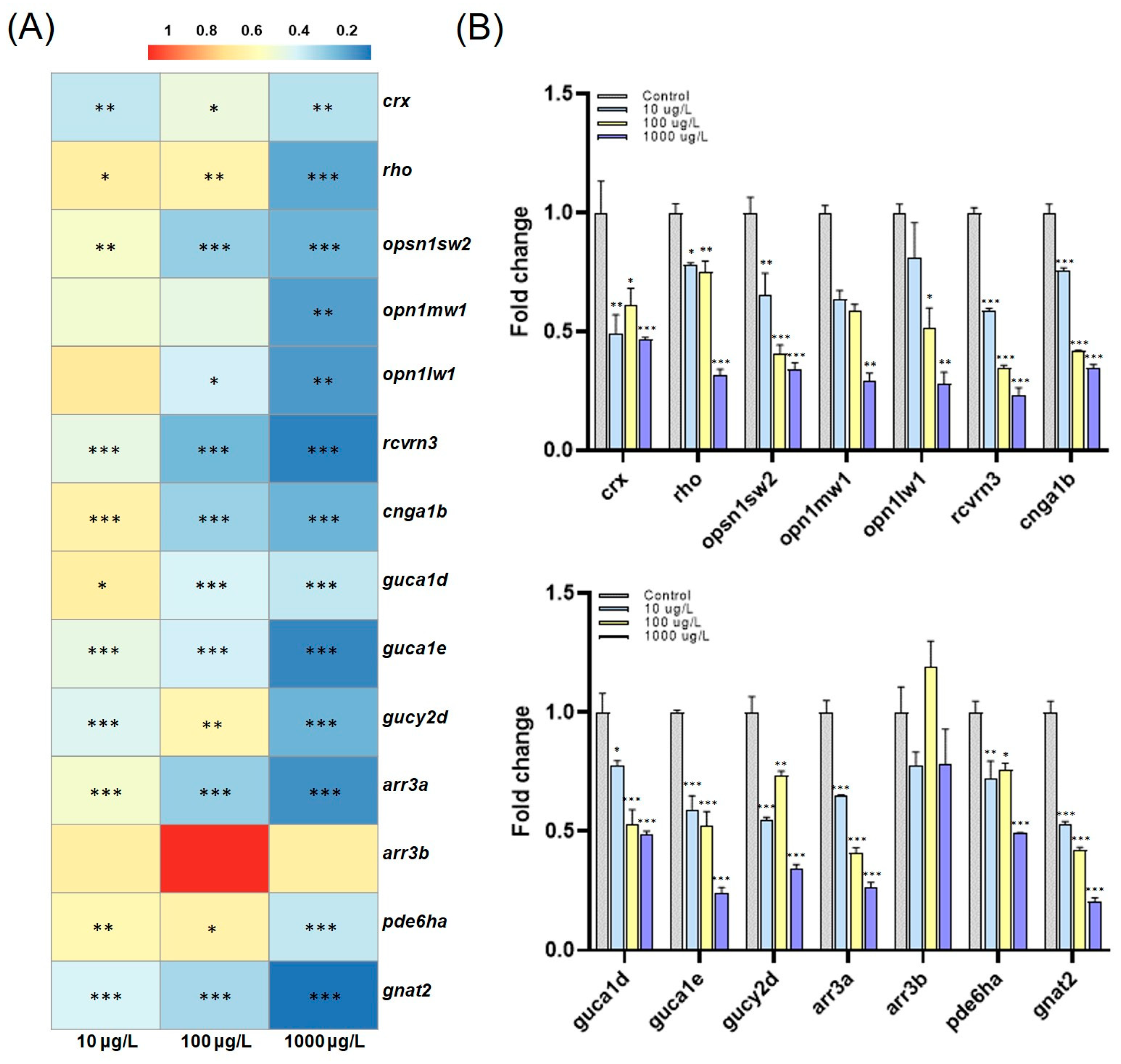
3.2. Changes in Phototactic Responses after PFOS, PFOA, PFBS, and PFBA Exposure
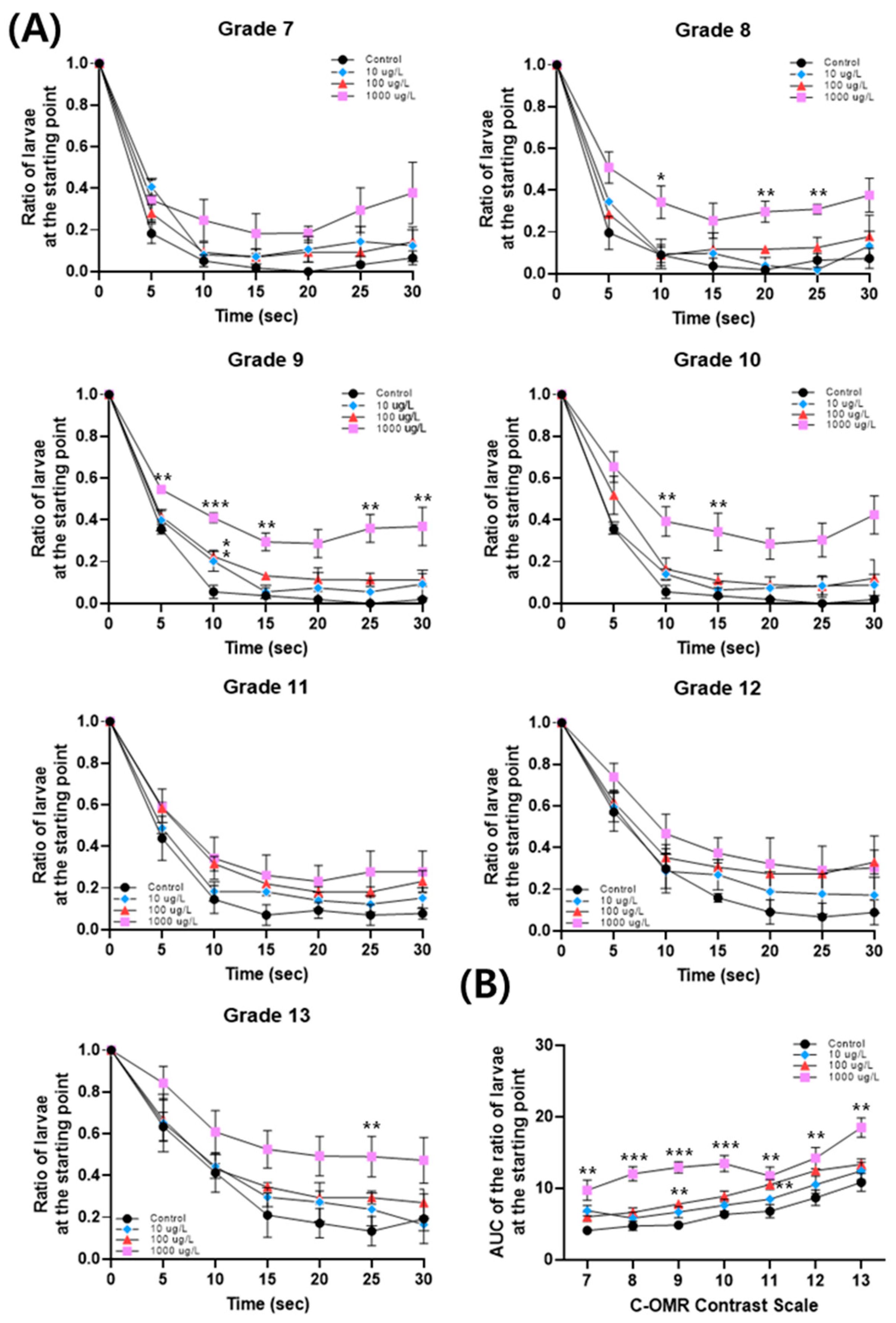
3.3. PFOS-Induced Changes in C-OMR Behavior
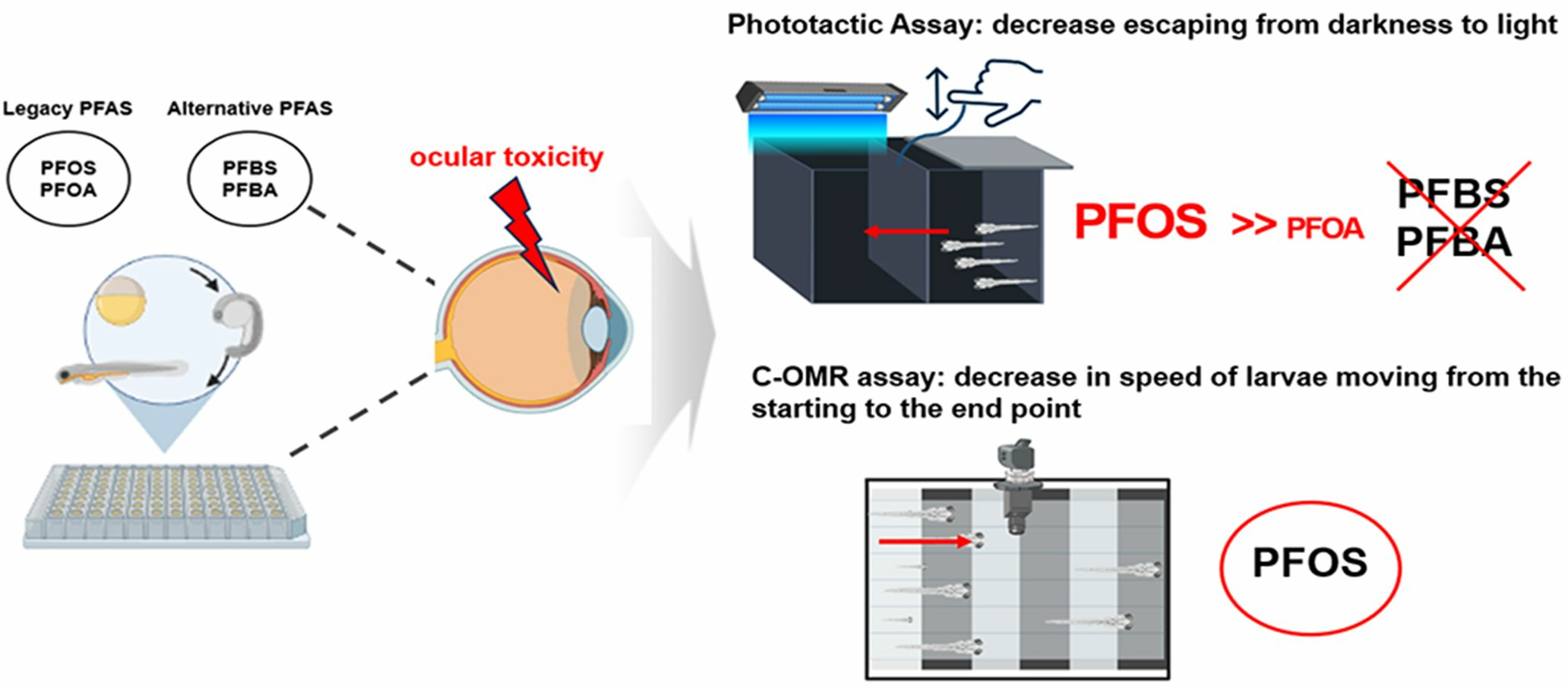
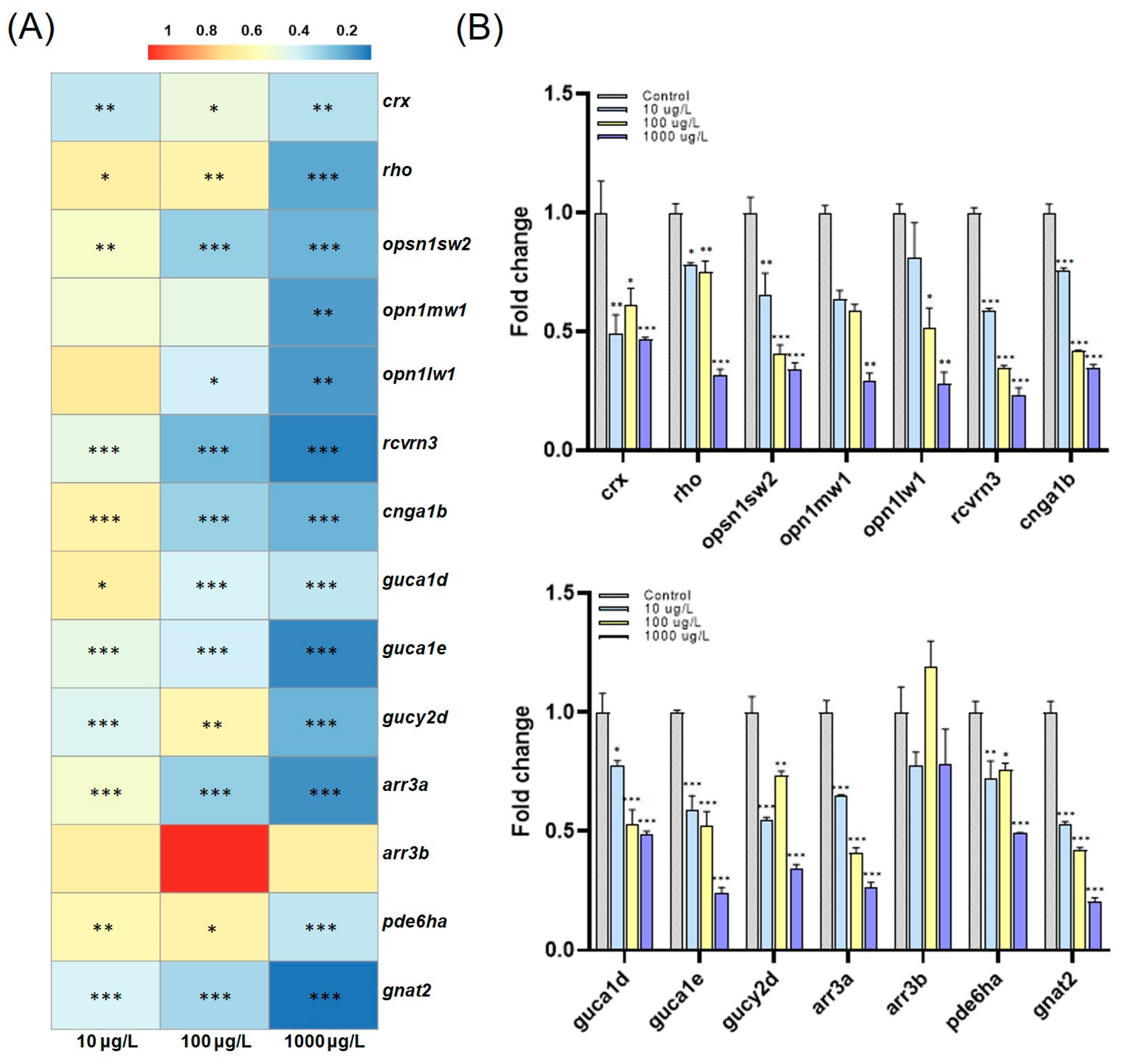
3.4. PFOS-Induced Altered Gene Expression
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuccaro, P.; Thompson, D.C.; de Boer, J.; Watterson, A.; Wang, Q.; Tang, S.; Shi, X.; Llompart, M.; Ratola, N.; Vasiliou, V. Artificial turf and crumb rubber infill: An international policy review concerning the current state of regulations. Environ. Chall. 2022, 9, 100620. [Google Scholar] [CrossRef] [PubMed]
- Land, M.; de Wit, C.A.; Bignert, A.; Cousins, I.T.; Herzke, D.; Johansson, J.H.; Martin, J.W. What is the effect of phasing out long-chain per- and polyfluoroalkyl substances on the concentrations of perfluoroalkyl acids and their precursors in the environment? A systematic review. Environ. Evid. 2018, 7, 4. [Google Scholar] [CrossRef]
- Hu, X.C.; Andrews, D.Q.; Lindstrom, A.B.; Bruton, T.A.; Schaider, L.A.; Grandjean, P.; Lohmann, L.; Carignan, C.C.; Blum, A.; Balan, S.A.; et al. Detection of Poly- and Perfluoroalkyl Substances (PFASs) in U.S. Drinking Water Linked to Industrial Sites, Military Fire Training Areas, and Wastewater Treatment Plants. Environ. Sci. Technol. Lett. 2016, 3, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Macheka, L.R.; Abafe, O.A.; Mugivhisa, L.L.; Olowoyo, J.O. Occurrence and infant exposure assessment of per and polyfluoroalkyl substances in breast milk from South Africa. Chemosphere 2022, 288, 132601. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Lu, Y.; Lu, Y.; Wang, T.; Zhu, Z.; Li, Q.; Meng, J.; Su, H.; Johnson, A.C.; Sweetman, A.J. Coupled production and emission of short chain perfluoroalkyl acids from a fast developing fluorochemical industry: Evidence from yearly and seasonal monitoring in Daling River Basin, China. Environ. Pollut. 2016, 218, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tsui, M.M.P.; Shi, Q.; Hu, C.; Wang, Q.; Zhou, B.; Lam, P.K.; Lam, J.C.W. Accumulation of perfluorobutane sulfonate (PFBS) and impairment of visual function in the eyes of marine medaka after a life-cycle exposure. Aquat. Toxicol. 2018, 201, 1–10. [Google Scholar] [CrossRef]
- Wang, Z.; Cousins, I.T.; Scheringer, M.; Hungerbuehler, K. Hazard assessment of fluorinated alternatives to long-chain perfluoroalkyl acids (PFAAs) and their precursors: Status quo, ongoing challenges and possible solutions. Environ. Int. 2015, 75, 172–179. [Google Scholar] [CrossRef]
- Gestri, G.; Link, B.A.; Neuhauss, S.C.F. The visual system of zebrafish and its use to model human ocular Diseases. Dev. Neurobiol. 2012, 72, 302–327. [Google Scholar] [CrossRef]
- Fitzgerald, J.A.; Könemann, S.; Krümpelmann, L.; Županič, A.; vom Berg, C. Approaches to Test the Neurotoxicity of Environmental Contaminants in the Zebrafish Model: From Behavior to Molecular Mechanisms. Environ. Toxicol. Chem. 2021, 40, 989–1006. [Google Scholar] [CrossRef]
- Carvalho, P.S.M.; Tillitt, D.E. 2,3,7,8-TCDD effects on visual structure and function in swim-up rainbow trout. Environ. Sci. Technol. 2004, 38, 6300–6306. [Google Scholar] [CrossRef]
- Huang, L.; Zuo, Z.; Zhang, Y.; Wu, M.; Lin, J.J.; Wang, C. Use of toxicogenomics to predict the potential toxic effect of Benzo(a)pyrene on zebrafish embryos: Ocular developmental toxicity. Chemosphere 2014, 108, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Hong, Q.; Qin, D.N.; Kou, C.Z.; Zhang, C.M.; Guo, M.; Guo, X.R.; Chi, X.; Tong, M.L. Effects of embryonic exposure to polychlorinated biphenyls on zebrafish (Danio rerio) retinal development. J. Appl. Toxicol. 2012, 32, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Hen Chow, E.S.; Yu Hui, M.N.; Cheng, C.W.; Cheng, S.H. Cadmium affects retinogenesis during zebrafish embryonic development. Toxicol. Appl. Pharmacol. 2009, 235, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zeeshan, M.; Dang, Y.; Liang, L.Y.; Gong, Y.C.; Li, Q.Q.; Tan, Y.W.; Fan, Y.Y.; Lin, L.Z.; Yang, Z.; et al. Environmentally relevant concentrations of F–53B induce eye development disorders-mediated locomotor behavior in zebrafish larvae. Chemosphere 2022, 308, 136130. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish, 3rd ed.; University of Oregon Press: Eugene, OR, USA, 2022. [Google Scholar]
- Lindstrom, A.B.; Strynar, M.J.; Delinsky, A.D.; McMilan, L.; Libelo, E.L.; Neill, M.; Thomas, L. Application of WWTP biosolids and resulting perfluorinated compound contamination of surface and well water in Decatur, Alabama, USA. Environ. Sci. Technol. 2011, 45, 8015–8021. [Google Scholar] [CrossRef]
- Yin, T.; Chen, H.; Reinhard, M.; Yi, X.; He, Y.; Gin, K.Y.H. Perfluoroalkyl and polyfluoroalkyl substances removal in a full-scale tropical constructed wetland system treating landfill leachate. Water Res. 2017, 125, 418–426. [Google Scholar] [CrossRef]
- Brockerhoff, S.E.; Hurleyt, J.B.; Janssen-Bienholdt, U.; Neuhauss, S.C.F.; Driever, W.; Dowling, J.E. A Behavioral Screen for Isolating Zebrafish Mutants with Visual System Defects. Proc. Natl. Acad. Sci. USA 1995, 92, 10545–10549. [Google Scholar] [CrossRef]
- Fromme, H.; Tittlemier, S.A.; Völkel, W.; Wilhelm, M.; Twardella, D. Perfluorinated compounds—Exposure assessment for the general population in western countries. Int. J. Hyg. Environ. Health 2009, 212, 239–270. [Google Scholar] [CrossRef]
- Liu, W.; Dong, G.; Luo, Y.; Liu, L.; Cao, Z.; Li, X.; Jin, Y. Estimation of Reference Values for PFOS and PFOA in Human Biomonitoring and Relevance of Exposure among Family Members in China. J. Environ. Prot. 2012, 3, 353–361. [Google Scholar] [CrossRef]
- Jin, Y.H.; Saito, N.; Harada, K.H.; Inoue, K.; Koizumi, A. Historical Trends in Human Serum Levels of Perfluorooctanoate and Perfluorooctane Sulfonate in Shenyang, China. Tohoku J. Exp. Med. 2007, 212, 63–70. [Google Scholar] [CrossRef]
- Jin, Y.H.; Dong, G.H.; Shu, W.Q.; Ding, M.; Zhai, C.; Wang, L.; Liu, X.; Saito, N.; Sasaki, K. Comparison of Perfluorooctane Sulfonate and Perfluorooctane Acid in Serum of Non-Occupational Human from Shenyang and Chongqing Areas. J. Hyg. Res. 2006, 35, 560–563. [Google Scholar]
- Koun, S.; Eom, Y.; Kim, M.J.; Kim, S.; Lee, H.I.; Park, H.C.; Song, J.S.; Kim, H.M. Development of an experimental model for ocular toxicity screening in Zebrafish. Biochem. Biophys. Res. Commun. 2021, 559, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Corbo, C.C. Partitioning of gene expression among zebrafish photoreceptor subtypes. Sci. Rep. 2021, 11, 17340. [Google Scholar] [CrossRef] [PubMed]
- Vasamsetti, B.M.K.; Chon, K.; Yoon, C.Y.; Kim, J.; Choi, J.Y.; Hwang, S.; Park, K.H. Transcriptome Profiling of Etridiazole-Exposed Zebrafish (Danio rerio) Embryos Reveals Pathways Associated with Cardiac and Ocular Toxicities. Int. J. Mol. Sci. 2023, 24, 155067. [Google Scholar] [CrossRef]
- Stiebel-Kalish, H.; Reich, E.; Rainy, N.; Vatine, G.; Nisgav, Y.; Tovar, A.; Gothilf, Y.; Bach, M. Gucy2f zebrafish knockdown—A model for Gucy2d-related leber congenital amaurosis. Eur. J. Hum. Genet. 2012, 20, 884–889. [Google Scholar] [CrossRef] [PubMed]
- McCurley, A.T.; Callard, G.V. Characterization of housekeeping genes in zebrafish: Male-female differences and effects of tissue type, developmental stage and chemical treatment. BMC Mol. Biol. 2008, 9, 102. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Organization for Economic Co-Operation and Development [OECD]. Guidelines for the Testing of Chemicals, Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD Publishing: Paris, France, 2013. [Google Scholar]
- Deeti, S.; O’Farrell, S.; Kennedy, B.N. Early safety assessment of human oculotoxic drugs using the zebrafish visualmotor response. J. Pharmacol. Toxicol. Methods 2014, 69, 1–8. [Google Scholar] [CrossRef]
- Wolf, S.; Dubreuil, A.M.; Bertoni, T.; Böhm, U.L.; Bormuth, V.; Candelier, R.; Karpenko, S.; Hildebrand, D.G.C.; Bianco, I.H.; Monasson, R.; et al. Sensorimotor computation underlying phototaxis in zebrafish. Nat Commun. 2017, 8, 651. [Google Scholar] [CrossRef]
- Maurer, C.M.; Schönthaler, H.B.; Mueller, K.P.; Neuhauss, S.C.F. Distinct retinal deficits in a zebrafish pyruvate dehydrogenase-deficient mutant. J. Neurosci. 2010, 30, 11962–11972. [Google Scholar] [CrossRef]
- Fernandes, A.M.; Fero, K.; Arrenberg, A.B.; Bergeron, S.A.; Driever, W.; Burgess, H.A. Deep brain photoreceptors control light-seeking behavior in zebrafish larvae. Curr. Biol. 2012, 22, 2042–2047. [Google Scholar] [CrossRef] [PubMed]
- Ulhaq, M.; Örn, S.; Carlsson, G.; Morrison, D.A.; Norrgren, L. Locomotor behavior in zebrafish (Danio rerio) larvae exposed to perfluoroalkyl acids. Aquat. Toxicol. 2013, 144–145, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Zhang, J.; Chen, Y.; Wang, L.; Wang, M.; Xiong, D.; Sun, Y. Combined Effects of PFOS and PFOA on Zebrafish (Danio rerio) Embryos. Arch. Environ. Contam. Toxicol. 2013, 64, 668–675. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, X.; Wei, P.; Tian, H.; Wang, W.; Ru, S. Long-term exposure to bisphenol S damages the visual system and reduces the tracking capability of male zebrafish (Danio rerio). J. Appl. Toxicol. 2018, 3, 248–258. [Google Scholar] [CrossRef]
- Faria, B.M.; Duman, F.; Zheng, C.X.; Waisbourd, M.; Gupta, L.; Ali, M.; Zangalli, C.; Lu, L.; Wizov, S.S.; Spaeth, E.; et al. Evaluating contrast sensitivity in age-related macular degeneration using a novel computer-based test, the spaeth/richman contrast sensitivity test. Retina 2015, 35, 1465–1473. [Google Scholar] [CrossRef]
- Wilensky, J.T.; Hawkins, A. Comparison of contrast sensitivity, visual acuity, and Humphrey visual field testing in patients with glaucoma. Trans. Am. Ophthalmol. Soc. 2001, 99, 213–217. [Google Scholar] [PubMed]
- Shi, Q.; Wang, Z.; Chen, L. Optical toxicity of triphenyl phosphate in zebrafish larvae. Aquat. Toxicol. 2019, 210, 139–147. [Google Scholar] [CrossRef]
- Wang, F.; Shih, K.M.; Li, X.Y. The partition behavior of perfluorooctanesulfonate (PFOS) and perfluorooctanesulfonamide (FOSA) on microplastics. Chemosphere 2015, 119, 841–847. [Google Scholar] [CrossRef]
- Fuentes, S.; Vicens, P.; Colomina, M.T.; Domingo, J.L. Behavioral effects in adult mice exposed to perfluorooctane sulfonate (PFOS). Toxicology 2007, 242, 123–129. [Google Scholar] [CrossRef]
- Wang, F.; Liu, C.; Shih, K. Adsorption behavior of perfluorooctanesulfonate (PFOS) and perfluorooctanoate (PFOA) on boehmite. Chemosphere 2012, 89, 1009–1014. [Google Scholar] [CrossRef]
- Christou, M.; Ropstad, E.; Brown, S.; Kamstra, J.H.; Fraser, T.W.K. Developmental exposure to a POPs mixture or PFOS increased body weight and reduced swimming ability but had no effect on reproduction or behavior in zebrafish adults. Aquat. Toxicol. 2021, 237, 105882. [Google Scholar] [CrossRef]
- Lee, J.E.; Choi, K. Perfluoroalkyl substances exposure and thyroid hormones in humans: Epidemiological observations and implications. Ann. Pediatr. Endocrinol. Metab. 2017, 22, 6–14. [Google Scholar] [CrossRef]
- Zeeshan, M.; Yang, Y.; Zhou, Y.; Huang, W.; Wang, Z.; Zeng, X.Y.; Liu, R.Q.; Yang, B.Y.; Hu, L.W.; Zeng, S.W. Incidence of ocular conditions associated with perfluoroalkyl substances exposure: Isomers of C8 Health Project in China. Environ. Int. 2020, 137, 105555. [Google Scholar] [CrossRef]
- Pacione, L.R.; Szeg, M.J.; Ikeda, S.; Nishina, P.M.; McInnes, R.R. Progress toward understanding the genetic and biochemical mechanisms of inherited photoreceptor degenerations. Annu. Rev. Neurosci. 2003, 26, 657–700. [Google Scholar] [CrossRef]
- Morris, A.C.; Fadool, J.M. Studying rod photoreceptor development in zebrafish. Physiol. Behav. 2005, 86, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Bridges, K.N.; Magnuson, J.T.; Curran, T.E.; Barker, A.; Roberts, A.P.; Venables, B.J. Alterations to the vision-associated transcriptome of zebrafish (Danio rerio) following developmental norethindrone exposure. Environ. Toxicol. Pharmacol. 2019, 69, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Doerre, G.; Malicki, J. Genetic analysis of photoreceptor cell development in the zebrafish retina. Mech. Dev. 2002, 110, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Hertz, H.; Blancas-Velazquez, A.S.; Rath, M.F. The role of homeobox gene-encoded transcription factors in regulation of phototransduction: Implementing the primary pinealocyte culture as a photoreceptor model. J. Pineal Res. 2021, 71, e12753. [Google Scholar] [CrossRef] [PubMed]
- Krupnick, J.G.; Gurevich, V.V.; Benovic, J.L. Mechanism of quenching of phototransduction: Binding competition between arrestin and transducin for phosphorhodopsin. J. Biol. Chem. 1997, 272, 18125–18131. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.P.; Brown, B.M.; Craft, C.M. Visual arrestin 1 acts as a modulator for N-ethylmaleimide-sensitive factor in the photoreceptor synapse. J. Neurosci. 2010, 30, 9381–9391. [Google Scholar] [CrossRef]
- Iqubal, A.; Ahmed, M.; Ahmad, S.; Sahoo, R.C.; Iqubal, K.M.; Haque, S.E. Environmental neurotoxic pollutants: Review. Environ. Sci. Pollut. Res. Int. 2020, 27, 41175–41198. [Google Scholar] [CrossRef] [PubMed]
- Lamb, T.D.; Pugh, E.N. Phototransduction, dark adaptation, and rhodopsin regeneration: The proctor lecture. Investig. Ophthalmol. Vis. Sci. 2006, 47, 5138–5152. [Google Scholar] [CrossRef] [PubMed]
- Muto, A.; Orger, M.B.; Wehman, A.M.; Smear, M.C.; Kay, J.N.; Page-McCaw, P.S.; Gahtan, E.; Xiao, T.; Nevin, L.M.; Gosse, L.J.; et al. Forward Genetic Analysis of Visual Behavior in Zebrafish. PLoS Genet. 2005, 1, e66. [Google Scholar] [CrossRef] [PubMed]
- Chhetri, J.; Jacobson, G.; Gueven, N. Zebrafish—On the move towards ophthalmological research. Eye 2014, 28, 367–380. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-s.; Jang, S.; Eom, Y.; Kim, K.-T. Comparing Ocular Toxicity of Legacy and Alternative Per- and Polyfluoroalkyl Substances in Zebrafish Larvae. Toxics 2023, 11, 1021. https://doi.org/10.3390/toxics11121021
Lee H-s, Jang S, Eom Y, Kim K-T. Comparing Ocular Toxicity of Legacy and Alternative Per- and Polyfluoroalkyl Substances in Zebrafish Larvae. Toxics. 2023; 11(12):1021. https://doi.org/10.3390/toxics11121021
Chicago/Turabian StyleLee, Han-seul, Soogyeong Jang, Youngsub Eom, and Ki-Tae Kim. 2023. "Comparing Ocular Toxicity of Legacy and Alternative Per- and Polyfluoroalkyl Substances in Zebrafish Larvae" Toxics 11, no. 12: 1021. https://doi.org/10.3390/toxics11121021
APA StyleLee, H.-s., Jang, S., Eom, Y., & Kim, K.-T. (2023). Comparing Ocular Toxicity of Legacy and Alternative Per- and Polyfluoroalkyl Substances in Zebrafish Larvae. Toxics, 11(12), 1021. https://doi.org/10.3390/toxics11121021